 ,1, 姬改革1, 单艳菊1, 章明1, 巨晓军1, 刘一帆1, 屠云洁1, 盛中伟1, 唐燕飞2, 蒋华莲2, 邹剑敏,1
,1, 姬改革1, 单艳菊1, 章明1, 巨晓军1, 刘一帆1, 屠云洁1, 盛中伟1, 唐燕飞2, 蒋华莲2, 邹剑敏,1Expression Analysis of IGF1-PI3K-Akt-Dependent Pathway Genes in Skeletal Muscle and Liver Tissue of Yellow Feather Broilers
SHU JingTing,1, JI GaiGe1, SHAN YanJu1, ZHANG Ming1, JU XiaoJun1, LIU YiFan1, TU YunJie1, SHENG ZhongWei1, TANG YanFei2, JIANG HuaLian2, ZOU JianMin,1通讯作者:
责任编辑: 林鉴非
收稿日期:2020-07-28接受日期:2020-10-16网络出版日期:2021-05-01
| 基金资助: |
Received:2020-07-28Accepted:2020-10-16Online:2021-05-01
作者简介 About authors
束婧婷,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (477KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
束婧婷, 姬改革, 单艳菊, 章明, 巨晓军, 刘一帆, 屠云洁, 盛中伟, 唐燕飞, 蒋华莲, 邹剑敏. IGF1-PI3K-Akt信号通路相关基因在黄羽肉鸡肌肉和肝脏中的表达[J]. 中国农业科学, 2021, 54(9): 2027-2038 doi:10.3864/j.issn.0578-1752.2021.09.018
SHU JingTing, JI GaiGe, SHAN YanJu, ZHANG Ming, JU XiaoJun, LIU YiFan, TU YunJie, SHENG ZhongWei, TANG YanFei, JIANG HuaLian, ZOU JianMin.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】禽类的生长发育是一个复杂的生理过程,不同生长阶段体重以及肌肉的生长速度与强度在不同品种以及不同生长环境之间有着较大差异,不同遗传背景对禽类的生长以及肌肉的产量有着重要影响[1,2]。研究表明,骨骼肌的“肥大”/“萎缩”,是由蛋白质合成/降解之间的平衡来控制的。IGF1-PI3K- Akt信号通路既参与蛋白质的合成又参与蛋白质的降解,对骨骼肌的发育、细胞增殖和转化等具有极其重要的作用[3,4],是调控生长发育的关键通路,肌肉的生长发育与 IGF1-PI3K-Akt通路相关基因的时空特异性表达密切相关[5,6,7]。探究IGF1-PI3K-Akt信号通路相关基因在不同黄羽肉鸡品种(系)肝脏和骨骼肌中的表达差异对于阐明黄羽肉鸡肌肉生长发育分子机理具有重要意义,将有助于了解鸡生长过程中激素调控的本质。【前人研究进展】IGFs系统由3个配体(IGF1,IGF2和胰岛素),3个受体(IGF1R,IGF2R和胰岛素受体IR)和6种结合蛋白(IGFBPs 1-6)组成[8]。IGF1是动物体内最重要的生长因子之一,IGF1可以通过IGF1-PI3K-Akt信号通路促进骨骼肌干细胞的增殖、肌细胞的分化以及成肌细胞融合为肌管[3,9],对动物胚胎以及出生后的生长发育有重要作用。肝脏是产生IGF1的主要场所,但许多组织细胞均可产生IGF1和IGF2,从而发挥自分泌和旁分泌作用。IGF1促进细胞生长的长期效应主要是通过IGF1R介导的[10,11]。IGF1R是一个2型络氨酸激酶跨膜糖蛋白受体,通常以包含2个细胞外α亚基和2个跨膜β亚基的异源二聚体的形式存在[12]。肌肉中IGF1R活性的丧失将导致肌纤维数目的减少和直径变小,从而影响骨骼肌的生长发育[13]。胰岛素受体底物(insulin receptor substrate,IRS)是一种信号传导蛋白,广泛分布于胰岛素敏感组织中。IRS1是IRS家族最早发现的成员,是胰岛素信号通路的重要中间体,调节肌细胞的葡萄糖转运,促进肌细胞的生长、增殖和分化。IRS1是IGF1的主要底物,处于IGF1R的下游,IGF1配体通过结合α亚基从而激活β亚基的内源酪氨酸激酶活性,进而结合并激活IRS和Shc,磷酸化的IRS1通过结合PI3K的p85调节亚基进一步激活PI3K-AKT-mTOR信号通路。在小鼠中,敲除 IRS1下调 IGF1的表达[14]和导致体重减轻 40%—70%[15]。在鸡上,在成肌细胞中过表达和敲低 IRS1,发现IRS1可以促进细胞增殖[16]。由此可见,IRS1是IGF1-PI3K-AKT通路的关键节点,在 IGF1 信号通路的作用十分重要,可直接影响 IGF1的表达进而调控肌肉的生长。