Physiological Mechanisms of Abiotic Stress Priming Induced the Crops Stress Tolerance: A Review
WANG Xiao,, CAI Jian, ZHOU Qin, DAI TingBo, JIANG Dong,National Technique Innovation Center for Regional Wheat Production, Nanjing Agricultural University/Key Laboratory of Crop Physiology Ecology and Production Management, Ministry of Agriculture and Rural Affairs, Nanjing 210095
Abstract Abiotic stress factors, including heat stress, cold stress, drought stress, waterlogging stress etc., are the limiting factors for plant growth and crop production. Moreover, the frequency, extent and duration of abiotic stresses have predicted to be increased with global climate change. Therefore, improving crops resistance to abiotic stress or finding strategy to reduce the adverse effects of abiotic stress on crop yield and quality is of great significance for ensuring stable crop production and food security. “Priming” by exposing plants to moderate stress in the early growth stage can induce plant resistance to a later severe stress episode which happened during critical stage of the plant growth. The priming is an adaptive strategy that primed plants could effectively mount a faster and/or stronger defense response and actively improve the defensive capacity of plants under stress, compared with non-primed plants. According to the reoccurred stress types, the priming can be generally separated to four types, including in-generational stress tolerance (the priming stimulus and the stress occurred at later stage are the same stress type), intra-generational cross tolerance (the priming stimulus and the stress occurred at later stage are different stress types), transgenerational stress tolerance (the priming stimulus and the stress occurred at next generations are the same stress type), and transgenerational cross tolerance (the priming stimulus and the stress occurred at next generations are different stress types). In this review, the main physiological mechanisms were discussed, including plant photosynthetic response mechanism, antioxidant mechanism, and osmotic regulation, signal transduction mechanisms (plant hormones, Ca2+, hydrogen peroxide, nitric oxide, etc.), and epigenetic modification mechanisms (DNA methylation, histone modification, etc.). Then, the perspectives for the further research on understanding the underlying mechanisms of stress priming and the application of priming effects in the crop production were suggested. After revealing the mechanisms of priming, the candidate genes and proteins which play key regulatory roles in the acquisition of crop stress tolerance are found. And then, we can stimulate the related genes and protein expression which can actively induce the formation of stress tolerance in the critical crops growth stage, thereby effectively alleviating the adverse effects of abiotic stress on crop yield during the critical stage, which is meaningful for the crop production. Keywords:abiotic stress;priming;intra-generational priming;transgenerational priming;physiological mechanisms;signal transduction mechanisms
PDF (1536KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 王笑, 蔡剑, 周琴, 戴廷波, 姜东. 非生物逆境锻炼提高作物耐逆性的生理机制研究进展[J]. 中国农业科学, 2021, 54(11): 2287-2301 doi:10.3864/j.issn.0578-1752.2021.11.004 WANG Xiao, CAI Jian, ZHOU Qin, DAI TingBo, JIANG Dong. Physiological Mechanisms of Abiotic Stress Priming Induced the Crops Stress Tolerance: A Review[J]. Scientia Acricultura Sinica, 2021, 54(11): 2287-2301 doi:10.3864/j.issn.0578-1752.2021.11.004
