Functional Identification of Grape Potassium Ion Transporter VviHKT1;7 Under Salt Stress
LIU Chuang,, GAO Zhen, YAO YuXin, DU YuanPeng,College of Horticultural Science and Engineering, Shandong Agricultural University/State Key Laboratory of Crop Biology, Tai’an 271018, Shandong
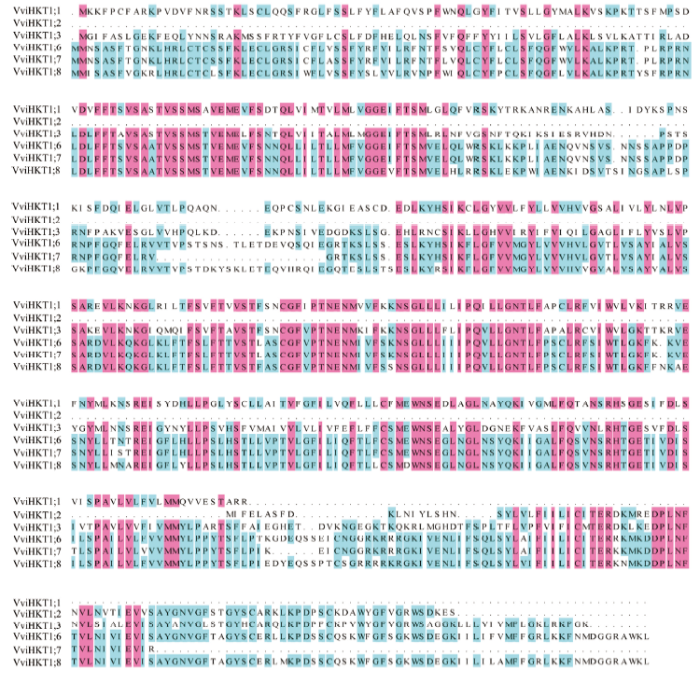
Abstract 【Objective】The aim of this study was to explore the role of VviHKT1;7 in the salt tolerance mechanism of grapes, so as to provide a theoretical reference for the subsequent cultivation of new salt-tolerant varieties. 【Method】DANMAN and MEGA software were used to analyze the biological information of VviHKT. The strongly salt resistant rootstocks SA15, SA17 and the commonly used rootstock 1103P tissue cultured seedlings were used as materials. Seedlings were treated under 100 mmol·L-1 NaCl for 0, 3, 6, 12, 24, 48 h, and the corresponding time of water treatment were taken as control. Real-time quantitative PCR (qRT-PCR) was used to detect the relative expression of HKT1 in the roots of grapes. VviHKT1;7 was cloned from SA17 cDNA and then linked with pRI101-AN-GFP, and the inflorescence of Arabidopsis thaliana was infected by Agrobacterium tumefaciens. Subsequently, T3 homozygous lines were screened out from resistant MS plates. Wild-type and transgenic Arabidopsis seeds were sowed on MS plates and MS plates (150 mmol·L-1NaCl added), their germination and growth were observed, and the root length and fresh weight were counted. The SA17 transgenic grape roots were obtained by Agrobacterium rhizogenes technology. After being treated with 100 mmol·L-1NaCl for 24 hours, the NMT in vivo physiological detector based on non-damaging micro-measurement technology was used to detect the net flow of Na+ and K+ instantaneous flow under salt stress in the roots of wild-type and transgenic grapes. 【Result】Multiple sequence alignment and phylogenetic tree analysis showed that VviHKT had high homology, among which the VviHKT1;7 open reading frame sequence length was 1 380 bp and it was the closest to VviHKT1;6. Salt stress significantly induced the expression of HKT1 gene in three varieties of grapes. Among them, the relative expression of VviHKT1;7 was up-regulated, which was still