【本研究切入点】IGF1-PI3K-AKT信号通路相关基因IGF1、IGF1R和IRS1对肌肉的生长发育具有重要的调控作用,目前关于这些基因在不同生长速度的黄羽肉鸡(广西麻鸡和花山麻鸡)不同部位肌肉和肝脏中表达情况还没有文献详细报道。【拟解决的关键问题】黄羽肉鸡按生长速度分为慢速型、中速型和快速型。广西麻鸡是我国重要的慢速型地方品种,其肉质鲜美,深受广大消费者欢迎,但其生长速度较慢(120日龄上市,平均上市体重1.5 kg)。花山麻鸡是利用快大型肉鸡和黄羽肉鸡杂交培育而成的适合屠宰后冷鲜上市的中速型黄羽肉鸡配套系(63—65日龄上市,平均上市体重1.9 kg),具有屠体美观、生长速度快、肉质较优等优点。本研究即以慢速型的广西麻鸡和中速型的花山麻鸡作为研究对象,研究其从胚胎期至出生后早期的生长发育特点,以及IGF1-PI3K-AKT信号通路相关基因IGF1、IGF1R和IRS1在胸腿肌和肝脏中的发育性表达规律,为进一步阐明IGF1-PI3K-AKT信号通路影响黄羽肉鸡肌肉发育的分子机理提供一定的参考,为黄羽肉鸡“分子编写育种”提供一定的遗传学依据。1 材料与方法
1.1 试验时间、地点
本试验于2019年4—8月在扬州江苏省家禽科学研究所养殖基地和家禽遗传育种重点实验室进行。1.2 试验动物
广西麻鸡种蛋来源于广西富凤农牧有限公司广西麻鸡保种群,花山麻鸡种蛋来源于江苏兴牧农业科技有限公司育种群。种蛋孵化前,进行称重、消毒和编号,品种内蛋重变异系数在5%以内。2个品种种蛋随机置入同一孵化箱,在相同条件下(温度为37.5—37.8℃,相对湿度为65%—75%)同时孵化。出雏后,在统一的条件下饲养至63日龄。种蛋入孵后的24 h设定为1胚龄,分别于9个时间点(9胚龄、12胚龄、16胚龄和出雏后0日龄、7日龄、21日龄、35日龄、49日龄和63日龄)采样,胚胎期种蛋用解剖刀刀柄敲碎钝端蛋壳,以无菌镊子移去蛋壳,换一无菌镊子轻夹胚胎颈部取出鸡胚放置准备好的吸水纸上,然后移至称量纸记录胚重(或体重),将一侧腿去爪后带骨称重,一侧胸带骨分离并称重。采集另一侧胸大肌和腿肌同一部位组织样品,采集肝脏称重后,置于液氮速冻,然后转入-80℃冰箱保存。每个时间点每个品种采样20只,胚胎期利用CHD1基因多态性鉴别公母,出雏后根据性腺组织判别公母。性别鉴定后,每个时间点每个品种选取母鸡6只用于测定。1.3 总RNA提取和cDNA合成
肌肉总RNA提取按TRNzol Universal 总RNA提取试剂(DP424)(天根,北京)的说明书进行。RNA样品经琼脂糖凝胶电泳鉴定总RNA完整性后,采用核酸定量仪NanoDrop2000(Thermo scientific,美国)测定浓度及根据A260nm/A280nm值确定RNA纯度。取2 μg总RNA,按照cDNA 第一链合成试剂盒HiScript Ⅲ RT SuperMix for qPCR(+gDNA wiper)(R323-01)(诺唯赞,南京)的使用说明书进行cDNA第一链的合成,RT产物保存在-20℃备用。1.4 引物设计
选择鸡肌肉特异性内参基因HSP70作为本研究的内参基因[17]。根据GenBank中鸡的IGF1 (GenBank登录号:NM_001004384.2)和 IGF1R (GenBank登录号:NM_205032.1 )相关cDNA序列设计引物,IRS1引物参考LI等[16]报道的序列进行设计,由上海英俊生物工程有限公司合成,引物信息见表1。Table 1
表1
表1基因引物序列
Table 1
| 基因名称 Gene name | 引物序列 Primer sequence | PCR条件 PCR conditions (℃) |
|---|---|---|
| IGF1 | 5′ primer: AGCTGGTTGATGCTCTTCAGTTCGT | 60 |
| 3′ primer: TCCTCAGGTCACAACTCTGGAAGC | ||
| IGF1R | 5′ primer: TTCAGGAACCAAAGGGCGA | 55 |
| 3′ primer: TGTAATCTGGAGGGCGATACC | ||
| IRS1 | 5′ primer: CTACCGCCTGTGCCTGACTAAC | 60 |
| 3′ primer: CGTCCCACCTCGATGAAGAAG | ||
| HSP70 | 5′ primer: TCTGCTCCTGTTGGATGTC | 60 |
| 3′ primer: TGGGAATGGTGGTGTTACG |
新窗口打开|下载CSV
1.5 实时荧光定量PCR
