Construction and Analysis of Broad-Spectrum Resistance Gene Combination Pattern for Japonica Rice in Lower Region of the Yangtze River, China
WU YunYu,1,2, XIAO Ning1,2,4, YU Ling1,2, CAI Yue1,2, PAN CunHong1,3, LI YuHong1,2, ZHANG XiaoXiang1,2, HUANG NianSheng1,2, JI HongJuan1,2, DAI ZhengYuan1,3, LI AiHong,1,2,31Lixiahe Institute of Agricultural Sciences of Jiangsu, Yangzhou 225007, Jiangsu 2Jiangsu Key Laboratory of Crop Genomics and Molecular Breeding, Yangzhou University, Yangzhou 225009, Jiangsu 3Jiangsu Co-Innovation Center for Modern Production Technology of Grain Crops, Yangzhou University, Yangzhou 225009, Jiangsu 4State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193
Abstract 【Objective】Gene pyramiding is one of the most effective ways to achieve broad-spectrum resistance against Magnaporthe oryzae. The objective of this study is to construct a set of polygene pyramiding lines (PPLs) under the background of japonica rice, to evaluate their resistance performances and analysis the components of their resistance effects using M. oryzae strains collected from lower region of the Yangtze River, China, thus providing broad-spectrum resistance gene combination pattern and germplasm resources for japonica rice resistance breeding in lower region of the Yangtze River, China. 【Method】Monogenic lines with multiple alleles of the Piz locus (Pigm, Pi40, Pi9, Pi2, Pizt and Piz) with the background of japonica rice 07GY31 as the backbone, crossed with other broad-spectrum resistance gene Pi1, Pi54 and Pi33, respectively using the incomplete NCII mating design. A total of 18 different PPLs were constructed using marker-assisted selection (MAS) and agronomic traits screening. Artificial inoculation assays at seedling and heading stage with 109 representative M. oryzae strains collected from lower region of the Yangtze River, combined with natural induction identification under multiple field environments were conducted to evaluate the resistance performances of different PPLs, and to analyze the component factors of the resistance effects of the PPLs. 【Result】Genotyping by sequencing (GBS) analysis shows that the constructed PPLs all have a high background recovery rate, which was ranging from 97.08% (PPLPiz/Pi33) to 99.08% (PPLPigm/Pi1), indicated that the genetic background of all PPLs was almost fully identical to that of the recurrent parent 07GY31. The seedling blast and panicle blast resistance levels of most PPLs were significantly higher than those of monogenic lines under artificial inoculation conditions, the PPLs with better