Transient Expression and the Effect on Proliferation and Apoptosis of Granule Cell Stimulating Factor in Ovarian Fibroblasts
LI RunTing,1,2, CHEN LongXin2, ZHANG LiMeng1,2, HE HaiYing1, WANG Yong1, YANG RuoChen1, DUAN ChunHui1, LIU YueQin1, WANG YuQin3, ZHANG YingJie,11College of Animal Science and Technology, Hebei Agricultural University, Baoding 071001, Hebei 2Molecular Biology Laboratory, Zhengzhou Normal University, Zhengzhou 450044 3College of Animal Science and Technology, Henan University of Science and Technology, Luoyang 471023, Henan
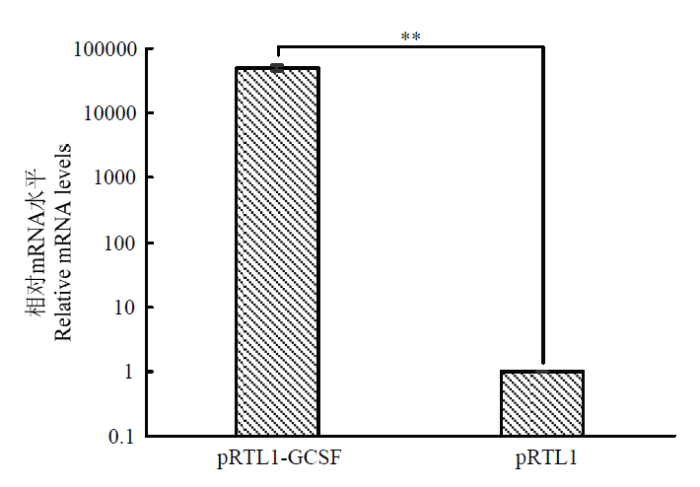
Abstract 【Objective】 The purpose of this paper is to study the transient expression of granule cell stimulating factor (GCSF) in ovarian fibroblast cells, and the influence of GCSF on proliferation, cell cycle, and apoptosis, to provide theoretical basis for molecular genetic breeding of sheep pluripotent stem cells induced by GCSF in the future. 【Method】 The sheep GCSF eukaryotic expression plasmid pRTL1-GCSF and the control vector plasmid pRTL1 were transfected into 1×105 cells·mL-1 sheep fibroblasts respectively. After 48 h of culture, the total RNA was extracted by Trizol method and reverse transcribed into cDNA. The transient expression level of sheep GCSF in fibroblasts was detected by real-time quantitative PCR. GCSF dependent cell line NFS-60 was used for the biological activity of GCSF secreted and expressed in the supernatant of sheep fibroblasts 48 hours after transfection, which was determined by cell viability detection reagent alamarBlue. The HEK 293F suspension culture was used to express the secreted GCSF protein. The GCSF protein expressed in the cell culture medium was purified by Ni-NTA resin and detected by SDS-PAGE. After adding the 30 ng·mL-1 purified GCSF protein, the proliferation of sheep fibroblasts was detected by alamarBlue at 24 h and 48 h, and the cell cycle and apoptosis of sheep fibroblasts were detected by flow cytometry. 【Result】 The expression level of GCSF in sheep fibroblasts was significantly increased after transfection for 48 h. In sheep fibroblasts, the expression level of GCSF transfected with pRTL1-GCSF plasmid was 50 615.92 ± 4 738.83 of that of pRTL1 empty control group. The fluorescence intensity of NFS-60 in the experimental group and positive control group was significantly higher than that in the negative control group and blank control group (P<0.01), but there was no significant difference between the experimental group and the positive control group (P>0.05). The results showed that sheep GCSF could significantly stimulate the proliferation of NFS-60 cells, indicating that the GCSF expressed in sheep fibroblasts had biological activity. After eukaryotic expression of secretory GCSF protein in HEK 293F cell line, the sheep GCSF protein