拟南芥对高温的跨代适应性可以持续至少2个世代。研究表明,F0及F1代经历过高温胁迫的拟南芥植株,在F3代高温处理下经过锻炼的植株生物量是未经锻炼植株的5倍[84]。传代记忆机制目前在模式植物拟南芥上的研究较为深入。组蛋白去甲基化酶、染色质重塑因子、转录因子、泛素连接酶和小分子RNAs共同组成的复杂的表观调控网络参与了植物对高温的传代记忆[85]。高温能激活热激转录因子heat shock transcription factor A2 (HsfA2)。HsfA2能直接结合H3K27me3去甲基化酶Relative of early flowering 6 (REF6)和染色质重塑因子Brahma (BRM) 启动子上的热激转录元件从而激活REF6和BRM的表达。REF6和BRM的上调表达反过来降低HSFA2位点上的H3K27me3修饰水平,从而降低H3K27me3对HSFA2转录的抑制作用。因此,REF6和HSFA2形成了一个正向反馈循环途径来维持植物对高温的传代记忆[85]。在小麦灌浆期进行高温锻炼获得子代在花后遭遇高温胁迫,与未锻炼的植株相比,锻炼的植株具有较高的籽粒产量,叶片光合能力及抗氧化酶活性[86]。通过转录组分析发现经过高温锻炼的小麦后代植株中赖氨酸特异性组蛋白去甲基化酶1(LSD1)的基因诱导表达,与光合作用、能量产生以及蛋白质转运和存储相关蛋白的编码基因表达趋势一致,说明跨代高温记忆可能是由表观遗传的改变和信号转导来诱发的,从而激发了热胁迫相关防御机制[86]。
3.2 低温跨代效应及其生理机制
低温会诱导跨代记忆的产生,其中一种典型的跨代低温记忆现象为“春化作用”。春化作用是植物延长低温处理促进开花的过程,一旦分生组织受到长时间的低温处理,他们就会记住春化效应,并且这种记忆是稳定的。拟南芥、小麦等一年生作物通常会经历春化作用,长时间暴露于低温环境可以通过H3K27me3甲基化修饰触发FLC(FLOWERING LOCUS C)基因的表观遗传沉默,使植物能在春季开花,这种状态一直持续到胚胎发育,胚胎发育早期抑制性修饰逐渐消失,活跃染色质标记H3K36me3建立,重新激活FLC基因[87]。目前已有综述概述了拟南芥以及温带禾本科植物(小麦、大麦以及短柄草)中春化作用的分子遗传调控网络、表观遗传修饰与核心蛋白修饰介导的记忆机制,春化作用的分子与表观遗传控制机理在双子叶植物(如拟南芥)和单子叶植物(如小麦)完全不同[88,89,90,91]。在小麦和大麦等温带作物中,春化促进开花途径包括VRN1、VRN2、VRN3和VRN-D4等春化基因的调控,其中VRN1编码一个类似FRUITFULL的MADS-box转录因子,在春化过程中起到至关重要的促进作用。在低温处理前,H3K27me3标记在TaVRN1及其大麦同源物中富集,这2个基因都受到抑制,冷诱导这2个基因都与H3K27me3的减少和H3K4me3的增加有关[92]。但低温处理的种子后代对低温胁迫下植株耐性的影响及其生理机制尚未有报道。
WOLLENWEBERB, PORTERJ R, SCHELLBERGJ. Lack of interaction between extreme high-temperature events at vegetative and reproductive growth stages in wheat Journal of Agronomy and Crop Science, 2003,189(3):142-150. DOI:10.1046/j.1439-037X.2003.00025.xURL [本文引用: 1]
STOCKERT F, QIND, PLATTNERG K, TIGNORM M, ALLENS K, BOSCHUNGJ, NAUELSA, XIAY, BEXV, MIDGLEYP M. Climate Change 2013: The physical science basis. contribution of working group I to the fifth assessment report of IPCC the intergovernmental panel on climate change Computational Geometry, 2013,18(2):95-123. DOI:10.1016/S0925-7721(01)00003-7URL [本文引用: 1]
TRENBERTHK E. Changes in precipitation with climate change Climate Research, 2011,47:123-138. DOI:10.3354/cr00953URL [本文引用: 1]
FIELDC, BARROSV, DOKKEND, MACHK, MASTRANDREAM, BILIRT, CHATTERJEEM, EBIK, ESTRADAY, GENOVAR. IPCC, 2014: Climate change 2014: impacts, adaptation, and vulnerability. Part a: global and sectoral aspects//Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, and New York, 2014: 1132. [本文引用: 1]
TAIA P, MARTINM V, HEALDC L. Threat to future global food security from climate change and ozone air pollution Nature Climate Change, 2014(9), 4:817-821. DOI:10.1038/nclimate2317URL [本文引用: 1]
ASSENGS, FOSTERI, TURNERN C. The impact of temperature variability on wheat yields Global Change Biology, 2011,17(2):997-1012. DOI:10.1111/gcb.2010.17.issue-2URL [本文引用: 1]
GUPTAR, GOPALR, JATM, JATR, SIDHUH, MINHASP, MALIKR. Wheat productivity in Indo-Gangetic plains of India during 2010: Terminal heat effects and mitigation strategies Private Circulation Only Conservation Agriculture, 2010,14:1-4. [本文引用: 1]
DARYANTOS, WANGL, JACINTHEP-A. Global synthesis of drought effects on maize and wheat production PLoS ONE, 2016,11(5):e0156362. DOI:10.1371/journal.pone.0156362URL [本文引用: 1]
KAURG, SINGHG, MOTAVALLIP P, NELSONK A, ORLOWSKIJ M, GOLDENB R. Impacts and management strategies for crop production in waterlogged or flooded soils: A review Agronomy Journal, 2020,112(3):1475-1501. DOI:10.1002/agj2.v112.3URL [本文引用: 1]