resistance to seedling blast are PPLPigm/Pi1, PPLPigm/Pi54, PPLPigm/Pi33, PPLPi9/Pi33, PPLPi9/Pi54, PPLPi40/Pi54, PPLPi40/Pi33, PPLPi40/Pi1 and PPLPi9/Pi1, respectively, and the PPLs with outstanding performance in panicle blast are PPLPigm/Pi1, PPLPigm/Pi54, PPLPigm/Pi33, PPLPi40/Pi33, PLPi40/Pi54, PPLPi40/Pi1 and PPLPizt/Pi33, respectively. Different resistance gene combinations produced different effects after pyramiding. High complementary effect and which could be fully expressed is the key factor for the improvement of the resistance level of seedling blast and panicle blast of the PPLs. In addition, PPLPigm/Pi1, PPLPigm/Pi54 and PPLPigm/Pi33 displayed broad-spectrum resistance in artificial inoculation at seedling and heading stage, and showed stable broad-spectrum resistance under different disease nurseries. Besides, agronomic traits evaluation also showed PPLs with these three gene combinations were at par to the recurrent parent. Therefore, Pigm/Pi1, Pigm/Pi54 and Pigm/Pi33 are broad-spectrum resistance gene combination patterns suitable for japonica rice resistance breeding in lower region of the Yangtze River, China. 【Conclusion】The combination pattern of resistance genes affects the resistance level of the PPLs, and high complementary effect and which could be fully expressed is the key factor for the improvement of the resistance level of the PPLs in japonica background. In addition, the development of PPLs and component factors analysis in this study provides valuable theoretical support and innovative germplasm resources for the precise breeding broad-spectrum japonica varieties in lower region of the Yangtze River, China. Keywords:lower region of the Yangtze River; japonica rice;Magnaporthe oryzae;rice blast;gene pyramiding;effect analysis
PDF (3784KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 吴云雨, 肖宁, 余玲, 蔡跃, 潘存红, 李育红, 张小祥, 黄年生, 季红娟, 戴正元, 李爱宏. 长江下游粳稻稻瘟病广谱抗性基因组合模式分析[J]. 中国农业科学, 2021, 54(9): 1881-1893 doi:10.3864/j.issn.0578-1752.2021.09.006 WU YunYu, XIAO Ning, YU Ling, CAI Yue, PAN CunHong, LI YuHong, ZHANG XiaoXiang, HUANG NianSheng, JI HongJuan, DAI ZhengYuan, LI AiHong. Construction and Analysis of Broad-Spectrum Resistance Gene Combination Pattern for Japonica Rice in Lower Region of the Yangtze River, China[J]. Scientia Acricultura Sinica, 2021, 54(9): 1881-1893 doi:10.3864/j.issn.0578-1752.2021.09.006
开放科学(资源服务)标识码(OSID):
0 引言