荧光定量PCR采用SYBR Green Ι法,参照ChamQ SYBR Color qPCR Master Mix(Q411-02)(诺唯赞,南京)试剂盒说明书进行。将每个待测样品cDNA产物取等体积混合后,用混合样做模板,对实时荧光定量PCR的退火温度、引物浓度、模板浓度等条件进行优化。20 μL的反应体系如下:2×ChamQ SYBR Color qPCR Master Mix 10 μL,上下游引物(10 μmol·L-1)各0.4 μL,50×ROX Reference Dye2 0.4 μL,模板2 μL,加ddH2O至20 μL。反应条件为:95℃ 30 s;95℃ 10 s,55℃或60℃ 30 s,95℃ 15 s,40个循环;95℃ 15s,60℃ 60 s,95℃ 15 s。 每次反应均设空白样品为阴性对照,每个样品设置3个平行。1.6 统计分析
定量的结果采用2-ΔΔCt法进行处理,分析基因的相对表达量。将广西麻鸡0日龄设为对照组,ΔΔCt=ΔCt(其他日(胚)龄)-ΔCt(0日龄)。采用 SPSS 20.0统计分析软件进行分析,组间差异显著性用独立样本T检验,One-way ANOVA进行统计分析。Bivariate Correlation用来分析基因表达之间以及基因表达与生长发育相关性。所有数据以Mean ± SE表示,P<.05,表示差异显著;P<.01,表示差异极显著。2 结果
2.1 生长速度不同的两个品种鸡生长发育过程中体重、肌肉和肝脏的发育性变化
两个品种鸡生长发育过程中体重的变化如图1-A所示,两个品种体重均呈持续上升态势,存在显著的胚龄(日龄)和品种效应。在所研究的9个发育阶段,花山麻鸡胚重(体重)均高于广西麻鸡,除在9和12胚龄差异不显著外(P>0.05),其他胚龄(日龄)均达到显著水平(P<0.05或P<0.01),尤其表现在出雏后,花山麻鸡体重的增长速度远快于广西麻鸡。两个品种胸肌重的变化趋势基本一致(图1-B),都是随着胚龄(日龄)的增长,胸肌重量逐渐增加,值得指出的是,在0日龄(出雏)时,两个品种的胸肌重均显著下降,显著低于16胚龄时的胸肌重(P<0.05),与12胚龄时的胸肌重量无显著差异(P>0.05),从7日龄开始,花山麻鸡的胸肌重均显著高于广西麻鸡(P<0.05或P<0.01)。两个品种腿肌重随日龄的增长持续增加(图1-C),品种间比较发现,总体上花山麻鸡的腿肌重要高于广西麻鸡,在胚胎期差异不显著,从出雏开始即表现出显著的差异(P<0.05或P<0.01)。两个品种肝脏重的变化趋势基本一致(图1-D),从16胚龄到63日龄,两个品种鸡肝脏重显著(P<0.05)递增;品种间比较发现,在胚胎期以及出雏时,两个品种鸡肝脏重相差不显著(P>0.05),从7日龄开始,花山麻鸡肝脏重显著(P<0.05)或极显著(P<0.01)高于广西麻鸡。图1
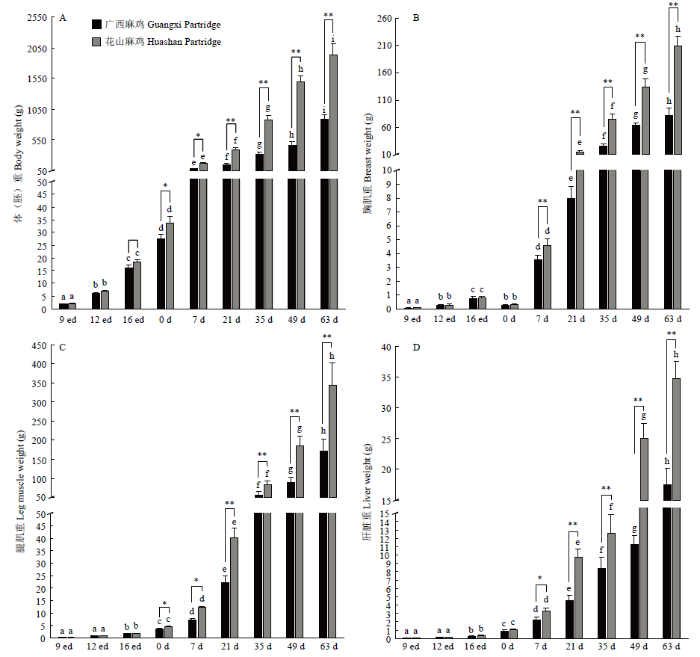 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同品种鸡生长发育过程中体重、胸肌重、腿肌重和肝脏重的变化
A:体重;B:胸肌重;C:腿肌重;D:肝脏重。ed表示胚龄,d表示日龄;品种内不同胚龄或日龄比较,不同小写字母表示差异显著(P<0.05),相同小写字母表示差异不显著(P>0.05)*表示同一胚龄的不同品种间差异显著(P<0.05),**表示同一胚龄的不同品种间差异极显著(P<0.01)
Fig. 1The profiles of body weight, breast muscle weight, thigh muscle weight and liver weight during growth stages in different chicken breeds
A: Body weight; B: Breast muscle weight; C: Leg muscle weight; D: Liver weight. ed indicates embryonic days, and d indicates age after hatching. Bar diagram values with the same letter are not significantly different between different age (P>0.05), and values with the different letters are significantly different between different ages (P<0.05). Superscript * indicates significantly different between two breeds in the same age at the level of 0.05, and superscript ** indicates significantly different between two breeds in the same age at the level of 0.01
2.2 IGF1基因在不同鸡种胚胎期和出生后早期胸肌、腿肌和肝脏中的表达规律