increased after long-term stress. The relative expression of VviHKT1;7 reached the peak at 6 or 12 h under salt stress, and its relative expression in SA17 and SA15 was significantly higher than 1103P. Results of germination and growth experiments in Arabidopsis showed that there was no significant difference between wild-type and transgenic Arabidopsis under normal conditions, but the germination rate, root length and fresh weight of transgenic Arabidopsis were significantly higher than those of wild type under salt stress. Fluorescence detection experiments showed that green fluorescence could be seen in the transgenic grape roots under fluorescence, rather than in the wild-type roots. Further, qRT-PCR results also showed that the relative expression of VviHKT1;7 in the transgenic grape roots was 20-folds higher than that in the wild-type roots. The results of ion flow rate detection showed that the net flow of Na + both in wild-type and transgenic roots showed efflux under normal conditions. Besides, no significant difference was found between wild-type and transgenic roots (208 and 205 pmol·cm-2·s-1) and the fluctuation range of ion flow rate in each time period was small. After salt stress, the Na+ net fluxes of them increased significantly, and the fluctuations in each time period also increased; the average net fluxes of wild-type and transgenic roots were 1 053 and 1 340 pmol·cm-2·s-1, respectively. Under normal conditions, the K+ absorption and efflux of the two roots were in a dynamic equilibrium. Salt stress significantly induced K+ efflux, and the efflux of K+in transgenic roots was significantly smaller than that in the wild type, which were 406 and 952 pmol·cm-2·s-1, respectively. The results indicated that the ability of removing Na+ and keeping K+ of transgenic roots was significantly greater than that of wild type. 【Conclusion】VviHKT1;7 played an important role in the response of grapes to salt stress, and the overexpression of this gene could improve the adaptability of Arabidopsis and grape roots under salt stress. Keywords:grape;VviHKT1;7;salt stress;transgene;functional identification