【研究意义】由稻瘟病菌(Magnaporthe oryzea)引起的稻瘟病是威胁世界水稻安全生产的主要真菌类病害之一[1],控制该病害最为经济有效、绿色安全的策略是利用抗病基因培育抗病品种[2],但目前已鉴定的绝大多数稻瘟病抗性基因为垂直抗性基因,其中部分抗性基因存在不能兼顾苗瘟和穗瘟抗性、抗谱窄等缺点,同时由于稻瘟病菌的致病性分化快、生理小种多、变异频繁,导致抗病品种应用3—5年后便“丧失”抗性变为感病品种。抗性基因聚合被证明是培育广谱抗病品种的主要途径之一[3,4],然而,不同的抗性基因聚合后会产生不同的抗性效应。因此,合理利用不同抗性基因进行聚合并评价其抗性效应,进而发掘有效的广谱抗性基因组合模式对培育广谱抗病品种、保障水稻安全生产具有重要现实意义和实践应用价值。【前人研究进展】宿主对病原菌的抗性反应可分为完全抗性和部分抗性两种类型,完全抗性通常由单个主效抗病基因控制,而部分抗性通常由多个微效抗性基因控制[5,6]。水稻稻瘟病主效抗病基因大多为来自STAND(signal transduction ATPases with numerous domains)亚家族中的NBS-LRR蛋白家族成员[7],NBS-LRR蛋白以直接或间接方式识别病原菌无毒蛋白后激活下游基因表达从而激发水稻抗病免疫反应[8,9,10,11]。目前,已鉴定的稻瘟病主效抗性基因超过100个,部分抗性基因约347个[12,13]。其中有28个主效抗性基因(Pit、Pish、Pi37、Pi64、Pib、Pid2、Pid3、Pi25、Pi2、Pi9、Pizt、Pi50、Pigm、Pizh、Pi36、Pi5/Pii、Pia/Pi-CO39、Pi1、Pi54/Pi54rh、Pikm、Pikp、Pik、Pike、Pita和Ptr)[14,15,16]及5个部分抗性基因(pi21、Pi35、Pi63、Pb1和Pid3-I1)[17,18,19,20,21]被成功克隆,为稻瘟病抗性育种提供了有效基因资源,但由于稻瘟病菌群体的高度变异及新的毒性种群的出现,许多携带单一抗病基因(Pi9、Pita、Pi54和Pib)的抗性品种在大规模种植一段时间后失去了抗性效应变为感病品种[22,23,24]。为降低抗性丧失风险,利用不同抗病基因进行基因聚合是培育广谱、持久抗病品种的一个有效途径。JIANG等[25]和XIAO等[26]分别报道聚合Pi1/Pi2/D12和Pi46/Pi2的改良系对中国南方稻区稻瘟病菌菌株具有广谱抗性,而聚合Pi2/Pi54则对印度稻瘟病菌群体表现出广谱抗性[4]。但抗病基因聚合后并不简单表现出抗性效应累加,有的还存在正向甚至负向互作。国际水稻研究所以Co39为背景构建携带不同抗性基因的聚合系,鉴定结果显示Piz5/Pita的基因聚合系抗性不如单基因抗性品系[27]。同样,XIAO等[28]研究发现在粳稻07GY31背景下Pi9/Pi54双基因聚合系穗瘟抗性显著低于Pi9单基因系;WU等[29]以籼稻扬稻6号为背景,构建了15种不同的抗性基因聚合系,发现Pigm/Pi1、Pigm/Pi54 和Pigm/Pi33为籼稻背景下的广谱抗性基因组合模式,同时也发现部分基因组合产生了负向互作效应。此外,由于稻瘟病菌生理小种依赖于水稻宿主生存,生理小种基因型分布与水稻宿主种植区域有关,来自籼稻、粳稻种植区域的生理小种被明显分为“籼稻群”和“粳稻群”[30,31]。因此,在不同种植区需选择与当地稻瘟病菌菌群非亲和性的抗性基因进行基因聚合,确保基因聚合后可有效提高其抗性水平。【本研究切入点】长江下游地区为我国主要粳稻种植区之一,约占全国粳稻总种植面积的40%[32,33],该地区粳稻主要含有Pish、Pita、Pib、Pikm和Pi54等抗性基因[34,35]。近年来,随着耕作制度和气候条件的变化,稻瘟病在长江下游粳稻种植区常态化发生。如2014年江苏水稻稻瘟病发生面积98万公顷,2016年上升至128.7万公顷,且发病面积呈逐年增加趋势,水稻生产存在严重安全隐患。表明长江下游应用品种现有携带的抗性基因已远不能控制稻瘟病的发生,亟需导入新的抗性基因/基因组合。【拟解决的关键问题】以粳稻07GY31为背景的Piz基因座不同复等位基因(Pigm、Pi40、Pi9、Pi2、Pizt和Piz)单基因系为核心,采用不完全NCII交配设计,分别与其他广谱抗性基因(Pi1、Pi54和Pi33)单基因系杂交,经过分子标记辅助选择和农艺性状筛选,构建18种不同基因组合的双基因聚合系。从长江下游不同粳稻种植区采集稻瘟病菌菌株进行苗瘟和穗瘟的人工接种鉴定,以及开展病圃的自然诱发鉴定,评价不同双基因聚合系的抗性效应,并分析双基因聚合系抗性效应的构成因子,从而明确适宜长江下游粳稻的稻瘟病广谱抗性基因组合模式,以期为该地区粳稻抗稻瘟病育种提供种质资源。
A、B:双基因聚合系、单基因系和轮回亲本间苗瘟和穗瘟的综合比较分析;C、D:双基因聚合系、单基因系和轮回亲本的苗瘟、穗瘟抗性表现 Fig. 2Resistance performances of PPLs, NILs and the recurrent parent for seedling and panicle blast resistance under artificial inoculation
A、B: Comprehensive comparative analysis on seedling blast and panicle blast RF of PPLs, NILs and the recurrent parent; C、D: Resistance performances of PPLs, NILs and the recurrent parent for seedling blast and panicle blast resistance
A:双基因聚合系苗瘟抗性效应分析桑基图Analysis of resistance effect of PPLs against seedling blast with sankey diagram;B:双基因聚合系穗瘟抗性效应分析桑基图Analysis of resistance effect of PPLs against panicle blast with sankey diagram。OE:重叠效应Overlapping effect;CE:互补效应Complementary effect;IE:互作效应Interaction effect Fig. 3Analysis of resistance effect of PPLs with sankey diagram
