Research Progress on Mechanisms of Apoptosis to Postmortem Tenderization in Muscle
HUANG Feng,, WEI QiChao, LI Xia, LIU ChunMei, ZHANG ChunHui,Institute of Food Science and Technology, Chinese Academy of Agricultural Sciences/Key Laboratory of Agro-Products Processing, Ministry of Agriculture, Beijing 100193
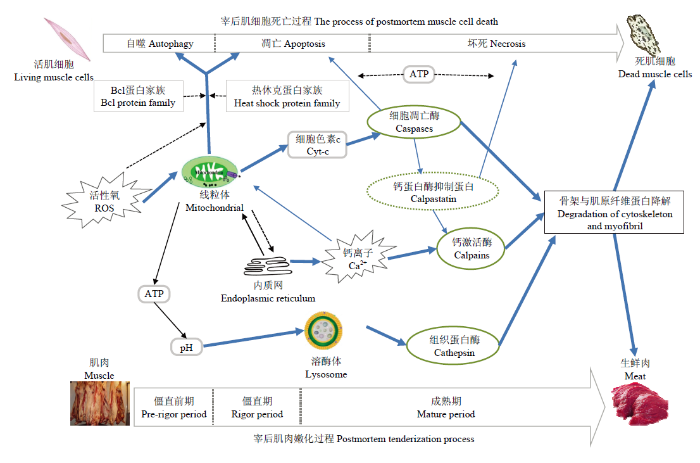
Abstract Tenderness has been considered as one of the most important eating quality characteristics of meat, while inconsistent changes of tenderness in postmortem (PM) muscles can significantly reduce the purchasing intention. Therefore, investigating mechanism of postmortem muscle tenderization is becoming more and more important in the past decade. Since apoptosis definition was highlighted in the PM tenderization process, it has been widely concerned. Shortly after slaughter, the reactive oxygen species (ROS) was significantly accumulated, and ATP (adenosine triphosphate) was gradually exhausted within muscle fibers, which inevitably led to skeletal cell death. Both PM cell death and meat tenderization refer to the activation of muscle endogenous enzymes by a series of regulatory factors, followed by the degradation of muscle structural proteins. The two biochemical processes are considered to be highly related. PM muscle cells die mainly through apoptosis. Besides, in the early stage of PM, when a small amount of ROS is produced, the cells start their defense system by autophagy and ATP therefore gradually deplete muscle cells in the later stage, which may change from apoptosis to necrosis. It has been documented that mitochondrial pathway is crucial for the apoptosis activation in PM muscles. The release of apoptotic factors from mitochondria is the master node of the intracellular death cascade reaction. The opening status of mitochondrial outer membrane directly determines the way, in which the muscle fiber dies. In this paper, the release mechanism of apoptotic factors induced by PM mitochondrial damage was discussed from the perspectives of mitochondrial membrane permeability and cristae remodeling, and the regulation of mitochondrial damage on PM muscle tenderization was discussed. Moreover the underlying mechanism behind it was also analyzed to reveal the effect of mitochondria on muscle pH regulation through energy metabolism, the release of apoptotic factors and regulation of apoptosis enzyme activity. At the same time, the interaction between mitochondria and endoplasmic reticulum were discussed, focusing on Ca2+ signal transduction and cell apoptosis process. The interaction between mitochondria and lysosome was further investigated, by highlighting the stability of lysosomal membrane and the subsequently released cathepsin to activate Bax and Bid to accelerate mitochondrial membrane permeability. Caspases were activated and involved in the limited degradation of some myofibrils in the early stage of PM tenderization, followed by the inactivation resulting from the decreasing ATP or other factors with the extended PM time. Therefore, caspases maybe only involve the early stage of tenderization. This review could provide a theoretical reference for the perfection of PM muscle tenderization investigations. Keywords:postmortem tenderization;apoptosis;caspases;mitochondrial damage;apoptotic factors