BRUCET J A, MATTHESM C, NAPIERJ A, PICKETTJ A. Stressful “memories” of plants: Evidence and possible mechanisms Plant Science, 2007,173(6):603-608. DOI:10.1016/j.plantsci.2007.09.002URL [本文引用: 2]
BALMERA, PASTORV, GAMIRJ, FLORSV, MAUCH-MANIB. The ‘prime-ome’: Towards a holistic approach to priming Trends in Plant Science, 2015,20(7):443-452. DOI:10.1016/j.tplants.2015.04.002URL [本文引用: 3]
MOLINIERJ, RIESG, ZIPFELC, HOHNB. Transgeneration memory of stress in plants Nature, 2006,442(7106):1046-1049. DOI:10.1038/nature05022URL [本文引用: 1]
L?MKEJ, B?URLEI. Epigenetic and chromatin-based mechanisms in environmental stress adaptation and stress memory in plants Genome Biology, 2017,18(1):124. DOI:10.1186/s13059-017-1263-6URL [本文引用: 1]
ENGELBERTHJ, ALBORNH T, SCHMELZE A, TUMLINSONJ H. Airborne signals prime plants against insect herbivore attack Proceedings of the National Academy of Sciences of the USA, 2004,101(6):1781-1785. [本文引用: 1]
TONJ, D’ALESSANDROM, VIOLAINEJOURDIE1 G J, DANIELLEKARLEN1 M H, MAUCH-MANIB, TURLINGS1T C J. Priming by airborne signals boosts direct and indirect resistance in maize The Plant Journal, 2007,49(1):16-26. DOI:10.1111/tpj.2007.49.issue-1URL [本文引用: 1]
BECKERSG J, JASKIEWICZM, LIUY, UNDERWOODW R, HES Y, ZHANGS, CONRATHU. Mitogen-activated protein kinases 3 and 6 are required for full priming of stress responses in Arabidopsis thaliana The Plant Cell, 2009,21(3):944-953. DOI:10.1105/tpc.108.062158URL [本文引用: 1]
WANGX, LIUF, JIANGD. Priming: A promising strategy for crop production in response to future climate Journal of Integrative Agriculture, 2017,16(12):60345-60347. [本文引用: 1]
CHINNUSAMYV, ZHUJ K. Epigenetic regulation of stress responses in plants Current Opinion in Plant Biology, 2009,12(2):133-139. DOI:10.1016/j.pbi.2008.12.006URL [本文引用: 4]
MIGICOVSKYZ, YAOY, KOVALCHUKI. Transgenerational phenotypic and epigenetic changes in response to heat stress in Arabidopsis thaliana Plant Signaling and Behavior, 2014,9(2):e27971. DOI:10.4161/psb.27971URL [本文引用: 1]
HAUSERM T, AUFSATZW, JONAKC, LUSCHNIGC. Transgenerational epigenetic inheritance in plants Biochimica et Biophysica Acta (BBA)-Gene Regulatory Mechanisms, 2011,1809(8):459-468. [本文引用: 1]
ZHANGX, ZHOUQ, WANGX, CAIJ, DAIT, CAOW, JIANGD. Physiological and transcriptional analyses of induced post-anthesis thermo-tolerance by heat-shock pretreatment on germinating seeds of winter wheat Environmental and Experimental Botany, 2016,131:181-189. DOI:10.1016/j.envexpbot.2016.08.002URL [本文引用: 2]
WANGX, CAIJ, JIANGD, LIUF, DAIT, CAOW. Pre-anthesis high-temperature acclimation alleviates damage to the flag leaf caused by post-anthesis heat stress in wheat Journal of Plant Physiology, 2011,168(6):585-593. DOI:10.1016/j.jplph.2010.09.016URL [本文引用: 3]
WANGX, CAIJ, LIUF, DAIT, CAOW, WOLLENWEBERB, JIANGD. Multiple heat priming enhances thermo-tolerance to a later high temperature stress via improving subcellular antioxidant activities in wheat seedlings Plant Physiology and Biochemistry, 2013,74:185-192. DOI:10.1016/j.plaphy.2013.11.014URL [本文引用: 3]
WANGX, CAIJ, LIUF, JINM, YUH, JIANGD, WOLLENWEBERB, DAIT, CAOW. Pre-anthesis high temperature acclimation alleviates the negative effects of post-anthesis heat stress on stem stored carbohydrates remobilization and grain starch accumulation in wheat Journal of Cereal Science, 2012,55(3):331-336. DOI:10.1016/j.jcs.2012.01.004URL [本文引用: 1]