A:3个病圃的不同双基因聚合系的小区健康穗率与人工接种下穗瘟抗性频率之间的关系Relationship between HPP of PPLs at three disease nurseries and the RF of PPLs in artificial inoculation evaluation;B:双基因聚合系在不同病圃的穗瘟抗性表现Resistance performance of PPLs against panicle blast among different disease nurseries。JT:金坛Jintan;LJ:庐江Lujiang;CX:长兴Changxing Fig. 4Resistance performances of PPLs in the three blast nurseries
DEANR, VANKAN J A, PRETORIUSZ A, HAMMOND-KOSACKK E, DIPIETRO A, SPANUP D, RUDDJ J, DICKMANM, KAHMANNR, ELLISJ, FOSTERG D. The top 10 fungal pathogens in molecular plant pathology Molecular Plant Pathology, 2012,13(4):414-430. [本文引用: 1]
KHUSHG S, JENEK. Current status and future prospects for research on blast resistance in rice (Oryza sativa L.)//Advances in Genetics, Genomics and Control of Rice Blast Disease Springer, 2009: 1-10. [本文引用: 1]
GOUDAP K, SAIKUMARS, VARMAC M, NAGESHK, THIPPESWARMYS, SHENOYV, RAMESHAM S, SHASHIDHARH E. Marker-assisted breeding of Pi-1 and Piz-5 genes imparting resistance to rice blast in PRR78, restorer line of Pusa RH-10 Basmati rice hybrid Plant Breeding, 2013,132(1):61-69. [本文引用: 1]
ELLURR K, KHANNAA, YADAVA, PATHANIAS, RAJASHEKARAH, SINGHV K, GOPALAKRISHNAN S, BHOWMICKP K, NAGARAJANM, VINODK K, PRAKASHG, MONDALK K, SINGHN K, VINODPRABHU K, SINGHA K. Improvement of Basmati rice varieties for resistance to blast and bacterial blight diseases using marker assisted backcross breeding Plant Science, 2016,242:330-341. [本文引用: 2]
NIKSR E, QIX, MARCELT C. Quantitative resistance to biotrophic filamentous plant pathogens: Concepts, misconceptions, and mechanisms Annual Review of Phytopathology, 2015,53:445-470. [本文引用: 1]
WANGG L, MACKILLD J, BONMANJ M, MCCOUCHS R, CHAMPOUXM C, NELSONR J. RFLP mapping of genes conferring complete and partial resistance to blast in a durably resistant rice cultivar Genetics, 1994,136(4):1421-1434. [本文引用: 1]
LUKASIKE, TAKKENF L. STANDing strong, resistance proteins instigators of plant defence Current Opinion in Plant Biology, 2009,12(4):427-436. [本文引用: 1]
RAYS, SINGHP K, GUPTAD K, MAHATOA K, SARKARC, RATHOURR, SINGHN K, SHARMAT R. Analysis of Magnaporthe oryzae genome reveals a fungal effector, which is able to induce resistance response in transgenic rice line containing resistance gene, Pi54 Frontiers in Plant Science, 2016,7:1140. [本文引用: 1]
JIAY, MCADAMSS A, BRYANG T, HERSHEYH P, VALENTB. Direct interaction of resistance gene and avirulence gene products confers rice blast resistance The EMBO Journal, 2000,19(15):4004-4014. [本文引用: 1]