PDF (1940KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 黄峰, 魏起超, 李侠, 刘春梅, 张春晖. 细胞凋亡对宰后肌肉嫩化作用机理的研究进展[J]. 中国农业科学, 2021, 54(10): 2192-2202 doi:10.3864/j.issn.0578-1752.2021.10.014 HUANG Feng, WEI QiChao, LI Xia, LIU ChunMei, ZHANG ChunHui. Research Progress on Mechanisms of Apoptosis to Postmortem Tenderization in Muscle[J]. Scientia Acricultura Sinica, 2021, 54(10): 2192-2202 doi:10.3864/j.issn.0578-1752.2021.10.014
HOLMAN B WB, VAN DE VEN RJ, MAO YW, COOMBS C EO, HOPKINS DL. Using instrumental (CIE and reflectance) measures to predict consumers' acceptance of beef colour Meat Science, 2017,127:57-62. DOI:10.1016/j.meatsci.2017.01.005URL [本文引用: 1]
SHACKELFORD SD, WHEELER TL, MEADE MK, REAGAN JO, BYRNES BL, KOOHMARAIEM. Consumer impressions of Tender Select beef Journal of Animal Science, 2001,79(10):2605-2614. DOI:10.2527/2001.79102605xURL [本文引用: 1]
KOOHMARAIEM, KENT MP, SHACKELFORD SD, VEISETHE, WHEELER TL. Meat tenderness and muscle growth: Is there any relationship? Meat Science, 2002,62(3):345-352. DOI:10.1016/S0309-1740(02)00127-4URL [本文引用: 1]
KOOHMARAIEM, GEESINK GH. Contribution of postmortem muscle biochemistry to the delivery of consistent meat quality with particular focus on the calpain system Meat Science, 2006,74(1):34-43. DOI:10.1016/j.meatsci.2006.04.025URL [本文引用: 2]
GEESINK GH, KUCHAYS, CHISHTI AH, KOOHMARAIEM. μ-calpain is essential for postmortem proteolysis of muscle proteins Journal of Animal Science, 2006,84(10):2834-2840. DOI:10.2527/jas.2006-122URL [本文引用: 1]
OUALIA, HERRERA-MENDEZ CH, COULISG, BECILAS, BOUDJELLALA, AUBRYL, SENTANDREU MA. Revisiting the conversion of muscle into meat and the underlying mechanisms Meat Science, 2006,74(1):44-58. DOI:10.1016/j.meatsci.2006.05.010URL [本文引用: 3]
SENTANDREU MA, COULISG, OUALIA. Role of muscle endopeptidases and their inhibitors in meat tenderness Trends in Food Science & Technology, 2002,13(12):400-421. [本文引用: 1]
NIKOLETOPOULOUV, MARKAKIM, PALIKARASK, TAVERNARAKISN. Crosstalk between apoptosis, necrosis and autophagy Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 2013,1833(12):3448-3459. DOI:10.1016/j.bbamcr.2013.06.001URL [本文引用: 1]
HUANGM, HUANGF, HUANG JC, XU BC, ZHOU GH, XU XL. Advances in research on postmortem tenderization mechanism of endogenous proteolytic enzymes in muscle Scientia Agricultura Sinica, 2011,44(15):3214-3222. (in Chinese) [本文引用: 1]
OUALIA, HERRERA-MENDEZ CH, COULISG, SAMIRAB, BOUDJELLALA, HARHOURAK, AUBRYL, SENTANDREU MA. Meat tenderisation and muscle cell death, two highly related events Tehnologija Mesa, 2007,48(1/2):1-15. [本文引用: 1]