CHARNGY Y, LIUH C, LIUN Y, CHIW T, WANGC N, CHANGS H, WANGT T. A heat-inducible transcription factor, HsfA2, is required for extension of acquired thermotolerance in Arabidopsis Plant Physiology, 2007,143(1):251-262. DOI:10.1104/pp.106.091322URL [本文引用: 1]
SEDAGHATMEHRM, MUELLER-ROEBERB, BALAZADEHS. The plastid metalloprotease FtsH6 and small heat shock protein HSP21 jointly regulate thermomemory in Arabidopsis Nature Communications, 2016,7:12439. DOI:10.1038/ncomms12439URL [本文引用: 1]
LIQ Z, LIC W, YANGT W. Stress response and memory mediated by DNA methylation in plants Plant Physiology Journal, 2014,50(6):725-734. (in Chinese) [本文引用: 1]
STIEFA, ALTMANNS, HOFFMANNK, PANTB D, SCHEIBLEW R, B?URLEI. Arabidopsis miR156 regulates tolerance to recurring environmental stress through SPL transcription factors The Plant Cell, 2014,26(4):1792-1807. DOI:10.1105/tpc.114.123851URL [本文引用: 1]
LINGY, SERRANON, GAOG, ATIAM, MOKHTARM, WOOY H, BAZINJ, VELUCHAMYA, BENHAMEDM, CRESPIM, GEHRINGC, REDDYA S N, MAHFOUZM M. Thermopriming triggers splicing memory in Arabidopsis Journal of Experimental Botany, 2018,69(10):2659-2675. DOI:10.1093/jxb/ery062URL [本文引用: 1]
WANGW, WANGX, ZHANGJ, HUANGM, CAIJ, ZHOUQ, DAIT, JIANGD. Salicylic acid and cold priming induce late-spring freezing tolerance by maintaining cellular redox homeostasis and protecting photosynthetic apparatus in wheat Plant Growth Regulation, 2020,90(1):109-121. DOI:10.1007/s10725-019-00553-8URL [本文引用: 2]
LIX, CAIJ, LIUF, DAIT, CAOW, JIANGD. Cold priming drives the sub-cellular antioxidant systems to protect photosynthetic electron transport against subsequent low temperature stress in winter wheat Plant Physiology and Biochemistry, 2014,82:34-43. DOI:10.1016/j.plaphy.2014.05.005URL [本文引用: 2]
JIANL S, LUC F, LIJ H. Increment of chilling tolerance and its physiological basis in chilling-sensitive corn sprouts and tomato seedlings after cold-hardening at optimum temperatures Acta Agronomica Sinica, 2005,31(8):971-976. (in Chinese) [本文引用: 2]
WANGW, WANGX, HUANGM, CAIJ, ZHOUQ, DAIT, CAOW, JIANGD. Hydrogen peroxide and abscisic acid mediate salicylic acid-induced freezing tolerance in wheat Frontiers in Plant Science, 2018,9:1137. DOI:10.3389/fpls.2018.01137URL [本文引用: 1]
ZUTHERE, SCHAARSCHMIDTS, FISCHERA, ERBANA, PAGTERM, MUBEENU, GIAVALISCOP, KOPKAJ, SPRENGERH, HINCHAD K. Molecular signatures associated with increased freezing tolerance due to low temperature memory in Arabidopsis Plant Cell and Environment, 2019,42(3):854-873. [本文引用: 1]
ZHAOM G, CHENL, ZHANGL L, ZHANGW H. Nitric reductase- dependent nitric oxide production is involved in cold acclimation and freezing tolerance in Arabidopsis Plant Physiology, 2009,151(2):755-767. DOI:10.1104/pp.109.140996URL [本文引用: 1]
HUANGG B, ZHANGX H, YANGS L, LIJ Y, XUC H, RONGZ Y, YANGL Y, GONGM. Involvement of osmotic regulation in enhancement of drought resistance in tobacco (Nicotiana tabacum L.) plants through circular drought-hardening Plant Physiology Journal, 2012,48(5):465-471. (in Chinese) [本文引用: 1]
SELOTED S, KHANNA-CHOPRAR. Drought acclimation confers oxidative stress tolerance by inducing co-ordinated antioxidant defense at cellular and subcellular level in leaves of wheat seedlings Physiologia Plantarum, 2006,127(3):494-506. DOI:10.1111/j.1399-3054.2006.00678.xURL [本文引用: 1]
WANGX, VIGNJEVICM, JIANGD, JACOBSENS, WOLLENWEBERB. Improved tolerance to drought stress after anthesis due to priming before anthesis in wheat (Triticum aestivum L.) var. Vinjett Journal of Experimental Botany, 2014,65(22):6441-6456. DOI:10.1093/jxb/eru362URL [本文引用: 3]