PARKC H, SHIRSEKARG, BELLIZZIM, CHENS, SONGKUMARNP, XIEX, SHIX, NINGY, ZHOUB, SUTTIVIRIYAP, WANGM, UMEMURAK, WANGG L. The E3 ligase APIP10 connects the effector AvrPiz-t to the NLR receptor Piz-t in rice PLoS Pathogens, 2016,12(3):e1005529. [本文引用: 1]
FUJISAKIK, ABEY, ITOA, SAITOHH, YOSHIDAK, KANZAKIH, KANZAKIE, UTSUSHIH, YAMASHITAT, KAMOUNS, TERAUCHIR. Rice Exo70 interacts with a fungal effector, AVR-Pii, and is required for AVR-Pii-triggered immunity The Plant Journal, 2015,83(5):875-887. [本文引用: 1]
ASHKANIS, RAFIIM, SHABANIMOFRADM, GHASEMZADEHA, RAVANFARS A, LATIFM. Molecular progress on the mapping and cloning of functional genes for blast disease in rice (Oryza sativa L.): Current status and future considerations Critical Reviews in Biotechnology, 2016,36(2):353-367. [本文引用: 1]
SHARMAT, RAIA, GUPTAS, VIJAYANJ, DEVANNAB, RAYS. Rice blast management through host-plant resistance: Retrospect and prospects Agricultural Research, 2012,1(1):37-52. [本文引用: 1]
XIEZ, YANB X, SHOUJ Y, TANGJ, WANGX, ZHAIK R, LIUJ Y, LIQ, LUOM Z, DENGY W, HEZ H. A nucleotide-binding site-leucine-rich repeat receptor pair confers broad-spectrum disease resistance through physical association in rice Philosophical Transactions of the Royal Society B, 2019,374(1767):20180308. [本文引用: 1]
WANGB H, EBBOLED J, WANGZ H. The arms race between Magnaporthe oryzae and rice: Diversity and interaction of Avr and R genes Journal of Integrative Agriculture, 2017,16(12):2746-2760. [本文引用: 1]
ZHAOH J, WANGX Y, JIAY L, MINKENBERGB, WHEATLEYM, FANJ B, JIAM H, FAMOSOA, EDWARDSJ D, WAMISHEY, VALENTB, WANGG L, YANGY N. The rice blast resistance gene Ptr encodes an atypical protein required for broad-spectrum disease resistance Nature Communications, 2018,9(1):2039. [本文引用: 1]
FUKUOKAS, YAMAMOTOS I, MIZOBUCHIR, YAMANOUCHIU, ONOK, KITAZAWAN, YASUDAN, FUJITAY, NGUYENT T, KOIZUMIS, SUGIMOTOK, MATSUMOTOT, YANOM. Multiple functional polymorphisms in a single disease resistance gene in rice enhance durable resistance to blast Scientific Reports, 2014,4:4550. [本文引用: 1]
FUKUOKAS, SAKAN, KOGAH, ONOK, SHIMIZUT, EBANAK, HAYASHIN, TAKAHASHIA, HIROCHIKAH, OKUNOK, YANOM. Loss of function of a proline-containing protein confers durable disease resistance in rice Science, 2009,325(5943):998-1001. [本文引用: 1]
HAYASHIN, INOUEH, KATOT, FUNAOT, SHIROTAM, SHIMIZUT, KANAMORIH, YAMANEH, HAYANO-SAITOY, MATSUMOTOT, YANOM, TAKATSUJIH. Durable panicle blast-resistance gene Pb1 encodes an atypical CC-NBS-LRR protein and was generated by acquiring a promoter through local genome duplication The Plant Journal, 2010,64(3):498-510. [本文引用: 1]