IGF1基因在广西麻鸡和花山麻鸡不同发育阶段胸肌、腿肌和肝脏中的表达存在显著的胚(日)龄、组织和品种效应,结果见表2。胸肌中,两个品种鸡IGF1 mRNA的表达均呈现先升高后降低的表达模式,在9胚龄时表达量较高,到12胚龄达到最高,显著高于后续的胚(日)龄(P<0.05),随后显著下降(P<0.05),并在后续的发育阶段均维持在较低的水平,63日龄表达量最低。在腿肌中,两个品种IGF1 mRNA的表达变化趋势较为一致,在9胚龄时表达量最高,9胚龄至12胚龄基本保持稳定,随后显著下降(P<0.05),至出雏时降至最低,7日龄开始回升,随后21日龄至63日龄基本保持稳定。在肝脏中,广西麻鸡IGF1 mRNA的表达呈现先升高后降低的表达模式,胚胎期未检测到表达,出雏即有表达,随后升高,至21日龄达到表达高峰,相对表达量为7日龄时的14倍(P<0.05),随后维持在较高的表达水平,但63日龄又有显著下降(P<0.05)。花山麻鸡肝脏中IGF1 mRNA的表达同样呈现先升高后降低的表达模式,在胚胎期表达量极低,9—16胚龄仅微量表达,出雏时显著升高(P<0.05),随后持续升高,至21日龄表达量显著上调(P<0.05),相对表达量达到7日龄时的13倍,接着继续上升,至49日龄时达到表达高峰后基本保持稳定。Table 2
表2
表2广西麻鸡和花山麻鸡不同发育时期骨骼肌和肝脏中IGF1、IGF1R和IRS1基因的相对表达量
Table 2
| 基因Gene | 组织Tissue | 品种 Breed | 9 ed | 12 ed | 16 ed | 0 d | 7 d | 21 d | 35 d | 49 d | 63 d |
|---|---|---|---|---|---|---|---|---|---|---|---|
| IGF1 | 胸肌 Breast | GXMJ | 9.65±1.06 a | 9.78±0.37a | 5.18±1.07b | 1.51±0.33 c | 0.27±0.08d | 0.19±0.05d | 0.14±0.03d | 0.06±0.005e | 0.05±0.005e |
| HSMJ | 1.12±0.18b** | 1.92±0.26a** | 0.98±0.26b** | 0.42±0.07c* | 0.24±0.08c | 0.18±0.05c | 0.17±0.03c | 0.13±0.04c | 0.05±0.004d | ||
| 腿肌 Leg | GXMJ | 54.92±2.51a | 54.77±5.06a | 33.20±3.39 b | 1.32±0.39c | 2.08±0.37cd | 2.86±0.44d | 2.51±0.48cd | 3.42±0.85d | 1.58±0.16c | |
| HSMJ | 10.44±1.5a** | 10.31±0.2a** | 1.77±0.20b** | 0.26±0.65c* | 2.74±0.57d | 1.53±0.26b | 3.79±0.86d | 2.25±0.42b | 1.06±0.18b | ||
| 肝脏 Liver | GXMJ | 0 | 0 | 0 | 1.23±0.23c | 4.02±1.07c | 57.62±6.46a | 47.84±5.54a | 52.47±7.05a | 29.81±3.59b | |
| HSMJ | 0.04±0.01e* | 0.22±0.02d* | 0.31±0.04d | 3.02±0.52c* | 5.11±1.47c | 66.06±6.99b* | 74.44±9.4ab* | 80.87±12.0a* | 79.71±9.1a** | ||
| IGF 1R | 胸肌 Breast | GXMJ | 3.50±0.23 a | 1.64±0.15b | 2.63±0.33b | 1.32±0.33 c | 0.06±0.01d | 0.08±0.01d | 0.10±0.02d | 0.03±0.00d | 0.04±0.00d |
| HSMJ | 0.45±0.07a** | 0.24±0.04b** | 0.25±0.06b** | 0.17±0.07bc* | 0.02±0.00c | 0.02±0.00c | 0.05±0.00c | 0.03±0.00c | 0.01±0.00c | ||
| 腿肌 Leg | GXMJ | 6.14±0.86a | 4.21±0.56b | 2.31±0.37c | 1.18±0.28d | 0.46±0.08d | 0.37±0.05d | 0.49±0.05d | 0.30±0.04d | 0.21±0.06d | |
| HSMJ | 1.24±0.23a** | 1.21±0.27a* | 0.18±0.05b** | 0.10±0.01b** | 0.17±0.02b | 0.12±0.02b | 0.29±0.06b | 0.17±0.02b | 0.17±0.01b | ||
| 肝脏 Liver | GXMJ | 0.15±0.02c | 0.16±0.02 c | 0.37±0.03c | 0.96±0.06b | 1.62±0.19ab | 1.96±0.27a | 1.48±0.16ab | 1.54±0.19ab | 1.35±0.28ab | |