PDF (4302KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 刘闯, 高振, 姚玉新, 杜远鹏. 葡萄钾离子转运基因VviHKT1;7在盐胁迫下的功能鉴定[J]. 中国农业科学, 2021, 54(9): 1952-1963 doi:10.3864/j.issn.0578-1752.2021.09.012 LIU Chuang, GAO Zhen, YAO YuXin, DU YuanPeng. Functional Identification of Grape Potassium Ion Transporter VviHKT1;7 Under Salt Stress[J]. Scientia Acricultura Sinica, 2021, 54(9): 1952-1963 doi:10.3864/j.issn.0578-1752.2021.09.012
A:拟南芥种子在MS板上一周后的发芽状况;B:拟南芥种子在添加150 mmol·L-1 NaCl的MS板上一周后的发芽情况;C:NaCl胁迫后发芽率统计。**表示差异极显著(P<0.01);WT代表野生型,OE-1、OE-2、OE-3代表不同的转基因株系。下同 Fig. 5Germination of wild-type and transgenic Arabidopsis seeds
A: Germination status of Arabidopsis seeds after one week on MS plate; B: Germination of Arabidopsis seeds on MS plate with 150 mmol·L-1 NaCl added for one week; C: Statistics of germination rate after NaCl stress. ** indicates extremely significant difference (P<0.01); WT stands for wild type, OE-1, OE-2, and OE-3 stand for different transgenic lines. The same as below
JAMESR A, BLAKEC, BYRTC S, MUNNSR. Major genes for Na+ exclusion, Nax1 and Nax2 (wheat HKT1;4 and HKT1;5), decrease Na+ accumulation in bread wheat leaves under saline and waterlogged conditions Journal of Experimental Botany, 2011,62(8):2939-2947. [本文引用: 1]
LIUJ, CHAOT J, QiG M, QIG M, LIUY Z, HANY F. Booming development of Chinese grape industry Sino-Overseas Grapevine & Wine, 2020(1):1-8. (in Chinese) [本文引用: 1]
MAASE V, HOFFMANG J. Crop salt tolerance-current assessment Journal of the Irrigation and Drainage Division, 1977,103(2):115-134. [本文引用: 1]
CHINNUSAMYV, ZHUJ H, ZHUJ K. Salt stress signaling and mechanisms of plant salt tolerance Genetic Engineering, 2006,27:141-177. [本文引用: 1]
DEINLEINU, STEPHANA B, HORIET, LUOW, XUG H, SCHROEDERJ I. Plant salt-tolerance mechanisms Trends in Plant Science, 2014,19(6):371-379. [本文引用: 1]
NEUMANNP M. Chapter 2-recent advances in understanding the regulation of whole-plant growth inhibition by salinity, drought and colloid stress Advances in Botanical Research, 2011,57:33-48. [本文引用: 1]
QINL, KANGW H, QIY L, CAIA J. Effects of salt stress on mesophyll cell structures and photosynthetic characteristics in leaves of wine grape (Vitis spp.) Scientia Agricultura Sinica, 2012,45(20):4233-4241. (in Chinese) [本文引用: 1]