BAHUGUNAR N, TAMILSELVANA, MUTHURAJANR, SOLISC A, JAGADISHS V K. Mild preflowering drought priming improves stress defences, assimilation and sink strength in rice under severe terminal drought Functional Plant Biology, 2018,45(8):827-839. DOI:10.1071/FP17248URL [本文引用: 1]
CHENX Y, LUOY P. Study on the compensatory effect of rewatering during the flowering stage after previous water stress in winter wheat Acta Agronomica Sinica, 2001,27(4):512-516. (in Chinese) [本文引用: 1]
CHENX Y, LUOY P. Compensatory effects of water-recovery during different growth durations on winter wheat under water stress Chinese Journal of Eco-Agriculture, 2002,10(1):35-37. (in Chinese) [本文引用: 1]
WANGX, MAOZ, ZHANGJ, HEMATM, HUANGM, CAIJ, ZHOUQ, DAIT, JIANGD. Osmolyte accumulation plays important roles in the drought priming induced tolerance to post-anthesis drought stress in winter wheat (Triticum aestivum L.) Environmental and Experimental Botany, 2019,166:103804. DOI:10.1016/j.envexpbot.2019.103804URL [本文引用: 1]
LIP, YANGH, WANGL, LIUH, HUOA H, ZHANGC, LIUA, ZHUA, HUJ, LINY. Physiological and transcriptome analyses reveal short-term responses and formation of memory under drought stress in rice Frontiers in Genetics, 2019,10:55. DOI:10.3389/fgene.2019.00055URL [本文引用: 3]
HARBA, KRISHNANA, AMBAVARAMM M, PEREIRAA. Molecular and physiological analysis of drought stress in Arabidopsis reveals early responses leading to acclimation in plant growth Plant Physiology, 2010,154(3):1254-1271. DOI:10.1104/pp.110.161752URL [本文引用: 1]
VIRLOUVETL, FROMMM. Physiological and transcriptional memory in guard cells during repetitive dehydration stress New Phytologist, 2015,205(2):596-607. DOI:10.1111/nph.2014.205.issue-2URL [本文引用: 1]
LIUN, AVRAMOVAZ. Molecular mechanism of the priming by jasmonic acid of specific dehydration stress response genes in Arabidopsis Epigenetics and Chromatin, 2016,9(1):8. DOI:10.1186/s13072-016-0057-5URL [本文引用: 2]
WANGX, LIQ, XIEJ, HUANGM, CAIJ, ZHOUQ, DAIT, JIANGD. Abscisic acid and jasmonic acid are involved in drought priming- induced tolerance to drought in wheat The Crop Journal, 2020,9(1):120-132. DOI:10.1016/j.cj.2020.06.002URL [本文引用: 1]
DUANH, LIJ, ZHUY, JIAW, WANGH, JIANGL, ZHOUY. Responsive changes of DNA methylation in wheat (Triticum aestivum) under water deficit Scientific Reports, 2020,10(1):7938. DOI:10.1038/s41598-020-64660-7URL [本文引用: 1]
DINGY, FROMMM, AVRAMOVAZ. Multiple exposures to drought ‘train’ transcriptional responses in Arabidopsis Nature Communications, 2012,3:740. DOI:10.1038/ncomms1732URL [本文引用: 1]
KIMJ M, TOT K, ISHIDAJ, MATSUIA, KIMURAH, SEKIM. Transition of chromatin status during the process of recovery from drought stress in Arabidopsis thaliana Plant and Cell Physiology, 2012,53(5):847-856. DOI:10.1093/pcp/pcs053URL [本文引用: 1]
LIC, JIANGD, WOLLENWEBERB, LIY, DAIT, CAOW. Waterlogging pretreatment during vegetative growth improves tolerance to waterlogging after anthesis in wheat Plant Science, 2011,180(5):672-678. DOI:10.1016/j.plantsci.2011.01.009URL [本文引用: 2]
LIC Y, CAIJ, JIANGD, DAIT B, CAOW X. Effects of hardening by pre-anthesis waterlogging on grain yield and quality of post- anthesis waterlogged wheat (Triticum aestivum L. cv Yangmai 9) Acta Ecologica Sinica, 2011,31(7):1904-1910. (in Chinese) [本文引用: 1]
KATO-NOGUCHIH. Anoxia tolerance in rice roots acclimated by several different periods of hypoxia Journal of Plant Physiology, 2003,160(5):565-568. DOI:10.1078/0176-1617-00868URL [本文引用: 2]