HERRERA-MENDEZ CH, BECILAS, BOUDJELLALA, OUALIA. Meat ageing: Reconsideration of the current concept Trends in Food Science & Technology, 2006,17(8):394-405. [本文引用: 1]
CAO JX, SUN WQ, ZHOU GH, XU XL, PENG ZQ, HU ZL. Morphological and biochemical assessment of apoptosis in different skeletal muscles of bulls during conditioning Journal of Animal Science, 2010,88(10):3439-3444. DOI:10.2527/jas.2009-2412URL [本文引用: 2]
BECILAS, HERRERA-MENDEZ CH, COULISG, LABASR, ASTRUCT, PICARDB, BOUDJELLALA, PELISSIERP, BREMAUDL, OUALIA. Postmortem muscle cells die through apoptosis European Food Research and Technology, 2010,231(3):485-493. DOI:10.1007/s00217-010-1296-5URL [本文引用: 1]
HERRERA-MENDEZ CH, BECILAS, COULISG, SENTANDREU MA, AUBRYL, OUALIA. Purification and partial characterization of antithrombin III from bovine skeletal muscle and possible role of thrombin in postmortem apoptosis development and in efficiency of low voltage electrical stimulation Food Research International, 2010,43(1):356-363. DOI:10.1016/j.foodres.2009.10.012URL [本文引用: 1]
KEMP CM, PARRT, BARDSLEY RG, BUTTERY PJ. Comparison of the relative expression of caspase isoforms in different porcine skeletal muscles Meat Science, 2006,73(3):426-431. DOI:10.1016/j.meatsci.2005.12.009URL [本文引用: 2]
CHENL, FENG XC, LUF, XU XL, ZHOU GH, Li QY, GUO XY. Effects of camptothecin, etoposide and Ca2+ on caspase-3 activity and myofibrillar disruption of chicken during postmortem ageing Meat Science, 2011,87(3):165-174. DOI:10.1016/j.meatsci.2010.10.002URL [本文引用: 2]
UNDERWOOD KR, MEANS WJ, DUM. Caspase 3 is not likely involved in the postmortem tenderization of beef muscle Journal of Animal Science, 2008,86(4):960-966. DOI:10.2527/jas.2007-0549URL [本文引用: 1]
ADHIHETTY PJ, HOOD DA. Mechanisms of apoptosis in skeletal muscle Basic & Applied Myology, 2003,13(4):171-179. [本文引用: 1]
WANGL, LIU YH, WEIL. New theories and concepts of the mode of cell death Medicine and Philosophy (Clinical Decision Making Forum Edition), 2011,32(14):61-63. (in Chinese) [本文引用: 2]
LANAA, ZOLLAL. Apoptosis or autophagy, that is the question: Two ways for muscle sacrifice towards meat Trends in Food Science & Technology, 2015,46(2):231-241. [本文引用: 2]
JIAX. The research of the effect of autophagy on the apoptosis of mutton [D]. Yangling: Northwest A&F University, 2017. (in Chinese) [本文引用: 1]
DEGTEREVA, HUANG ZH, BOYCEM, LI YQ, JAGTAPP, MIZUSHIMAN, CUNY GD, MITCHISON TJ, MOSKOWITZ MA, YUAN JY. Chemical inhibitor of nonapoptotic cell death with therapeutic potential for ischemic brain injury Nature Chemical Biology, 2005,1(2):112-119. DOI:10.1038/nchembio711URL [本文引用: 1]
HENCKELP, KARLSSONA, JENSEN MT, OKSBJERGN, PETERSEN JS. Metabolic conditions in porcine longissimus muscle immediately pre-slaughter and its influence on peri- and post mortem energy metabolism Meat Science, 2002,62(2):145-155. DOI:10.1016/S0309-1740(01)00239-XURL [本文引用: 1]
BOCK FJ, TAIT S WG. Mitochondria as multifaceted regulators of cell death Nature Reviews Molecular Cell Biology, 2020,21(2):85-100. DOI:10.1038/s41580-019-0173-8URL [本文引用: 1]