was purified. After 30 ng·mL-1 sheep GCSF was added to sheep fibroblasts, the cell viability of GCSF test group was not significantly different from that of culture medium dilution control group for 24 h and 48 h, but the distribution of cell cycle was significantly changed. At 24 h, compared with the control group, the proportion of G1 phase cells increased from (55.29±1.68)% to (69.37±0.24)%, the difference was very significant (P<0.01); the proportion of S phase cells changed from (15.99±0.38)% to (15.39±0.60)%, the difference was not significant (P>0.05); G2/M phase cells increased significantly (P<0.05), and the proportion increased from (22.88±1.00)% to (26.76±0.82)%. The results showed that 24 hours after the addition of sheep GCSF, the number of cells in division and interphase increased significantly. At 48 h, compared with the control group, the proportion of G1 phase cells decreased from (65.96±0.37)% to (45.69±0.26)%, the difference was very significant (P<0.01); the proportion of S phase cells increased from (13.45±1.33)% to (37.87±2.43)%, the difference was very significant (P<0.01); the proportion of G2/M phase cells changed from (16.42±1.29)% to (21.80±1.86)%, the difference was not significant (P>0.05). The results showed that the number of cells in interphase was significantly decreased and the number of cells in DNA replication state increased significantly at 48 h after adding GCSF. Compared with the control group, the apoptosis rates of the control group (Ctr) and the experimental group (GCSF) were (7.51±0.38)% and (9.16±0.46)% respectively at 24 h culture. At 48 h, the apoptosis rates of the control group and the experimental group were (5.73±0.29)% and (5.39±0.27)%, respectively. At 72 h, the apoptosis rates of control group (Ctr) and experimental group (GCSF) were (8.88±0.45)% and (5.41±0.27)%, respectively. There was a significant difference between 24 h and 72 h (P<0.01), but there was no significant difference at 48 h (P>0.05). The results showed that GCSF promoted the apoptosis within 24 hours, and the apoptosis was inhibited with the prolongation of time. 【Conclusion】 In conclusion, sheep fibroblasts can express GCSF instantaneously and have biological activity. GCSF did not affect the proliferation of sheep fibroblasts, but could regulate its cell cycle and affect cell apoptosis. The results laid a foundation for breeding sheep with high immunity and disease resistance by GCSF mediated by sheep fibroblasts. Keywords:GCSF;sheep;transformation;bio-activity;cell cycles;apoptosis