GARNCZARSKAM, RATAJCZAKL. Hypoxia induces anoxia tolerance in roots and shoots of lupine seedlings Acta Physiologiae Plantarum, 2003,25(1):47-53. DOI:10.1007/s11738-003-0035-yURL [本文引用: 2]
SAGLIOP H, DREWM C, PRADETA. Metabolic acclimation to anoxia induced by low (2-4 kpa partial-pressure) oxygen pretreatment (hypoxia) in root-tips of Zea-mays Plant Physiology, 1988,86(1):61-66. DOI:10.1104/pp.86.1.61URL [本文引用: 2]
WANGX, HUANGM, ZHOUQ, CAIJ, DAIT, CAOW, JIANGD. Physiological and proteomic mechanisms of waterlogging priming improves tolerance to waterlogging stress in wheat (Triticum aestivum L.) Environmental and Experimental Botany, 2016,132:175-182. DOI:10.1016/j.envexpbot.2016.09.003URL [本文引用: 1]
ZHOUR G, FANZ H, LIX Z, WANGZ W, HANW. The effect of heat acclimation on cellular membrane thermostability in wheat Chinese Journal of Pesticide Science, 1993,10(1):33-37. (in Chinese) [本文引用: 1]
VANBUER J, CVETKOVICJ, BAIERM. Cold regulation of plastid ascorbate peroxidases serves as a priming hub controlling ROS signaling in Arabidopsis thaliana. BMC Plant Biology, 2016,16(1):163-163. DOI:10.1186/s12870-016-0856-7URL [本文引用: 1]
LEUENDORFJ E, FRANKM, SCHMüLLINGT. Acclimation, priming and memory in the response of Arabidopsis thaliana seedlings to cold stress Scientific Reports, 2020,10(1):689. DOI:10.1038/s41598-019-56797-xURL [本文引用: 1]
LIT H, WANGX, CAIJ, ZHOUQ, DAIT B, JIANGD. Comprehensive evaluation of drought priming on plant tolerance in different wheat cultivars Journal of Triticeae Crops, 2018,38(1):65-73. (in Chinese) [本文引用: 1]
MAHAJANS, TUTEJAN. Cold, salinity and drought stresses: An overview, Archives of Biochemistry and Biophysics 2005,444(2):139-158. [本文引用: 1]
KANGJ H, WUH L, HUANGL D. Cross adaptation of stress on maize seedlings under drought induced Agricultural Research in the Arid Areas, 2008(6):143-148. (in Chinese) [本文引用: 1]
WANGX, VIGNJEVICM, LIUF, JACOBSENS, JIANGD, WOLLENWEBERB. Drought priming at vegetative growth stages improves tolerance to drought and heat stresses occurring during grain filling in spring wheat Plant Growth Regulation, 2015,75(3):677-687. DOI:10.1007/s10725-014-9969-xURL [本文引用: 1]
ZHANGX, WANGX, ZHUANGL, GAOY, HUANGB. Abscisic acid mediation of drought priming-enhanced heat tolerance in tall fescue (Festuca arundinacea) and Arabidopsis Physiologia Plantarum, 2019,167(4):488-501. DOI:10.1111/ppl.v167.4URL [本文引用: 1]
ZHANGX, XUY, HUANGB. Lipidomic reprogramming associated with drought stress priming enhanced heat tolerance in tall fescue (Festuca arundinacea) Plant, Cell and Environment, 2019,42(3):947-958. DOI:10.1111/pce.v42.3URL [本文引用: 1]
SIMINOVITCHD. Twenty-four-hour induction of freezing and drought tolerance in plumules of winter rye seedlings by desiccation stress at room temperature in the dark Plant Physiology, 1982,69(1):250-255. DOI:10.1104/pp.69.1.250URL [本文引用: 2]
AROCAR, IRIGOYENJ J, SáNCHEZ-DíAZM J P P. Drought enhances maize chilling tolerance. II. Photosynthetic traits and protective mechanisms against oxidative stress Physiologia Plantarum, 2003,117(4):540-549. DOI:10.1034/j.1399-3054.2003.00065.xURL [本文引用: 2]
LIX, TOPBJERGH, JIANGD, LIUF. Drought priming at vegetative stage improves the antioxidant capacity and photosynthesis performance of wheat exposed to a short-term low temperature stress at jointing stage Plant and Soil, 2015,393:307-318. DOI:10.1007/s11104-015-2499-0URL [本文引用: 1]