| HSMJ | 0.21±0.05d | 0.36±0.04d | 0.50±0.03d | 1.21±0.18c | 1.77±0.23bc | 1.78±0.30bc | 1.88±0.24bc | 2.60±0.35b* | 5.74±0.91a* | ||
| IRS1 | 胸肌 Breast | GXMJ | 2.07±0.09a | 1.69±0.12a | 1.51±0.16a | 1.50±0.13a | 0.17±0.02b | 0.30±0.06b | 0.37±0.03b | 0.22±0.03b | 0.21±0.02b |
| HSMJ | 0.56±0.05a** | 0.39±0.03a* | 0.31±0.02a* | 0.55±0.06a* | 0.09±0.01c | 0.16±0.01b | 0.21±0.03b | 0.13±0.04b | 0.02±0.00c* | ||
| 腿肌 Leg | GXMJ | 5.84±0.59a | 4.32±0.49b | 1.93±0.11c | 1.51±0.04cd | 0.79±0.06cd | 1.07±0.13cd | 0.90±0.10cd | 0.80±0.06cd | 0.61±0.05d | |
| HSMJ | 2.08±0.26a** | 2.28±0.54a* | 0.63±0.10b* | 0.73±0.04b* | 0.26±0.04b* | 0.40±0.06b* | 0.60±0.09b | 0.48±0.07b | 0.38±0.04b | ||
| 肝脏 Liver | GXMJ | 0.09±0.00c | 0.22±0.04c | 0.10±0.01c | 0.92±0.20b | 1.80±0.27ab | 3.08±0.46a | 2.84±0.55a | 3.02±0.74a | 2.63±0.22a | |
| HSMJ | 0.08±0.00f | 0.11±0.01f | 0.08±0.01f | 0.76±0.13e | 1.52±0.11e | 2.58±0.15d | 3.53±0.53c | 6.29±0.73b* | 16.05±2.01a** |
新窗口打开|下载CSV
品种间比较发现,在胸腿肌中,IGF1 mRNA在广西麻鸡胚胎期至出雏的表达量都显著(P<0.05)或极显著(P<0.01)高于花山麻鸡,而出雏后至63日龄,两者差异不显著(P>0.05);而在肝脏中IGF1基因则是在花山麻鸡中的表达要高于广西麻鸡,且除7日龄差异不显著(P>0.05)外,其他日(胚)龄均达到了显著水平(P<0.05)。组织间比较发现,两品种腿肌中IGF1基因各阶段的表达量均高于胸肌组织,在胚胎期,肌肉中IGF1基因的表达量均高于肝脏,而在出雏后的各发育阶段,肝脏中IGF1基因的表达量要高于肌肉组织,且在肝脏和肌肉中的表达量变化规律截然相反。
2.3 IGF1R基因在不同鸡种胚胎期和出生后早期胸肌、腿肌和肝脏中的表达规律
IGF1R基因在广西麻鸡和花山麻鸡不同发育阶段胸肌、腿肌和肝脏中的表达存在显著的胚(日)龄、组织和品种效应,结果见表2。IGF1R基因在两个品种胸肌中的表达变化趋势较为一致,在9胚龄时表达量最高,到12胚龄显著下调(P<0.05),16胚龄略有升高后持续下调,7日龄下降幅度最大,达到显著水平(P<0.05),然后在整个出生后早期表达量均维持在极低水平,值得指出的是,IGF1R基因在花山麻鸡所研究的发育阶段胸肌中的表达量均极低。IGF1R基因在两个品种腿肌中的表达模式较为一致,在9胚龄时表达量最高,在广西麻鸡中要显著高于其他胚(日)龄(P<0.05),而花山麻鸡中除了与12胚龄差异不显著外(P>0.05),与其他胚(日)龄的表达量均达到显著水平(P<0.05),随后持续下降,在出雏后的各个发育阶段表达量均极低。在肝脏中,广西麻鸡IGF1R 基因的表达呈现先升高后降低的表达模式,在9胚龄时表达量最低,然后持续升高至21日龄达到表达高峰后略有下降,但基本保持稳定,总体上出雏(0日龄)及出雏后的表达量显著高于胚胎期(P<0.05),21日龄表达量显著高于0日龄(P<0.05),而与其他日龄差异不显著(P>0.05);花山麻鸡IGF1R 基因的表达呈现持续升高的表达趋势,9胚龄表达量最低,随后升高,但整个胚胎期表达量差异不显著(P>0.05),出雏时显著升高(P<0.05),随后持续升高,至63日龄时达到表达高峰,表达量显著高于其他日(胚)龄(P<0.05),总体上出雏(0日龄)及出雏后的表达量显著高于胚胎期(P<0.05)。品种间比较来看,总体上,IGF1R基因在广西麻鸡胸腿肌中的表达量要高于花山麻鸡,在胚胎期至出雏达到显著(P<0.05)或极显著(P<0.01)水平,而出雏后品种间差异不显著(P>0.05);而在肝脏中的表达量则是花山麻鸡高于广西麻鸡,在49和63日龄达到显著水平(P<0.05)。组织间比较发现,总体上两品种腿肌中IGF1R基因的表达量高于胸肌组织,胚胎期至出雏(0日龄),肌肉中IGF1R基因的表达量要高于肝脏,而在出雏后的各发育阶段,肝脏中IGF1R基因的表达量要高于肌肉组织,且肝脏和肌肉中的表达量变化规律在胚胎期和出生后早期截然相反。