PDF (1756KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 李闰婷, 陈龙欣, 张丽萌, 何海迎, 王泳, 杨若晨, 段春辉, 刘月琴, 王玉琴, 张英杰. 粒细胞集落刺激因子在羊成纤维细胞中的表达及对细胞增殖和凋亡的影响[J]. 中国农业科学, 2021, 54(11): 2434-2444 doi:10.3864/j.issn.0578-1752.2021.11.015 LI RunTing, CHEN LongXin, ZHANG LiMeng, HE HaiYing, WANG Yong, YANG RuoChen, DUAN ChunHui, LIU YueQin, WANG YuQin, ZHANG YingJie. Transient Expression and the Effect on Proliferation and Apoptosis of Granule Cell Stimulating Factor in Ovarian Fibroblasts[J]. Scientia Acricultura Sinica, 2021, 54(11): 2434-2444 doi:10.3864/j.issn.0578-1752.2021.11.015
采用GraphPad Prism 6.0软件统计分析。细胞试验数据以平均值±SEM表示,t检验分析使用Multiple t tests-one per row程序,P<0.01表示差异极显著,P<0.05表示差异显著,P≥0.05表示差异不显著。定量结果根据2-ΔΔCt法[15]进行处理,用内参基因GAPDH对羊的不同组织中GCSF的表达水平进行均一化处理。
**代表差异极显著(P<0.01),*代表差异显著(P<0.05),ns代表差异不显著(P>0.05) Fig. 5Cell cycles analysis of fibroblast cells with GCSF adding
** means extremely significant difference between the treatments (P<0.01), * means significant difference between treatments (P<0.05), ns means no significant difference between treatments (P>0.05)
BEGLEYC G, LOPEZA F, NICOLAN A, WARREND J, VADASM A, SANDERSONC J, METCALFD. Purified colony-stimulating factors enhance the survival of human neutrophils and eosinophils in vitro: a rapid and sensitive microassay for colony-stimulating factors Blood, 1986,68(1):162-166. DOI:10.1182/blood.V68.1.162.162URL [本文引用: 2]
CLARKS C, KAMENR. The human hematopoietic colony- stimulating factors Science, 1987,236(4806):1229-1237. DOI:10.1126/science.3296190URL [本文引用: 2]
AVALOSB R, GASSONJ C, HEDVATC, QUANS G, BALDWING C, WEISBARTR H, WILLIAMSR E, GOLDED W, DIPERSIOJ F. Human granulocyte colony-stimulating factor: biologic activities and receptor characterization on hematopoietic cells and small cell lung cancer cell lines Blood, 1990,75(4):851-857. DOI:10.1182/blood.V75.4.851.851URL [本文引用: 2]
WANGF, LIUC, PANC Y. Progress of induced pluripotent stem cells (iPSCs) of big domestic animals Journal of Agricultural Biotechnology, 2014,22(10):1286-1297. (in Chinese) [本文引用: 1]
TAID P. Research on generation of arbas cashmere goat induced pluripotent stem cells [D]. Huhehaote: Inner Mongolia University, 2016. (in Chinese) [本文引用: 1]
LIH H, ZHOUG, FUX B, QUZ L, SUNT Z, GUS F. Culture and identification of human skin fibroblasts Chinese Critical Care Medicine, 2005(2):89-91+63. (in Chinese) [本文引用: 1]
CIBELLIJ B, STICES L, GOLUEKEP J, KANEJ J, JERRYJ, BLACKWELLC, PONCEDE LEON F A, ROBLJ M. Cloned transgenic calves produced from nonquiescent fetal fibroblasts Science, 1998,280(5367):1256-1258. DOI:10.1126/science.280.5367.1256URL [本文引用: 1]
HUANGL L, SHIG Q, BAIJ. In vitro culture of sheep embryonic fibroblasts Heilongjiang Journal of Animal Reproduction, 2006,14(2):4-5. (in Chinese) [本文引用: 2]
DOB H, KANGH J, SONGJ A, NGUYENM T, PARKS, YOOJ, NGUYENA N, KWONG G, JANGJ, JANGM, LEES, SOS, SIMS, LEEK J, OSBORNM J, CHOEH. Granulocyte colony-stimulating factor (GCSF) fused with Fc Domain produced from E. coli is less effective than Polyethylene Glycol-conjugated GCSF Scientific Reports, 2017,7(1):1-9. DOI:10.1038/s41598-016-0028-xURL [本文引用: 1]