XUX, HAYASHIN, WANGC T, FUKUOKAS, KAWASAKIS, TAKATSUJIH, JIANGC J. Rice blast resistance gene Pikahei-1(t), a member of a resistance gene cluster on chromosome 4, encodes a nucleotide-binding site and leucine-rich repeat protein Molecular Breeding, 2014,34(2):691-700. [本文引用: 1]
INUKAIT, NAGASHIMAS, KATOM. Pid3-I1 is a race-specific partial-resistance allele at the Pid3 blast resistance locus in rice Theoretical and Applied Genetics, 2019,132(2):395-404. [本文引用: 1]
VARIARM, CRUZCV, CARRILLOM, BHATTJ, SANGARR. Rice blast in India and strategies to develop durably resistant cultivars//Advances in Genetics, Genomics and Control of Rice Blast Disease Springer, 2009: 359-373. [本文引用: 2]
WANB J, LIUK, ZHAOS L, ZHUJ W, LIUY Y, ZHANGG Y, ZHUG Y, WANGA M, TANGH S, SUNM F, YANG H. Distribution of rice blast resistance genes Pi-ta, Pi-b, Pigm and Pi54 in backbone parent and their relationships with neck blast resistance Southwest China Journal of Agricultural Sciences, 2020,33(1):1-6. (in Chinese) [本文引用: 1]
ZHUY L, FANF J, XIEY L, WUY B, QIAOZ Y, ZHANGJ D. Detection and evaluation of blast resistance genes in late mature medium japonica lines in Jiangsu Province Jiangsu Agricultural Sciences, 2018,46(19):106-109. (in Chinese) [本文引用: 1]
JIANGH C, FENGY T, BAOL, LIX, GAOG J, ZHANGQ L, XIAOJ H, XUC G, HEY Q. Improving blast resistance of Jin 23B and its hybrid rice by marker-assisted gene pyramiding Molecular Breeding, 2012,30(4):1679-1688. [本文引用: 1]
XIAOW M, YANGQ Y, HUANGM, GUOT, LIUY Z, WANGJ F, YANGG L, ZHOUJ Y, YANGJ Y, ZHUX Y, CHENZ Q, WANGH. Improvement of rice blast resistance by developing monogenic lines, two-gene pyramids and three-gene pyramid through MAS Rice, 2019,12(1):78. [本文引用: 1]
HITTALMANIS, PARCOA, MEWT, ZEIGLERR, HUANGN. Fine mapping and DNA marker-assisted pyramiding of the three major genes for blast resistance in rice Theoretical and Applied Genetics, 2000,100(7):1121-1128. [本文引用: 1]
XIAON, WUY Y, PANC H, YUL, CHENY, LIUG Q, LIY H, ZHANGX X, WANGZ P, DAIZ Y, LIANGC Z, LIA H. Improving of rice blast resistances in japonica by pyramiding major R genes Frontiers in Plant Science, 2017,7:1918. [本文引用: 1]
WUY Y, XIAON, CHENY, YUL, PANC H, LIY H, ZHANGX X, HUANGN S, JIH J, DAIZ Y, CHENX J, LIA H. Comprehensive evaluation of resistance effects of pyramiding lines with different broad-spectrum resistance genes against Magnaporthe oryzae in rice (Oryza sativa L.) Rice, 2019,12(1):11. [本文引用: 7]
ZHONGZ H, CHENM L, LINL Y, HANY J, BAOJ D, TANGW, LINL L, LINY H, SOMAIR, LUL, et al. Population genomic analysis of the rice blast fungus reveals specific events associated with expansion of three main clades The ISME Journal, 2018,12(8):1867-1878. [本文引用: 2]