2.4 IRS1基因在不同鸡种胚胎期和出生后早期胸肌、腿肌和肝脏中的表达规律
IRS1基因在广西麻鸡和花山麻鸡不同发育阶段胸肌、腿肌和肝脏中的表达存在显著的胚(日)龄、组织和品种效应,结果见表2。在胸肌中,IRS1基因在两品种鸡中的表达趋势较为一致,在9胚龄时表达量最高,在胚胎期至出雏基本维持稳定,到7日龄显著下降(P<0.05),随后均维持在较低水平。在腿肌中,IRS1基因在广西麻鸡中呈现先下调后上调再下调的表达趋势,9胚龄时表达量最高,并显著高于其他胚(日)龄(P<0.05),随后逐渐降低并保持在相对平稳表达状态,63日龄表达量最低;IRS1基因在花山麻鸡中呈现先升高后降低的趋势,在12胚龄时达到峰值,之后显著降低(P<0.05)并保持平稳表达状态,9胚龄,12胚龄表达量显著高于其他胚(日)龄(P<0.05),其他胚(日)龄间表达量差异不显著(P>0.05)。在肝脏中,广西麻鸡IRS1 基因的表达呈现先升高后降低的表达模式,在9胚龄时表达量最低,然后持续升高至21日龄达到表达高峰后略有下降,但基本保持稳定,总体上出雏(0日龄)及出雏后的表达量显著高于胚胎期(P<0.05),21日龄表达量显著高于0日龄(P<0.05),而与其他日龄差异不显著(P>0.05);花山麻鸡IRS1基因的表达呈现持续升高的表达趋势,至63日龄时达到表达高峰,表达量显著高于其他日(胚)龄(P<0.05),总体上出雏(0日龄)及出雏后的表达量显著高于胚胎期(P<0.05),胚胎期各胚龄间表达量差异不显著(P>0.05),而出雏后各日龄间差异均达到显著水平(P<0.05)。品种间比较来看,总体上,IGF1R基因在广西麻鸡胸腿肌中的表达量要高于花山麻鸡,在胚胎期至出雏达到显著(P<0.05)水平,而出雏后胸肌中除63日龄差异显著(P<0.05)外,其他日龄点差异均不显著(P>0.05),腿肌中除7日龄和21日龄差异显著(P<0.05)外,其他日龄点差异均不显著(P>0.05)。组织间比较发现,总体上两品种腿肌中IRS1基因的表达量高于胸肌组织,胚胎期至出雏(0日龄),肌肉中IRS1基因的表达量要高于肝脏,而在出雏后的各发育阶段,肝脏中IRS1基因的表达量要高于肌肉组织,且肝脏和肌肉中的表达量变化规律在胚胎期和出生后早期截然相反。
2.5 IGF1-PI3K-AKT信号通路相关基因表达的相关性
为深入了解IGF1-PI3K-AKT信号通路相关基因在鸡骨骼肌和肝脏早期发育过程中的相互作用机制,对IGF1,IGF1R以及IRS1基因在广西麻鸡和花山麻鸡胸腿肌和肝脏中的表达相关性进行分析见表3。由表可见,3个基因在两品种的胸腿肌和肝脏中均表现出极显著的正相关关系(P<0.01),即IGF1基因与IGF1R 基因表达水平之间存在极显著的正相关(P<0.01),IGF1和IRS1 基因表达水平之间存在极显著的正相关(P<0.01),IGF1R 和 IRS1基因表达水平之间存在极显著的正相关(P<0.01),这些结果表明所研究的IGF1-PI3K-AKT信号通路相关基因之间存在显著的正调控关系。Table 3
表3
表3广西麻鸡和花山麻鸡骨骼肌和肝脏中IGF1-PI3K-AKT信号通路相关基因表达的相关性
Table 3
| 参数 Parameters | 广西麻鸡Guangxi Partridge | 花山麻鸡Huashan Partridge | |||
|---|---|---|---|---|---|
| IGF1R | IRS1 | IGF1R | IRS1 | ||
| 胸肌 Breast muscle | IGF1 | 0.851** | 0.641** | 0.418** | 0.457** |
| IRS1 | 0.806** | 0.648** | |||
| 腿肌 Leg muscle | IGF1 | 0.884** | 0.873** | 0.719** | 0.670** |
| IRS1 | 0.854** | 0.884** | |||
| 肝脏 Liver | IGF1 | 0.494** | 0.682** | 0.489** | 0.389** |
| IRS1 | 0.759** | 0.911** | |||
新窗口打开|下载CSV
2.6 IGF1-PI3K-AKT信号通路相关基因表达量与生长性状的相关性
IGF1-PI3K-AKT信号通路对于动物的生长发育起到重要作用,但在不同物种中其作用方式也不同,目前在黄羽肉鸡中尚未见该信号通路对生长发育作用的系统报道,故本研究进一步对IGF1-PI3K-AKT信号通路相关基因表达量与体重等生长性状的相关性进行了分析,详见表4。由表可见,IGF1,IGF1R以及IRS1基因在两个品种鸡胸肌和腿肌中的表达量均与体重、胸(腿)肌重呈显著或极显著的负相关(P<0.05 or P<0.01),而在肝脏中的表达量与体重、胸肌重、腿肌重、肝脏重呈显著或极显著的正相关(P<0.05 or P<0.01)。Table 4