HUANGF, HUANGM, ZHANGH, GUOB, ZHANG DQ, ZHOU GH. Cleavage of the calpain inhibitor, calpastatin, during postmortem ageing of beef skeletal muscle Food Chemistry, 2014,148:1-6. DOI:10.1016/j.foodchem.2013.10.016URL [本文引用: 1]
SHI YX, MELNIKOV VY, SCHRIER RW, EDELSTEIN CL. Downregulation of the calpain inhibitor protein calpastatin by caspases during renal ischemia-reperfusion American Journal of Physiology-Renal Physiology, 2000,279(3):F509-F517. DOI:10.1152/ajprenal.2000.279.3.F509URL [本文引用: 1]
P?RN-ARES MI, SAMALIA, ORRENIUSS. Cleavage of the calpain inhibitor, calpastatin, during apoptosis Cell Death & Differentiation, 1998,5(12):1028-1033. [本文引用: 1]
CHUA BT, GUOK, LIP. Direct cleavage by the calcium-activated protease calpain can lead to inactivation of caspases Journal of Biological Chemistry, 2000,275(7):5131-5135. DOI:10.1074/jbc.275.7.5131URL [本文引用: 1]
GUPTAS, GOLLAPUDIS. Susceptibility of na?ve and subsets of memory T cells to apoptosis via multiple signaling pathways Autoimmunity Reviews, 2007,6(7):476-481. DOI:10.1016/j.autrev.2007.02.005URL [本文引用: 1]
NAKANISHIK, SUDOT, MORISHIMAN. Endoplasmic reticulum stress signaling transmitted by ATF6 mediates apoptosis during muscle development Journal of Cell Biology, 2005,169(4):555-560. [本文引用: 1]
BRUNELLE JK, CHANDEL NS. Oxygen deprivation induced cell death: An update Apoptosis, 2002,7(6):475-482. DOI:10.1023/A:1020668923852URL [本文引用: 1]
CAO JX, OU CR, ZOU YF, YE KP, ZHANG QQ, KHAN MA, PAN DD, ZHOUG. Activation of caspase-3 and its correlation with shear force in bovine skeletal muscles during postmortem conditioning Journal of Animal Science, 2013,91(9):4547-4552. DOI:10.2527/jas.2013-6469URL [本文引用: 2]
HUANGF, HUANGM, ZHANGH, ZHANG CJ, ZHANG DQ, ZHOU GH. Changes in apoptotic factors and caspase activation pathways during the postmortem aging of beef muscle Food Chemistry, 2016,190:110-114. DOI:10.1016/j.foodchem.2015.05.056URL [本文引用: 1]
WANG LL, HANL, MA XL, YU QL, ZHAO SN. Effect of mitochondrial apoptotic activation through the mitochondrial membrane permeability transition pore on yak meat tenderness during postmortem aging Food Chemistry, 2017,234:323-331. DOI:10.1016/j.foodchem.2017.04.185URL [本文引用: 4]
R?NNING SB, ANDERSEN PV, PEDERSEN ME, HOLLUNGK. Primary bovine skeletal muscle cells enters apoptosis rapidly via the intrinsic pathway when available oxygen is removed Plos ONE, 2017,12(8):e0182928. DOI:10.1371/journal.pone.0182928URL [本文引用: 1]
JIANGX. Mechanism of release of mitochondrial apoptotic proteins [D]. Beijing: Peking Union Medical College, 2014. (in Chinese) [本文引用: 1]
QUINTANA-CABRERAR, MEHROTRAA, RIGONIG, SORIANO ME. Who and how in the regulation of mitochondrial cristae shape and function Biochemical and Biophysical Research Communications, 2018,500(1):94-101. DOI:10.1016/j.bbrc.2017.04.088URL [本文引用: 2]