XIAON, WUY Y, WANGZ P, LIY H, PANC H, ZHANGX X, YUL, LIUG Q, ZHOUC H, JIH J, HUANGN S, JIANGM, DAIZ Y, LIA H. Improvement of seedling and panicle blast resistance in Xian rice varieties following Pish introgression Molecular Breeding, 2018,38:142. [本文引用: 1]
CHENB, ZHOUN B, GUOB W, HUANGD S, CHENZ P, HUAJ, HUOZ Y, ZHANGH C. Progress of “indica rice to japonica rice” in southern China Journal of Yangzhou University (Agricultural and Life Science Edition), 2017,38(1):67-72, 88. (in Chinese) [本文引用: 1]
YINM, LIUS W, CHUG, XUC M, WANGD Y, ZHANGX F, CHENS. Differences in yield and growth traits of different japonica varieties in the double cropping late season in the lower reaches of the Yangtze River Scientia Agricultura Sinica, 2020,53(5):890-903. (in Chinese) [本文引用: 1]
WUY Y, XIAON, YUL, PANC H, LIY H, ZHANGX X, LIUG Q, DAIZ Y, PANX B, LIA H. Combination patterns of major R genes determine the level of resistance to the M. oryzae in rice (Oryza sativa L.) PLoS ONE, 2015,10(6):e0126130. [本文引用: 2]
WANGX Q, DUH B, CHENX J, LIM Y, WANGJ N, XUZ W, FENGZ M, CHENZ X, ZUOS M. Analysis of blast resistant genes and neck blast resistance of japonica rice varieties/lines recently developed in Jiangsu Province Chinese Journal of Rice Science, 2020,34(5):413-424. (in Chinese) [本文引用: 1]
WUY Y, CHENY, PANC H, XIAON, YUL, LIY H, ZHANGX X, PANX B, CHENX J, LIANGC Z, DAIZ Y, LIA H. Development and evaluation of near-isogenic lines with different blast resistance alleles at the Piz locus in japonica rice from the lower region of the Yangtze River, China Plant Disease, 2017,101(7):1283-1291. [本文引用: 2]
PURIK D, SHRESTHAS M, CHHETRIG B K, JOSHIK D. Leaf and neck blast resistance reaction in tropical rice lines under green house condition Euphytica, 2009,165(3):523-532. [本文引用: 1]
LUY J, ZHENGK L. A simple method for isolation of rice DNA Chinese Journal of Rice Science, 1992,6(1):47-48. (in Chinese) [本文引用: 1]
POLANDJ A, BROWNP J, SORRELLSM E, JANNINKJ L. Development of high-density genetic maps for barley and wheat using a novel two-enzyme genotyping-by-sequencing approach PLoS ONE, 2012,7(2):e32253. [本文引用: 1]
KAWAHARAY, DELA BASTIDE M, HAMILTONJ P, KANAMORIH, MCCOMBIEW R, OUYANGS, SCHWARTZD C, TANAKAT, WUJ, ZHOUS, et al. Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data Rice, 2013,6(1):4. [本文引用: 1]
WALKERM A, PEDAMALLUC S, OJESINAA I, BULLMANS, SHARPET, WHELANC W, MEYERSONM. GATK PathSeq: A customizable computational tool for the discovery and identification of microbial sequences in libraries from eukaryotic hosts Bioinformatics, 2018,34(24):4287-4289. [本文引用: 1]