HX, ZL, ZT, FH, XL. Metabolomic analyses reveal substances that contribute to the increased freezing tolerance of alfalfa (Medicago sativa L.) after continuous water deficit BMC Plant Biology, 2020,20(1):15. DOI:10.1186/s12870-019-2233-9URL [本文引用: 2]
SIT, WANGX, ZHAOC, HUANGM, CAIJ, ZHOUQ, DAIT, JIANGD. The role of hydrogen peroxide in mediating the mechanical wounding-induced freezing tolerance in wheat Frontiers in Plant Science, 2018,9:327-327. DOI:10.3389/fpls.2018.00327URL [本文引用: 1]
MEIY Q, SONGS Q. Response to temperature stress of reactive oxygen species scavenging enzymes in the cross-tolerance of barley seed germination Journal of Zhejiang University Science B, 2010,11(12):965-972. DOI:10.1631/jzus.B1000147URL [本文引用: 1]
LIS L, XIAY Z, SUNZ Q. Effects of cold-shock on the growth and flower bud differentiation of tomato seedlings under high temperature stress Chinese Journal of Applied Ecology, 2016,27(2):477-483. [本文引用: 1]
KUZNETSOVV, RAKITINV, ZHOLKEVICHV. Effects of preliminary heat‐shock treatment on accumulation of osmolytes and drought resistance in cotton plants during water deficiency Physiologia Plantarum, 2002,107(4):399-406. DOI:10.1034/j.1399-3054.1999.100405.xURL [本文引用: 1]
GAOJ, LUOQ, SUNC, HUH, WANGF, TIANZ, JIANGD, CAOW, DAIT. Low nitrogen priming enhances photosynthesis adaptation to water-deficit stress in winter wheat (Triticum aestivum L.) seedlings Frontiers in Plant Science, 2019,10:818. DOI:10.3389/fpls.2019.00818URL [本文引用: 1]
GANGULYD R, CRISPP A, EICHTENS R, POGSONB J. The Arabidopsis DNA methylome is stable under transgenerational drought stress Plant Physiology, 175(2017) 1893-1912. DOI:10.1104/pp.17.00744URL [本文引用: 1]
RACETTEK, ZURWELLERB, TILLMANB, ROWLANDD. Transgenerational stress memory of water deficit in peanut production Field Crops Research, 2020,248:107712. DOI:10.1016/j.fcr.2019.107712URL [本文引用: 1]
DERNERJ D, TISCHLERC R, POLLEYH W, JOHNSONH B. Intergenerational above- and belowground responses of spring wheat (Triticum aestivum L.) to elevated CO2 Basic and Applied Ecology, 2004,5(2):145-152. DOI:10.1078/1439-1791-00217URL [本文引用: 1]
WHITTLEC, OTTOS, JOHNSTONM O, KROCHKOJ. Adaptive epigenetic memory of ancestral temperature regime in Arabidopsis thaliana Botany, 2009,87(6):650-657. DOI:10.1139/B09-030URL [本文引用: 1]
WANGX, XINC, CAIJ, ZHOUQ, DAIT, CAOW, JIANGD. Heat priming induces trans-generational tolerance to high temperature stress in wheat Frontiers in Plant Science, 2016,7:501. [本文引用: 2]
TAOZ, SHENL, GUX, WANGY, YUH, HEY. Embryonic epigenetic reprogramming by a pioneer transcription factor in plants Nature, 2017,551:124-128. DOI:10.1038/nature24300URL [本文引用: 1]
MICHAELSS, AMASINOR. Loss of FLOWERING LOCUS C activity eliminates the late-flowering phenotype of FRIGIDA and autonomous mutations but not responsiveness to vernalization The Plant Cell, 2001,13(4):935-941. DOI:10.1105/tpc.13.4.935URL [本文引用: 2]
GAZZANIS, GENDALLA, LISTERC, DEANC. Analysis of the molecular basis of flowering time variation in Arabidopsis accessions Plant Physiology, 2003,132(2):1107-1114. DOI:10.1104/pp.103.021212URL [本文引用: 2]
MICHAELSS, HEY, SCORTECCIK, AMASINOR. Attenuation of FLOWERING LOCUS C activity as a mechanism for the evolution of summer-annual flowering behavior Arabidopsis Proceedings of the National Academy of Sciences of the United States of America, 2003,100:10102-10107. [本文引用: 2]