BABYT, COLLINSC, TYERMANS D, GILLIHAMM. Salinity negatively affects pollen tube growth and fruit set in grapevines and cannot be ameliorated by silicon American Journal of Enology & Viticulture, 2016,67(2):218-228. [本文引用: 1]
WALKERR R, CLINGELEFFERP R. Rootstock attributes and selection for Australian conditions Australian Viticulture, 2009,13(4):70-76. [本文引用: 1]
LIC, LIX J, HANZ, LIUL P, LIB. Research advances on effects of abiotic stress on photosynthesis of grape Shandong Agricultural Sciences, 2017,49(12):144-148. (in Chinese) [本文引用: 1]
WALKERR R, BLACKMORED H, CLINGELEFFERP R, CORRELLR L. Rootstock effects on salt tolerance of irrigated field-grown grapevines (Vitis vinifera L. cv. Sultana) 2. Ion concentrations in leaves and juice Australian Journal of Grape and Wine Research, 2004,10(2):90-99. [本文引用: 1]
STEVENSR M, HARVEYG, PARTINGTOND L. Irrigation of grapevines with saline water at different growth stages: Effects on leaf, wood and juice composition Australian Journal of Grape & Wine Research, 2011,17(2):239-248. [本文引用: 1]
FRANCISCOR, WALTERG, JULIANI S. Sodium-driven potassium uptake by the plant potassium transporter HKT1 and mutations conferring salt tolerance Science, 1995,270(5242):1660-1663. [本文引用: 1]
UOZUMIN, KIME J, RUBIOF, YAMAGUCHIT, MUTOS, TSUBOIA, BAKKERE P, NAKAMURAT, SCHROEDERJ I. The Arabidopsis HKT1 gene homolog mediates inward Na+ currents in Xenopus laevis oocytes and Na+ uptake in Saccharomyces cerevisiae Plant Physiology, 2000,122(4):1249-1259. [本文引用: 1]
HORIET, YOSHIDAK, NAKAYAMAH, YAMADAK, OIKIS, SHINMYOA. Two types of HKT transporters with different properties of Na+ and K+ transport in Oryza sativa The Plant Journal, 2001,27(2):129-138. [本文引用: 1]
GARCIADEBLáSB, SENNM E, BA?UELOSM A, RODR?GUEZ-NAVARRO A. Sodium transport and HKT transporters: The rice model Plant Journal, 2003,34(6):788-801. [本文引用: 1]
MASERP, ECKELMANB, VAIDYANATHANR, HORIET, FAIRBAURND J, KUBOM, YAMAGAMIM, YAMAGUCHIK, NISHIMURAM, UOZUMIN, ROBERYSONW, SUSSMANM R, SCHROEDERJ I. Altered shoot/root Na+ distribution and bifurcating salt sensitivity in Arabidopsis by genetic disruption of the Na+ transporter AtHKT1 FEBS Letters, 2002,531(2):157-161. [本文引用: 1]
SCHACHTMAND P, SCHROEDERJ I. Structure and transport mechanism of a high-affinity potassium uptake transporter from higher plants Nature, 1994,370(6491):655-658. [本文引用: 1]