表4
表4不同鸡种IGF1-PI3K-AKT信号通路相关基因表达量与生长性状的相关性
Table 4
| 组织 Tissue | 参数 Parameters | 广西麻鸡Guangxi Partridge | 花山麻鸡Huashan Partridge | ||||
|---|---|---|---|---|---|---|---|
| IGF1 | IGF1R | IRS1 | IGF1 | IGF1R | IRS1 | ||
| 胸肌 Breast muscle | 体重Body weight | -0.453** | -0.537** | -0.467** | -0.519* | -0.430** | -0.594** |
| 胸肌重Breast weight | -0.405** | -0.483** | -0.424** | -0.441** | -0.361** | -0.542** | |
| 腿肌 Leg muscle | 体重Body weight | -0.413** | -0.471** | -0.451** | -0.389** | -0.332** | -0.370** |
| 腿肌重Leg weight | -0.377** | -0.435** | -0.417** | -0.362** | -0.284* | -0.321** | |
| 肝脏 Liver | 体重Body weight | 0.369* | 0.965** | 0.817** | 0.693** | 0.781** | 0.817** |
| 肝脏重Liver weight | 0.458** | 0.409** | 0.851** | 0.601** | 0.794** | 0.851** | |
| 胸肌重Breast weight | 0.352** | 0.453** | 0.783** | 0.563** | 0.696** | 0.752** | |
| 腿肌重Leg weight | 0.365** | 0.483** | 0.812** | 0.598** | 0.723** | 0.795** | |
新窗口打开|下载CSV
3 讨论
黄羽肉鸡是我国传统消费习惯形成的特殊消费食品,现已成为优质肉鸡的代名词,按生长速度可分为快速型(50—60 d出栏)、中速型(60—90 d出栏)和慢速型(90 d以上出栏)。黄羽肉鸡肉质好,但生长速度慢,产肉量少,因此,如何提高黄羽肉鸡产肉量和生产效率,是国内肉鸡育种工作者急需解决的问题。本研究选用慢速型的广西麻鸡和中速型的花山麻鸡作为研究黄羽肉鸡肌肉生长发育的试验模型,发现在所研究的发育阶段,两个品种的体重、腿肌重和肝脏重均呈持续增长趋势,而胸肌重则在胚胎发育晚期出现负增长现象,与课题组在清远麻鸡上的研究结果一致[17],也与禽类孵化后期组织和器官发育特点相符,即家禽孵化后期,由于尿囊的氧供应能力及营养供应达到极限,机体需要进行糖酵解、糖异生等糖代谢提供能量,因此,此阶段肝脏快速发育以满足代谢需要[18];胸肌是肝脏外糖原贮存的另一重要组织,在孵化后期当能量发生危机时,胸肌中蛋白质发生动员为糖的异生提供氨基酸底物,因此胚胎晚期鸭[18,19,20]、火鸡[21]等家禽胸肌会出现质量减轻。本试验发现鸡生长发育早期体重、骨骼肌重和肝脏重变化呈现显著的品种和时间特异性,经过选育的中速型花山麻鸡体重和组织重的增长在胚胎发育后期已经远远超过慢速型的广西麻鸡,说明遗传背景的差异是影响黄羽肉鸡生长发育的重要因素。本研究发现生长速度不同的黄羽肉鸡胚胎期和生后期肌肉和肝脏中IGF1-PI3K-Akt通路关键基因IGF1、IGF1R和IRS1的表达差异。总体上,在同一鸡种的同一个部位,3个基因的表达规律是一致的,在肌肉中表现出胚胎期表达量较高,出雏后下降的趋势,而在肝脏中则表现出胚胎期表达量较低,出雏后快速上调的趋势;相关性分析证实所研究的基因两两之间均存在极显著的正相关关系(P<0.01),提示所研究的3个基因在鸡胚胎期及出生后早期表达存在一致性,充分说明3个基因在功能上有着密切的关系。该结果进一步验证了前人的研究结果,IGF1 结合并激活 IGF1R,IGF1R 活化后,促进 IRS1 磷酸化,磷酸化的 IRS1 进一步激活PI3K和Akt,进而使整个 IGF1 信号通路活化[3,9]。