MACKILLD, BONMANJ. Inheritance of blast resistance in near-isogenic lines of rice Phytopathology, 1992,82(7):746-749. [本文引用: 1]
WUY Y, YUL, PANC H, DAIZ Y, LIY H, XIAON, ZHANGX X, JIH J, HUANGN S, ZHAOB H, et al. Development of near-isogenic lines with different alleles of Piz locus and analysis of their breeding effect under Yangdao 6 background Molecular Breeding, 2016,36(2):12. [本文引用: 1]
PENGH J, ZHANGJ, RAOZ W, PENGS Z, WUX L. Investigation on rice blast in different ecological areas Southwest China Journal of Agricultural Sciences, 1995,8(1):59-64. (in Chinese) [本文引用: 1]
DIVYAB, BISWASA, ROBINS, RABINDRANR, JOELA J. Gene interactions and genetics of blast resistance and yield attributes in rice (Oryza sativa L.) Journal of Genetics, 2014,93(2):415-424. [本文引用: 1]
CHAIPANYAC, TELEBANCO-YANORIAM J, QUIMEB, LONGYAA, KORINSAKS, KORINSAKS, TOOJINDAT, VANAVICHITA, JANTASURIYARATC, ZHOUB. Dissection of broad-spectrum resistance of the Thai rice variety Jao Hom Nin conferred by two resistance genes against rice blast Rice, 2017,10(1):18. [本文引用: 1]
CHENX L, JIAY L, JIAM H, PINSONS R M, WANGX Y, WUB M. Functional interactions between major rice blast resistance genes, Pi-ta and Pi-b, and minor blast resistance quantitative trait loci Phytopathology, 2018,108(9):1095-1103. [本文引用: 1]
LIW, DENGY W, NINGY S, HEZ H, WANGG L. Exploiting broad-spectrum disease resistance in crops: From molecular dissection to breeding Annual Review of Plant Biology, 2020,71:575-603. [本文引用: 1]
XUX, LVQ M, SHANGJ J, PANGZ Q, ZHOUZ Z, WANGJ, JIANGG H, TAOY, XUQ, LIX B, ZHAOX F, LIS G, XUJ C, ZHUL H. Excavation of Pid3 orthologs with differential resistance spectra to Magnaporthe oryzae in rice resource PLoS ONE, 2014,9(3):e93275. [本文引用: 1]
PRADHANS K, NAYAKD K, MOHANTYS, BEHERAL, BARIKS R, PANDITE, LENKAS, ANANDANA. Pyramiding of three bacterial blight resistance genes for broad-spectrum resistance in deepwater rice variety, Jalmagna Rice, 2015,8(1):51. [本文引用: 1]
 ,1,2, 肖宁1,2,4, 余玲1,2, 蔡跃1,2, 潘存红1,3, 李育红1,2, 张小祥1,2, 黄年生1,2, 季红娟1,2, 戴正元1,3, 李爱宏
,1,2, 肖宁1,2,4, 余玲1,2, 蔡跃1,2, 潘存红1,3, 李育红1,2, 张小祥1,2, 黄年生1,2, 季红娟1,2, 戴正元1,3, 李爱宏

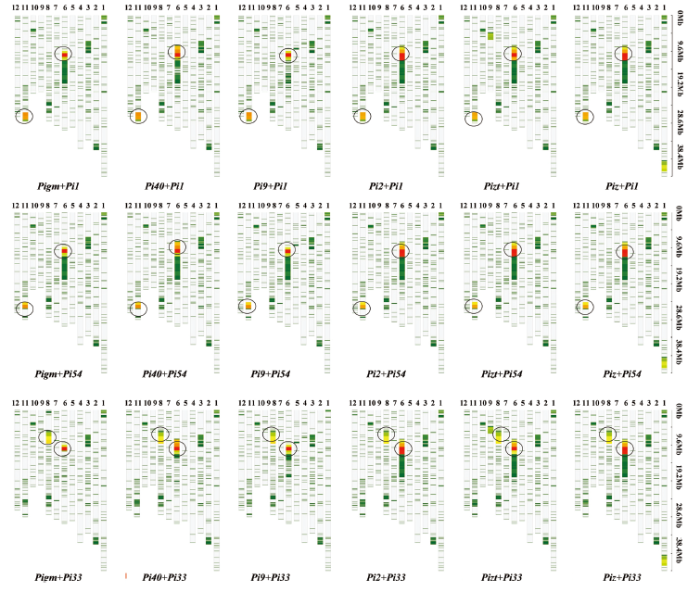 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT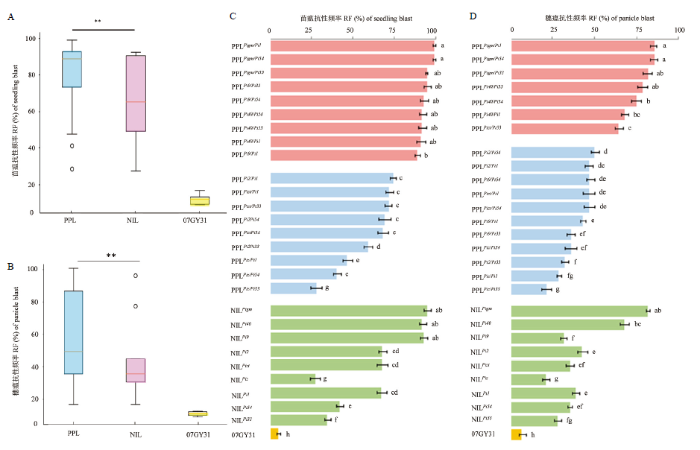 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT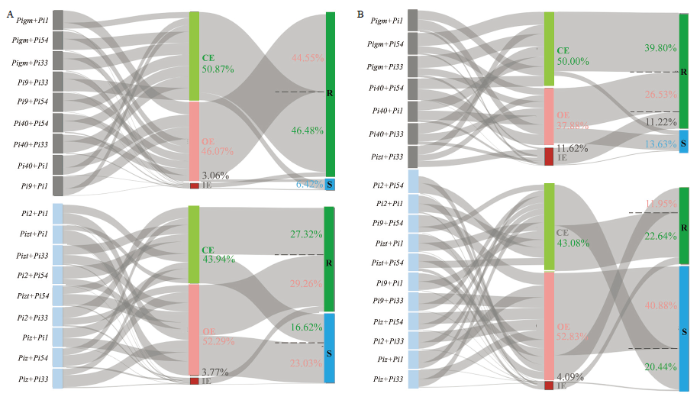 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT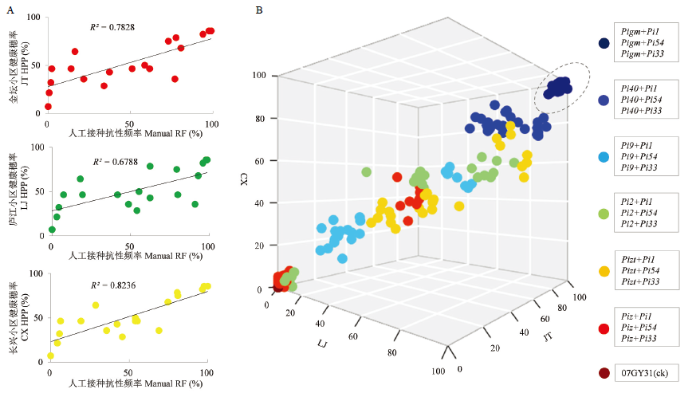 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}