WATERSS, GILLIHAMM, HRMOVAM. Plant high-affinity potassium (HKT) transporters involved in salinity tolerance: structural insights to probe differences in ion selectivity International Journal of Molecular Sciences, 2013,14(4):7660-7680. [本文引用: 1]
BEZOUWR F H M V, JANSSENE M, ASHRAFUZZAMANM, GHAHRAMANZADEHR, KILIANB, GRANERA, VISSERR G F, VANDER LINDEN C G. Shoot sodium exclusion in salt stressed barley (Hordeum vulgare L.) is determined by allele specific increased expression of HKT1;5 Journal of Plant Physiology, 2019,241:153029. [本文引用: 1]
SUZUKIK, YAMAJIN, COSTAA, OKUMAE, KOBAYASHIN I, KASHIWAGIT, KATSUHARAM, WANGC, TANOIK, MURATAY, SCHROEDERJ I, MAJ F, HORIET. OsHKT1;4-mediated Na+ transport in stems contributes to Na+ exclusion from leaf blades of rice at the reproductive growth stage upon salt stress BMC Plant Biology, 2016,16(1):22. [本文引用: 1]
WANGL, LIUY H, LID, FENGS J, YANGJ W, ZHANGJ J, ZHANGJ L, WANGD, GANY T. Improving salt tolerance in potato through overexpression of AtHKT1 gene BMC Plant Biology, 2019,19(1):357. [本文引用: 1]
FUQ Q. Salt tolerance identification and mechanism of hybrid rootstocks from ‘Zuo Shan 1’ [D]. Tai’an: Shandong Agricultural University, 2018. (in Chinese) [本文引用: 4]
WANGF P, ZHAOP P, ZHANGL, ZHAIH, DUY P. Functional characterization of WRKY46 in grape and its putative role in the interaction between grape and phylloxera (Daktulosphaira vitifoliae) Horticulture Research, 2019,6(1):803-814. [本文引用: 1]
GAOH B, ZHANGS J, SHENY B. Regurgitant from Orgyia ericae Germar induces calcium influx and accumulation of hydrogen peroxide in Ammopiptanthus mongolicus (Maxim. ex Kom.) Cheng f. cells Acta Ecologica Sinica, 2012,32(20):6520-6526. (in Chinese) [本文引用: 1]
ZHUJ K. Regulation of ion homeostasis under salt stress Current Opinion in Plant Biology, 2003,6(5):441-445. [本文引用: 1]
HAMAMOTOS, HORIET, HAUSERF, DEINLEINU, SCHROEDERJ, UOZUMIN. HKT transporters mediate salt stress resistance in plants: from structure and function to the field Current Opinion in Biotechnology, 2015,32:113-120. [本文引用: 1]