胰岛素样生长因子1(IGF1)是动物体内最重要的生长因子之一,动物体内近乎所有器官的生长和功能都与其有关[22]。IGF1 可以通过IGF1/IGF1R/PI3K/ Akt 信号通路促进骨骼肌干细胞的增殖、肌细胞的分化以及成肌细胞融合为肌管[3,23]。本研究发现,在所研究的发育阶段,骨骼肌中均能检测到IGF1基因的表达,总体上胚胎期的表达水平高于出雏后,然而在两个品种鸡胸肌和腿肌中的表达趋势并不一致,胸肌中IGF1表达高峰出现在12胚龄,腿肌中则在9胚龄即出现表达高峰,说明9胚龄至12胚龄很可能是鸡胸腿肌早期发育中的一个快速生长点,这一阶段是初级纤维形成的时期,IGF1基因在这一阶段的高表达也表明该基因可能对成肌细胞增殖的作用更大。另外由于胸肌和腿肌的发育依赖于不同的生肌程序和发育通路,相对于腿肌而言,胸肌属于晚熟部位,且胸腿肌在肌纤维类型组成上存在差异[24,25],这可能是IGF1基因在胸腿肌中的差异表达原因所在。肝脏是禽类出生后分泌IGF1的主要场所,肝脏中分泌IGF1不足将直接导致机体循环中IGF1浓度降低,从而影响动物的生长。本研究中,IGF1在两品种鸡胚胎期肝脏中的表达量均极低或者无表达,出雏开始表达量迅速升高,与KIKUCHI等[26]在鸡上的研究结果一致。IGF1基因的表达存在品种和组织差异,在胚胎期至出雏,广西麻鸡胸肌和腿肌中的表达量要显著高于花山麻鸡,出雏后品种间差异不显著;而在肝脏中IGF1基因在花山麻鸡中的表达量除7日龄外均显著高于广西麻鸡。组织间比较发现,胚胎期两品种鸡IGF1表达量均为腿肌>胸肌>肝脏,而出雏后则为肝脏>腿肌>胸肌,该结果与DUPONT [27]的结论一致,即禽类胚胎期血液中IGF1的来源与出雏后的来源不同,出生后IGF1的主要来源是肝组织,而在胚胎发育期间IGF1主要来源于肝外组织。
IGF1R基因在两品种鸡胸肌和腿肌中的表达趋势较为一致,均在9胚龄时表达量最高,然后持续下降并维持在较低的水平,与本课题组在清远麻鸡上的研究结果[17]以及在猪背最长肌上发现IGF1R mRNA 的表达随年(胎)龄增加而减少的结果[28]一致。在胚胎期至出雏,广西麻鸡胸腿肌中的表达量均显著高于花山麻鸡,我们推测IGF1R基因跟IGF1基因功能一致,在成肌细胞的增殖中发挥重要作用。在所研究的发育阶段肝脏中均能检测到IGF1R mRNA的表达,同样表现为出生后的表达量显著高于胚胎期,两个品种均是9胚龄时表达量最低,但高峰期存在品种差异,花山麻鸡63日龄最高,与其他各阶段差异显著,广西麻鸡则21日龄时表达量最高,且总体上花山麻鸡中的表达量要高于广西麻鸡,这也可能与不同生长速度黄羽肉鸡的生长发育规律有关,说明肝脏中IGF1R基因及其表达产物可能对鸡的生长发育有着重要的调节作用。
IRS1 在介导 IGF1 信号转导中发挥重要作用,是 IGF1对生物机体肌肉生长、分化调节功能的必需蛋白分子[29]。目前对鸡IRS1基因及其在不同组织中的表达规律等报道较少。本研究发现,在两品种鸡的各个发育阶段均能检测到IRS1 mRNA的表达,且表达模式存在品种、组织和胚(日)龄差异,在胚胎期胸腿肌中的表达量要高于出生后,表达高峰同样出现在9胚龄或12胚龄,说明IRS1可能对促进成肌细胞增殖具有重要作用,这也与LI等[16]研究结论一致。IGF1、IGF1R和IRS1基因在胸腿肌中均表现在9胚龄或12胚龄高表达,说明这3个基因在骨骼肌纤维的形成过程中发挥了重要作用。IRS1基因在两品种鸡胚胎期肝脏中的表达量均维持在较低的水平,9胚龄时表达量最低,从出雏开始即显著上调,广西麻鸡21日龄时表达量达到最高,而花山麻鸡则到63日龄达到最高,可能与两个品种鸡的生长发育规律相关。
IGF1、IGF1R和IRS1等3个基因在骨骼肌中的表达量与两品种鸡的体重和肌肉生长呈显著或极显著的负相关关系,再一次说明鸡骨骼肌中IGF1-PI3K-Akt通路相关基因是在促进胚胎期肌纤维形成而非肌纤维肥大中发挥作用。肝脏中3个基因的表达量与广西麻鸡和花山麻鸡的体重、胸肌重、腿肌重以及肝脏重则呈显著或极显著的正相关关系,与荣华等[30]在大围山微型鸡、本课题组在鸭[20]上的研究结果一致,说明在禽类生长发育过程中,IGF1信号通路发挥着重要作用,而肝脏则是重要的分泌器官。为了更深入了解IGF1-PI3K-Akt信号通路基因对不同类型黄羽肉鸡生长发育的调控规律,今后还应从蛋白水平上分析基因表达与生长性状的关系。
4 结论
本试验利用实时荧光定量PCR探究了IGF1-PI3K- Akt信号通路相关基因IGF1、IGF1R和IRS1在慢速型的广西麻鸡和中速型的花山麻鸡不同发育时段肌肉和肝脏中的表达量差异,研究结果表明上述3个基因表达存在品种、胚(日)龄和组织差异,在相同品种同一组织中表达模式趋于一致,在肌肉中表现出胚胎期表达量较高,出雏后下降的趋势,而在肝脏中则表现出胚胎期表达量较低,出雏后快速上调的趋势,3个基因表达之间的正相关关系也充分说明3个基因在功能上有着密切的关系,存在正向调控机制,而品种间表达量的差异可能与不同类型黄羽肉鸡的生长发育有关,提示这3个基因可能对鸡生长发育具有一定的调控作用,为进一步研究IGF1-PI3K-Akt信号通路在黄羽肉鸡生长发育过程中发挥的功能与表达调控机理提供了新的思路。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 4]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 3]
[D].
[本文引用: 2]
[D].
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}