ICHINOSEY, HARAN, OHTAM, ASOH, CHIKAMAH, KAWASAKIM, KUBOTAI, SHIMIZUT, YAGAWAK. Recombinant granulocyte colony-stimulating factor and lipopolysaccharide maintain the phenotype of and superoxide anion generation by neutrophils Infection and Immunity, 1990,58(6):1647-1652. DOI:10.1128/iai.58.6.1647-1652.1990URL [本文引用: 1]
CULLORJ S, SMITHW, FAIRLEYN, WOODS L, DELLINGERJ D, SOUZAL. Effects of human recombinant granulocyte colony stimulating factor (HR-GCSF) on the hemogram of lactating dairy cattle Veterinary Clinical Pathology, 1990,19(1):9-12. DOI:10.1111/vcp.1990.19.issue-1URL [本文引用: 5]
NIKOLAYN, EVELIINAP, ANTTIK, RAISAG, ANASTASIAS, ANASTASIAS, JOHANNAM, JARIK, JARIK, RASHIDG. Gender specific mechanism of synaptic impairment and its prevention by GCSF in a mouse model of ALS Frontiers in Cellular Neuroscience, 2011,5:26. [本文引用: 2]
MISHRAV V, CHOUDHARYS, SHARMAU, AGGARWALR, AGARWALR, GANDHIK, GORANIYAN. Effects of Granulocyte Colony-Stimulating Factor (GCSF) on persistent thin endometrium in Frozen Embryo Transfer (FET) cycles Journal of Obstetrics and Gynaecology of India, 2016,66(Suppl 1):407-411. DOI:10.1007/s13224-015-0775-9URL [本文引用: 2]
HAYSP A M A, KNUDSENN, NORMANT, KODERA, KRAYNOVV, HOL, CANNINGP C. Modified bovine g-csf polypeptides and their uses. China CN102159230A, 2011-08-17. (in Chinese) [本文引用: 2]
ZHANGY, AXIAK-BECHTELS, FRIEDMANCOWAN C, AMORIMJ, TSURUTAK, DECLUEA E. Evaluation of immunomodulatory effect of recombinant human granulocyte-macrophage colony-stimulating factor on polymorphonuclear cell from dogs with cancer in vitro Veterinary and Comparative Oncology, 2017,15(3):968-979. DOI:10.1111/vco.2017.15.issue-3URL [本文引用: 3]
ZHANGW, XUS, ZHANGZ F. Involvement of macrophage colony-stimulating factor (M-CSF) in the function of follicular granulosa cells and its molecular mechanism Journal of Medical Research, 2016,45(1):100-105. (in Chinese) [本文引用: 2]
WANGX Y, SHIT P, ZHAOZ D, HUW P, SHANGM Y, ZHANGL. The analysis of PI3K-AKT signal pathway based on the proteomic results of sheep embryonic skeletal muscle Scientia Agricultura Sinica, 2020,53(14):2956-2963. (in Chinese) [本文引用: 1]
LIT, MAA T, ZHANGY J, LIUY Q, DUANC H. Effects of silencing INHα gene by RNAi on cycle, apoptosis-related genes in sheep granulosa cells Chinese Journal of Animal and Veterinary Sciences, 2017,48(8):1551-1556. (in Chinese) [本文引用: 1]
LIANGS D, MAL Q, GAOZ Y, ZHUANGY Y, ZHAOY Z. Granulocyte colony-stimulating factor improves neurological function and angiogenesis in intracerebral hemorrhage rats European Review for Medical and Pharmacological Sciences, 2018,22(7):2005-2014. [本文引用: 2]