GOGVADZEV, ORRENIUSS, ZHIVOTOVSKYB. Multiple pathways of cytochrome c release from mitochondria in apoptosis Biochimica et Biophysica Acta (BBA)-Bioenergetics, 2006,1757(5):639-647. DOI:10.1016/j.bbabio.2006.03.016URL [本文引用: 1]
HUANG GM, WANG YP, SUNC, ZHANG XT, ZHANGH. Advances in the study of mitochondrial ultrastructure and its regulatory mechanism Progress in Biochemistry and Biophysics, 2019,46(12):1141-1149. (in Chinese) [本文引用: 1]
CHENL, DONGJ, YAN CJ, SONG ZY. The role and regulation of mitochondrial cristae remodeling Progress in Physiological Sciences, 2018,49(1):3-13. (in Chinese) [本文引用: 1]
ZHENGK, YANG MG, YAN CJ, TANG ML, SONG ZY. Mitochondrial dynamics and apoptosis Chinese Journal of Cell Biology, 2019,41(8):1467-1476. (in Chinese) [本文引用: 1]
MALHEIROS JM, BRAGA CP, GROVE RA, RIBEIRO FA, CALKINS CR, ADAMECJ, CHARDULO L AL. Influence of oxidative damage to proteins on meat tenderness using a proteomics approach Meat Science, 2019,148:64-71. DOI:10.1016/j.meatsci.2018.08.016URL [本文引用: 1]
MATARNEH SK, ENGLAND EM, SCHEFFLER TL, YEN CN, WICKS JC, SHIH, GERRARD DE. A mitochondrial protein increases glycolytic flux Meat Science, 2017,133:119-125. DOI:10.1016/j.meatsci.2017.06.007URL [本文引用: 1]
MATARNEH SK, BELINEM, DE LUZ E SILVAS, SHIH, GERRARD DE. Mitochondrial F1-ATPase extends glycolysis and pH decline in an in vitro model Meat Science, 2018,137:85-91. DOI:10.1016/j.meatsci.2017.11.009URL [本文引用: 1]
SCHEFFLER TL, MATARNEH SK, ENGLAND EM, GERRARD DE. Mitochondria influence postmortem metabolism and pH in an in vitro model Meat Science, 2015,110:118-125. DOI:10.1016/j.meatsci.2015.07.007URL [本文引用: 1]
LIU ZY, DU XN, DENGJ, GU MY, HU HL, GUIM, YIN CC, CHANG ZZ. The interactions between mitochondria and sarcoplasmic reticulum and the proteome characterization of mitochondrion- associated membrane from rabbit skeletal muscle Proteomics, 2015,15(15):2701-2704. DOI:10.1002/pmic.v15.15URL [本文引用: 1]
HUANGF, DING ZJ, ZHANG CJ, HU HH, ZHANGL, ZHANGH. Effects of calcium and zinc ions injection on caspase-3 activation and tenderness in post-mortem beef skeletal muscles International Journal of Food Science & Technology, 2018,53(3):582-589. [本文引用: 2]
XUEL, YIN CC. Progress in the study of mitochondria-endoplasmic reticulum physical coupling Chinese Journal of Cell Biology, 2013,35(12):1791-1796. (in Chinese) [本文引用: 1]
CHERUBINIM, LOPEZ-MOLINAL, GINESS. Mitochondrial fission in Huntington's disease mouse striatum disrupts ER-mitochondria contacts leading to disturbances in Ca2+ efflux and Reactive Oxygen Species (ROS) homeostasis Neurobiology of Disease, 2020,136:104741. DOI:10.1016/j.nbd.2020.104741URL [本文引用: 1]
O'HALLORAN GR, TROY DJ, BUCKLEY DJ, REVILLE WJ. The role of endogenous proteases in the tenderisation of fast glycolysing muscle Meat Science, 1997,47(3/4):187-210. DOI:10.1016/S0309-1740(97)00046-6URL [本文引用: 1]
ZHANG JY, LI MQ, YU QL, HANL, MA ZL. Effects of lysosomal-mitochondrial apoptotic pathway on tenderness in post-mortem bovine longissimus muscle Journal of Agricultural and Food Chemistry, 2019,67(16):4578-4587. DOI:10.1021/acs.jafc.9b00894URL [本文引用: 1]