YUANW, LUOX, LIZ, YANGW, WANGY, LIUR, DUJ, HEY. A cis cold memory element and a trans epigenome reader mediate Polycomb silencing of FLC by vernalization in Arabidopsis Nature Genetics, 2016,48:1527-1534. DOI:10.1038/ng.3712URL [本文引用: 1]
OLIVERS, FINNEGANE, DENNISE, PEACOCKW, TREVASKISB. Vernalization-induced flowering in cereals is associated with changes in histone methylation at the VERNALIZATION1 gene Proceedings of the National Academy of Sciences of the United States of America, 2009,106(20):8386-8391. [本文引用: 1]
NOSALEWICZA, SIECI?SKAJ, ?MIECHM, NOSALEWICZM, WI?CEKD, PECIOA, WACHD. Transgenerational effects of temporal drought stress on spring barley morphology and functioning Environmental and Experimental Botany, 2016,131:120-127. [本文引用: 1]
WANGX, ZHANGX, CHENJ, WANGX, CAIJ, ZHOUQ, DAIT, CAOW, JIANGD. Parental drought-priming enhances tolerance to post-anthesis drought in offspring of wheat Frontiers in Plant Science, 2018,9:261. DOI:10.3389/fpls.2018.00261URL [本文引用: 2]
ZHANGC, WANGN, ZHANGY, FENGQ, YANGC, LIUB. DNA methylation involved in proline accumulation in response to osmotic stress in rice (Oryza sativa) Genetics and Molecular Research, 2013,12(2):1269-1277. DOI:10.4238/2013.April.17.5URL [本文引用: 1]
ZHANGX, WANGX, ZHONGJ, ZHOUQ, WANGX, CAIJ, LIX, CAOW, JIANGD. Drought priming induces thermo-tolerance to post-anthesis high-temperature in offspring of winter wheat Environmental and Experimental Botany, 2016,127:26-36. DOI:10.1016/j.envexpbot.2016.03.004URL [本文引用: 1]
TABASSUMT, FAROOQM, AHMADR, ZOHAIBA, WAHIDA. Seed priming and transgenerational drought memory improves tolerance against salt stress in bread wheat Plant Physiology and Biochemistry, 2017,118:362-369. DOI:10.1016/j.plaphy.2017.07.007URL [本文引用: 1]
LIY, LIX, YUJ, LIUF. Effect of the transgenerational exposure to elevated CO2 on the drought response of winter wheat: Stomatal control and water use efficiency Environmental and Experimental Botany, 2017,136:78-84. DOI:10.1016/j.envexpbot.2017.01.006URL [本文引用: 1]
LIX, BRESTICM, TAND X, ZIVCAKM, ZHUX, LIUS, SONGF, REITERR J, LIUF. Melatonin alleviates low PS I-limited carbon assimilation under elevated CO2 and enhances the cold tolerance of offspring in chlorophyll b-deficient mutant wheat Journal of Pineal Research, 2018,64(1):e12453. DOI:10.1111/jpi.2018.64.issue-1URL [本文引用: 1]
LIX, JIANGH, LIUF, CAIJ, DAIT, CAOW, JIANGD. Induction of chilling tolerance in wheat during germination by pre-soaking seed with nitric oxide and gibberellin Plant Growth Regulation, 2013,71(1):31-40. DOI:10.1007/s10725-013-9805-8URL [本文引用: 1]
XIEJ J, WANGX, CAIJ, ZHOUQ, DAIT B, JIANGD. Effect of exogenous application of abscisic acid and jasmonic acid at seedling stage on post-anthesis drought stress and physiological mechanisms in wheat Journal of Triticeae Crops, 2018,38(2):221-229. (in Chinese) [本文引用: 1]
WANGX, ZHANGJ, SONGJ, HUANGM, CAIJ, ZHOUQ, DAIT, JIANGD. Abscisic acid and hydrogen peroxide are involved in drought priming‐induced drought tolerance in wheat (Triticum aestivum L.) Plant Biology, 2020,22(6). doi: 10.1111/plb.13143. [本文引用: 1]
LVZ, DIAOM, LIW, CAIJ, ZHOUQ, WANGX, DAIT, CAOW, JIANGD. Impacts of lateral spacing on the spatial variations in water use and grain yield of spring wheat plants within different rows in the drip irrigation system Agricultural Water Management, 2019,212:252-261. DOI:10.1016/j.agwat.2018.09.015URL [本文引用: 1]
 ,, 蔡剑, 周琴, 戴廷波, 姜东
,, 蔡剑, 周琴, 戴廷波, 姜东

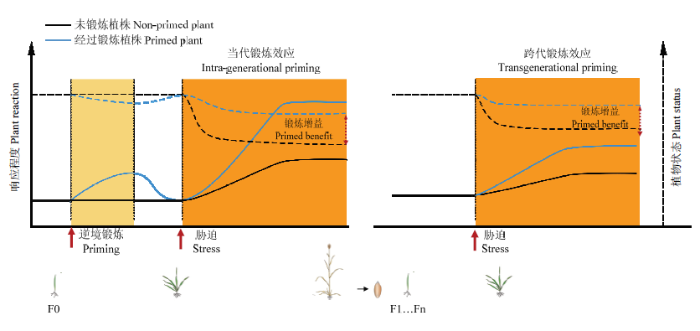 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}