LIR T, CHENL X, ZHANGL M, HEH Y, WANGY, YANGR C, DUANC H, LIUY Q, WANGY Q, ZHANGY J. Prokaryotic expression and purification of sheep granulocyte colony stimulating factor and its effect on granulosa cell Chinese Journal of Biotechnology, 2020,36(9):1817-1827. doi.org/10.13345/j.cjb.190584. (in Chinese) [本文引用: 3]
BAIY, ANND K, SHENW C. Recombinant granulocyte colony- stimulating factor-transferrin fusion protein as an oral myelopoietic agent Proceedings of the National Academy of Sciences of the United States of America, 2005,102(20):7292-7296. [本文引用: 1]
CUPPJ S, MILLERM A, MONTGOMERYK D, NIELSENT O, O'ConnellJ X, HUNTSMAND, vande R M, GILKSC B, WESTR B. Translocation and expression of CSF1 in pigmented villonodular synovitis, tenosynovial giant cell tumor, rheumatoid arthritis and other reactive synovitides The American Journal of Surgical Pathology, 2007,31(6):970-976. DOI:10.1097/PAS.0b013e31802b86f8URL [本文引用: 1]
ZHAIS Z, GUOH D, LIS Q, ZHAOX S, WANGY, XUL P, LIUK Y, HUANGX J, CHANGY J. Effects of granulocyte colony- stimulating factor on proliferation and apoptosis of B cells in bone marrow of healthy donors Transplantation Proceedings, 2020,52(1):345-352. [本文引用: 1]
MICKIENEG, DALGEDIENEI, DAPKUNASZ, ZVIRBLISG, PESLIAKASH, KAUPINISA, VALIUSM, MISTINIENEE, PLECKAITYTEM. Construction, purification, and characterization of a Homodimeric Granulocyte colony-stimulating factor Molecular Biotechnology, 2017,59(9/10):374-384. DOI:10.1007/s12033-017-0026-7URL [本文引用: 1]
YANJ J, RYUJ H, PIAOH, HWANGJ H, HAND, LEES K, JANGJ Y, LEEJ, KOOT Y, YANGJ. Granulocyte colony-stimulating factor attenuates renal ischemia-reperfusion injury by inducing myeloid- derived suppressor cells Journal of the American Society of Nephrology, 2020,31(4):731-746. DOI:10.1681/ASN.2019060601URL [本文引用: 1]
BROCKMEIERS L, LOVINGC L, EBERLEK C, HAUS J, MOUK T, KEHRLIM E JR. Administration of granulocyte-colony stimulating factor (G-CSF) to pigs results in a longer mean survival time after exposure to Streptococcus suis Veterinary Microbiology, 231:116-119. DOI:10.1016/j.vetmic.2019.03.010URL [本文引用: 1]
ARMENISEA, TREROTOLIP, CIRONEF, DENITTO A, DESARIO C, BERTAZZOLOW, PRATELLIA, DECARON. Use of recombinant canine granulocyte-colony stimulating factor to increase leukocyte count in dogs naturally infected by canine parvovirus Veterinary Microbiology, 2019,231:177-182. DOI:10.1016/j.vetmic.2019.03.015URL [本文引用: 1]
FRASERJ K, GUERRAJ J, NGUYENC Y, INDESJ E, GASSONJ C, NIMERS D. Characterization of a cell-type-restricted negative regulatory activity of the human granulocyte-macrophage colony- stimulating factor gene Molecular and Cellular Biology, 1994,14(3):2213-2221. [本文引用: 1]
PYKHTINAM B, ROMANOVV P, MIROSHNICHENKOS M, BEKLEMISHEVA B. Construction of a Pichia pastoris strain efficiently producing recombinant human granulocyte-colony stimulating factor (rhG-CSF) and study of its biological activity on bone marrow cells Molecular Biology Reports, 2020,47(1):607-620. DOI:10.1007/s11033-019-05169-9URL [本文引用: 1]