KEMP CM, BARDSLEY RG, PARRT. Changes in caspase activity during the postmortem conditioning period and its relationship to shear force in porcine longissimus muscle Journal of Animal Science, 2006,84(10):2841-2846. DOI:10.2527/jas.2006-163URL [本文引用: 1]
ZHANG MH, WANG DY, HUANGW, LIUF, ZHU YZ, XU WM, CAO JX. Apoptosis during postmortem conditioning and its relationship to duck meat quality Food Chemistry, 2013,138(1):96-100. DOI:10.1016/j.foodchem.2012.10.142URL [本文引用: 1]
BERNARDC, CASSAR-MALEKI, LE CUNFFM, DUBROEUCQH, RENANDG, HOCQUETTE JF. New indicators of beef sensory quality revealed by expression of specific genes Journal of Agricultural and Food Chemistry, 2007,55(13):5229-5237. DOI:10.1021/jf063372lURL [本文引用: 1]
LAVILLEE, SAYDT, MORZELM, BLIENTS, CHAMBONC, LEPETITJ, RENANDG, HOCQUETTE JF. Proteome changes during meat aging in tough and tender beef suggest the importance of apoptosis and protein solubility for beef aging and tenderization Journal of Agricultural and Food Chemistry, 2009,57(22):10755-10764. DOI:10.1021/jf901949rURL [本文引用: 1]
CHENL, FENG XC, ZHANG YY, LIU XB, ZHANG WG, LI CB, ULLAHN, Xu XL, ZHOU GH. Effects of ultrasonic processing on caspase-3, calpain expression and myofibrillar structure of chicken during post-mortem ageing Food Chemistry, 2015,177:280-287. DOI:10.1016/j.foodchem.2014.11.064URL [本文引用: 1]
HUANGM, HUANGF, XU XL, ZHOU GH. Influence of caspase3 selective inhibitor on proteolysis of chicken skeletal muscle proteins during post mortem aging Food Chemistry, 2009,115(1):181-186. DOI:10.1016/j.foodchem.2008.11.095URL [本文引用: 1]
HUANGM, HUANGF, MA HJ, XU XL, ZHOU GH. Preliminary study on the effect of caspase-6 and calpain inhibitors on postmortem proteolysis of myofibrillar proteins in chicken breast muscle Meat Science, 2012,90(3):536-542. DOI:10.1016/j.meatsci.2011.09.004URL [本文引用: 1]
KEMP CM, PARRT. The effect of recombinant caspase 3 on myofibrillar proteins in porcine skeletal muscle Animal, 2008,2(8):1254-1264. DOI:10.1017/S1751731108002310URL [本文引用: 1]
HUANGM, HUANGF, XUEM, XU XL, ZHOU GH. The effect of active caspase-3 on degradation of chicken myofibrillar proteins and structure of myofibrils Food Chemistry, 2011,128(1):22-27. DOI:10.1016/j.foodchem.2011.02.062URL [本文引用: 1]
MOHRHAUSER DA, UNDERWOOD KR, WEAVER AD. In vitro degradation of bovine myofibrils is caused by μ-calpain, not caspase-3 Journal of Animal Science, 2011,89(3):798-808. DOI:10.2527/jas.2010-3149URL [本文引用: 1]
HUANGF, HUANGM, ZHOU GH, XU XL, XUEM. In vitro proteolysis of myofibrillar proteins from beef skeletal muscle by caspase-3 and caspase-6 Journal of Agricultural and Food Chemistry, 2011,59(17):9658-9663. DOI:10.1021/jf202129rURL [本文引用: 1]
 ,, 魏起超, 李侠, 刘春梅, 张春晖
,, 魏起超, 李侠, 刘春梅, 张春晖

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT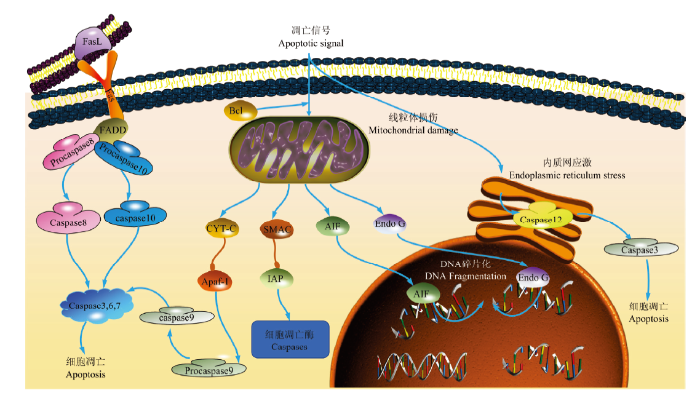 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}