HENDERSONS W, DUNLEVYJ D, WUY, BLACKMORED H, WALKERR R, EDWARDSE J, GILLIHAMM, WALKERA R. Functional differences in transport properties of natural HKT1;1 variants influence shoot Na+ exclusion in grapevine rootstocks The New Phytologist, 2018,217(3):1113-1127. [本文引用: 1]
HAUSERF, HORIET. A conserved primary salt tolerance mechanism mediated by HKT transporters: A mechanism for sodium exclusion and maintenance of high K+/Na+ ratio in leaves during salinity stress Cell & Environment, 2010,33(4):552-565. [本文引用: 1]
WUY, HENDERSONS W, WEGES, ZHENGF, WALKERA R, WALKERR R, GILLIHAMM. The grapevine NaE sodium exclusion locus encodes sodium transporters with diverse transport properties and localisation Journal of Plant Physiology, 2020,246/247:153113. [本文引用: 3]
XUM, CHENC H, CAIH, WUL. Overexpression of PeHKT1;1 improves salt tolerance in Populus Genes, 2018,9(10):475. [本文引用: 2]
MIANA, OOMENR J, LSAYENKOVS, SENTENACH, MAATHUISF J, VéRYA A. Over-expression of an Na+ and K+ permeable HKT transporter in barley improves salt tolerance Plant Journal, 2011,68(3):468-479. [本文引用: 2]
GUOQ, MENGS, TAOS C, FENGJ, FANX Q, XUP, XUZ Z, SHENX L. Overexpression of a samphire high-affinity potassium transporter gene SbHKT1 enhances salt tolerance in transgenic cotton Acta Physiologiae Plantarum, 2020,42(3):36. [本文引用: 2]
ROMERO-ARANDAM R, GONZáLEZ-FERNáNDEZP, PéREZ-TIENDA J R, LóPEZ-DIAZM R, ESPINOSAJ, GRANUME, TRAVERSOJ á, PINEDAB, GARCIA-SOGOB, MORENOV, ASINSM J, BELVERA. Na+ transporter HKT1;2 reduces flower Na+ content and considerably mitigates the decline in tomato fruit yields under saline conditions Plant Physiology and Biochemistry, 2020,154:341-352. [本文引用: 2]
 ,, 高振, 姚玉新, 杜远鹏
,, 高振, 姚玉新, 杜远鹏

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT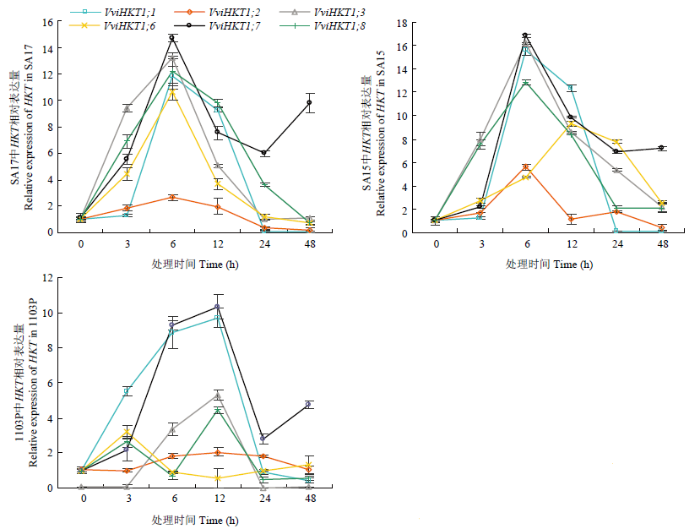 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT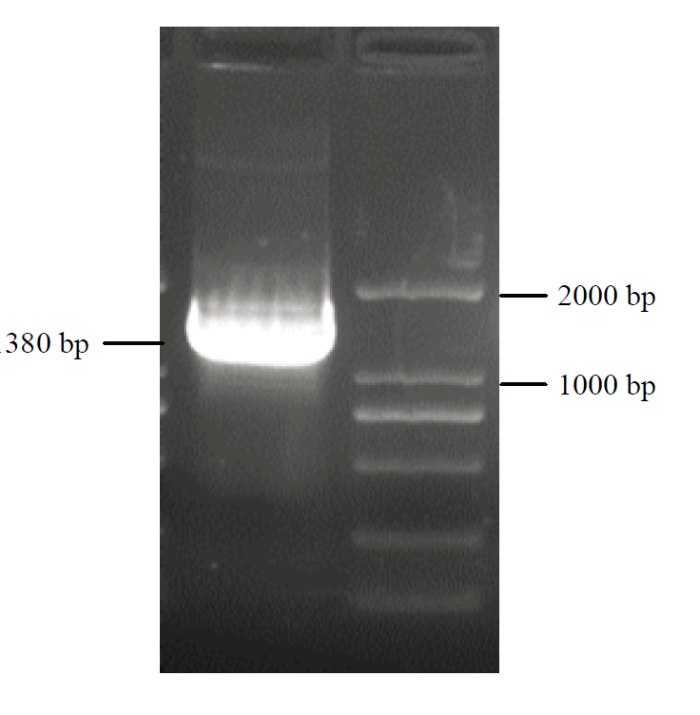 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT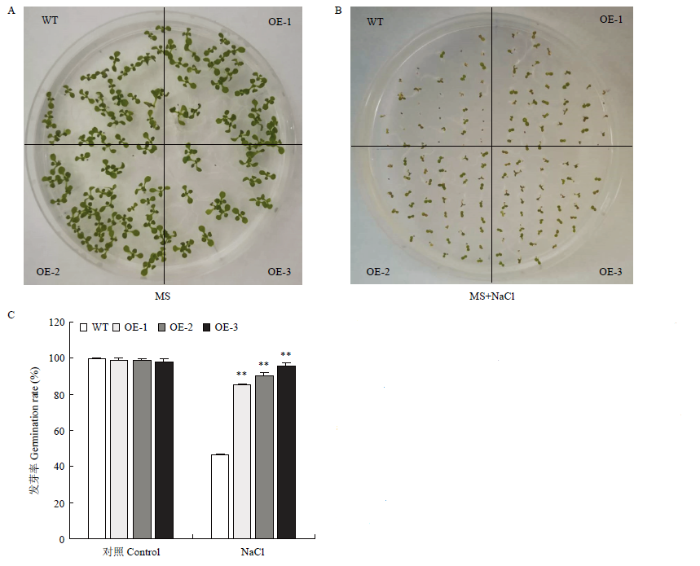 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT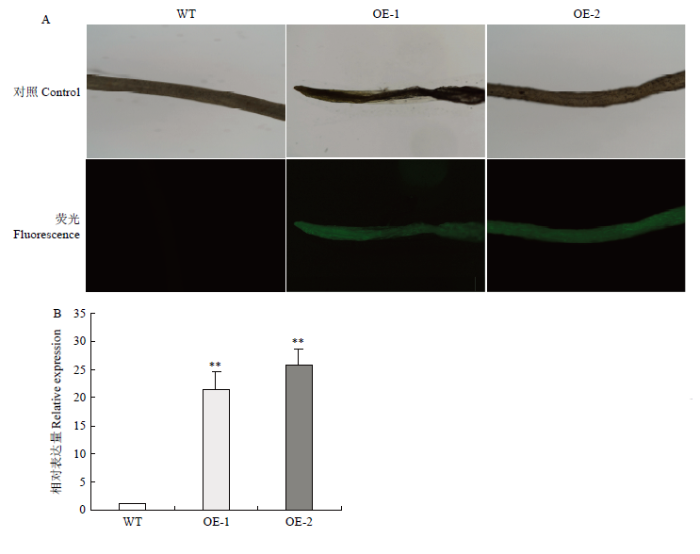 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT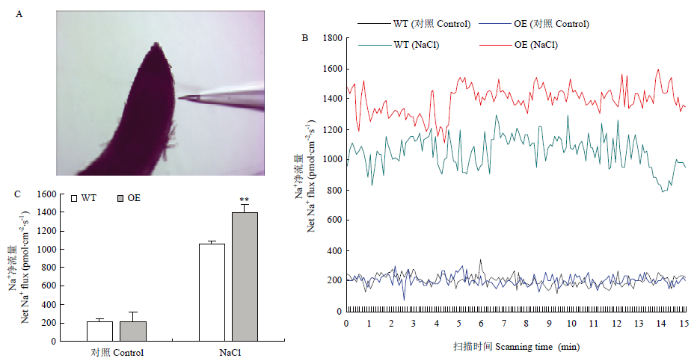 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT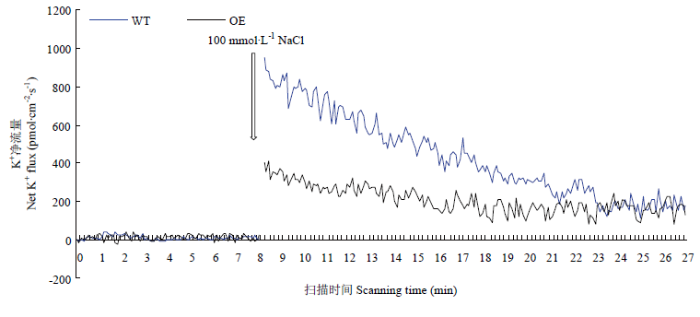 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}