FUY. Gene synthesis, prokaryotic expression and activity of human granulocyte colony stimulating factor [D]. Changchun: Jilin University, 2011. (in Chinese) [本文引用: 1]
WUY F, GUM H, YANGS H, WANGT F. Lower platelet count with increased density of platelet antigens in granulocyte colony- stimulating factor mobilized peripheral blood stem cell donors Journal of the Formosan Medical Association, 2020,119(1 Pt 2):204-210. DOI:10.1016/j.jfma.2019.04.003URL [本文引用: 1]
STEPHENSJ M, BENSINKM, BOWERSC, HOLLENBEAKC S. Risks and consequences of travel burden on prophylactic granulocyte colony-stimulating factor administration and incidence of febrile neutropenia in an aged medicare population Current Medical Research and Opinion, 2019,35(2):229-240. DOI:10.1080/03007995.2018.1465906URL [本文引用: 1]
MODIJ, MENZIE-SUDERAMJ, XUH, TRUJILLOP, MEDLEYK, MARSHALLM L, TAOR, PRENTICEH, WUJ Y. Mode of action of granulocyte-colony stimulating factor (G-CSF) as a novel therapy for stroke in a mouse model Journal of Biomedical Science, 2020,27(1):19. DOI:10.1186/s12929-019-0597-7URL [本文引用: 1]
PUTZE J, EDERJ M, REINHARDTT A, SACCOR E, CASASE, LIPPOLISJ D. Differential phenotype of immune cells in blood and milk following pegylated granulocyte colony-stimulating factor therapy during a chronic Staphylococcus aureus infection in lactating Holsteins Journal of Dairy Science, 2019,102(10):9268-9284. DOI:10.3168/jds.2019-16448URL [本文引用: 1]
DENIZY K, BURCUA, AKKIZ? Y, ?zdemirH H, SIVI?? Z, HüSEYINO, FERDA?. Congenital neutropenia patient with hypomorphic biallelic CSF3R mutation responding to GCSF Journal of Pediatric Hematology/Oncology, 2019,41(3):190-192. [本文引用: 1]
GUANX L, HOUL C, YANGH, HUJ, WANGC, WANGX P, BIS, XUX Y, WANGK X, HEM. Effects of G-CSF on expression of bcl-2 and caspase-3 around hematoma on acute intracerebral hemorrhage (ICH) in rats Journal of Apoplexy and Nervous Diseases, 2015,32(6):511-513. (in Chinese) [本文引用: 1]
YAOY L, LIY X, REHEMAN·A,ZHANG J, MENGC H, WANGH L, QIANY, CAOS X. Cloning of Sp1 Gene CDS region of Hu sheep and its effect on proliferation and apoptosis of granulosa cell Chinese Journal of Animal and Veterinary Sciences, 2017,48(11):2098-2106. (in Chinese) [本文引用: 1]
 ,1,2, 陈龙欣2, 张丽萌1,2, 何海迎1, 王泳1, 杨若晨1, 段春辉1, 刘月琴1, 王玉琴3, 张英杰
,1,2, 陈龙欣2, 张丽萌1,2, 何海迎1, 王泳1, 杨若晨1, 段春辉1, 刘月琴1, 王玉琴3, 张英杰

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT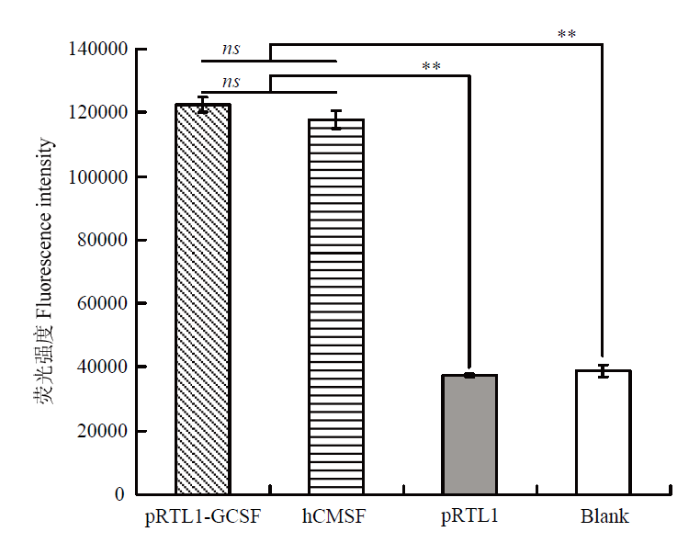 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT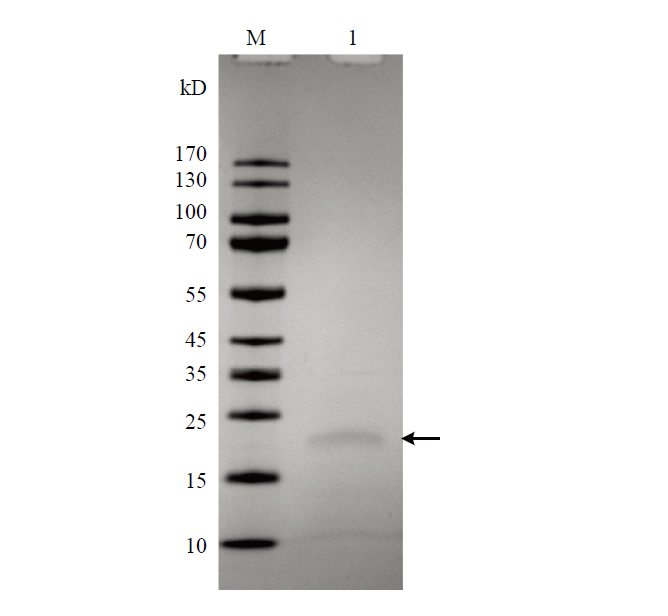 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT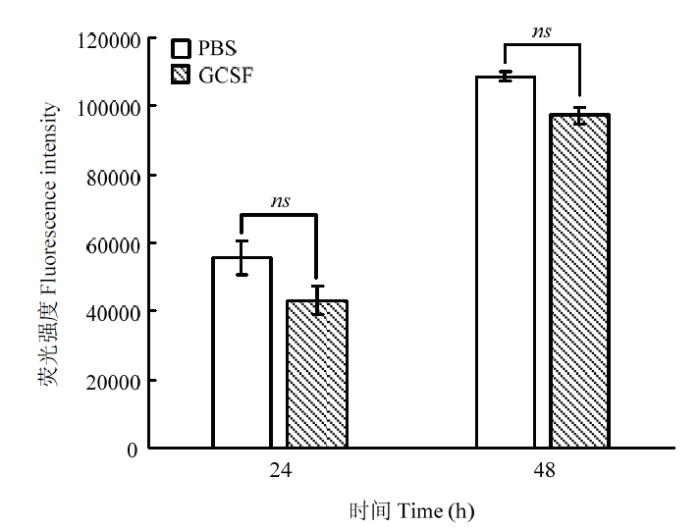 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT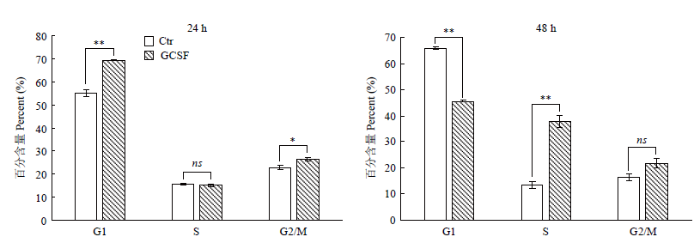 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}