Using CRISPR/Cas9-mediated Targeted Mutagenesis of ZmFKF1 Delayed Flowering Time in Maize
YANG Min,1, XU HuaWei2, WANG CuiLing2, YANG Hu1, WEI YueRong,11Institute of Fruit Tree Research, Guangdong Academy of Agricultural Sciences/ Key Laboratory of South Subtropical Fruit Biology and Genetic Resource Utilization (MOA)/Guangdong Province Key Laboratory of Tropical and Subtropical Fruit Tree Research, Guangzhou 510640 2College of Agriculture, Henan University of Science and Technology, Luoyang 471000, Henan
Abstract 【Objective】FKF1 is a key gene that plays an important role in many plant flowering pathways. In order to study the function of FKF1 in maize, ZmFKF1 editing mutants were obtained by using CRISPR/cas9 technology. In this study, we used these mutants as materials to clarify the role of ZmFKF1 in maize flowering pathway through phenotypic analysis and the expression analysis of key flowering genes. The result provide a theoretical basis for molecular breeding and genetic improvement of maize.【Method】 ZmFKF1 gene was cloned from B104, and its structure was determined by sequence alignment. The targeted sequences of ZmFKF1 were designed according to the principle of CRISPR/Cas9 technology, then these targeted sequences were compared and analyzed in maize reference genome, and the non-specific target site was excluded. Finally, ZmFKF1-T1 on exon 1 of ZmFKF1 was selected to construct CRISPR/cas9 gene editing expression vector. At the same time, B104 immature embryos were selected as explants to transform by Agrobacterium-mediated genetic transformation, the resistant calli were obtained through resistance screening, and then buds and roots were induced. The T0 generation ZmFKF1 gene editing positive plants were obtained and verified by using the specific primers of cas9. The target site amplification and sequencing analysis was used to determine whether the T1 transgenic lines had mutation at the expected target site of ZmFKF1 and the type of mutation, and screened the homozygous lines of ZmFKF1 site mutation. After obtaining the above materials, the flowering phenotypes of these materials were statistically analyzed with wild type as control. At the same time, qRT-PCR was used to detect the expression of key genes that related to flowering pathway in the above materials and further verify the phenotype. 【Result】The target sequence was designed on exon1 of ZmFKF1 to construct gene editing expression vector. The transgenic lines were obtained by genetic transformation realized site directed mutagenesis of ZmFKF1. A total of 18 T0 generation ZmFKF1 gene-edited lines were obtained, and 6 of them were generated mutation on the exon1 with two different mutations types, including single base insertion and multiple bases deletion. The phenotype analysis shown that the flowering time of three T2 generation ZmFKF1 homozygous lines was delayed compared with wild type B104. Furthermore, the expression levels of three related flowering pathway genes (ZmGI, conz1 and ZmZCN8) in the mutants were significantly lower than those in the wild type B104 (P<0.05), which was consistent with the late flowering phenotype of the mutants. 【Conclusion】CRISPR/Cas9 technology can be used to edit ZmFKF1 to obtain gene editing mutants, and the flowering time of these mutants is significantly delayed compared to wild type. Keywords:CRISPR/Cas9;gene editing;maize;ZmFKF1;flowering time
PDF (4395KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 杨敏, 胥华伟, 王翠玲, 杨护, 魏岳荣. 利用CRISPR/Cas9技术研究玉米ZmFKF1在开花过程中的作用[J]. 中国农业科学, 2021, 54(4): 696-707 doi:10.3864/j.issn.0578-1752.2021.04.003 YANG Min, XU HuaWei, WANG CuiLing, YANG Hu, WEI YueRong. Using CRISPR/Cas9-mediated Targeted Mutagenesis of ZmFKF1 Delayed Flowering Time in Maize[J]. Scientia Acricultura Sinica, 2021, 54(4): 696-707 doi:10.3864/j.issn.0578-1752.2021.04.003
以玉米B104幼胚为受体材料,利用农杆菌介导的遗传转化法进行遗传转化。试验材料采用常规种植与田间管理。sgRNA中间载体pYLgRNA-U6b、CRISPR/Cas9双元载体由华南农业大学刘耀光院士惠赠,试验所用引物及测序由生工生物工程(上海)股份有限公司完成。BsaⅠ购置于NEB公司,T4 DNA ligase和PrimeSTAR GXL DNA Polymerase购置于宝生物工程(大连)有限公司、KOD FX购置于东洋纺(上海)生物科技有限公司,其余分子试剂主要购置于天根生化科技(北京)有限公司和生工生物工程(上海)股份有限公司。大肠杆菌感受态细胞DH5α购自天根生化科技(北京)有限公司;农杆菌感受态细胞EHA105购自北京华越洋生物有限公司。
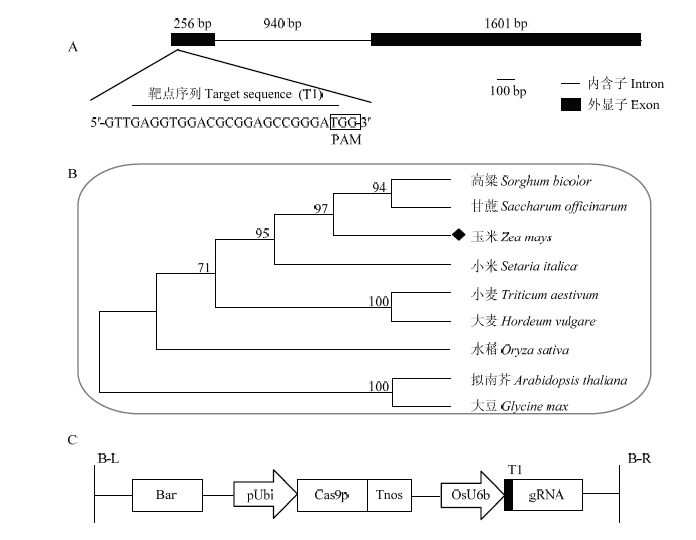
A:靶位点在ZmFKF1中的位置;B:9个物种的FKF1分子进化树分析;C:pYLCRISPR/Cas9-ZmFKF1-T1质粒T-DNA区示意图 Fig. 1Target site of the gRNA in the ZmFKF1 gene, phylogenetic tree analysis of FKF1 amino acid sequences in nine species and the diagram of T-DNA region of pYLCRISPR/Cas9-ZmFKF1-T1 plasmid
A: Position of the target in ZmFKF1 gene locus; B: Phylogenetic tree analysis of FKF1 amino acid sequences in 9 species; C: The diagram of T-DNA region of pYLCRISPR/Cas9-ZmFKF1-T1 plasmid
Table 2 表2 表2ZmFKF1编辑突变体突变类型分析 Table 2Mutation analysis on the exon1 of ZmFKF1 gene
株系 Line
序列 Sequence (5′-3′)
突变类型 Mutation type
是否纯合 Homozygous or not
B104
AGGTGGACGCGGAGCCGGGATGG
---
---
C1-6
AGGTGGACGCGGAGCTATGG
+1(T),-4(CGGG)
否 No
C2-12
AGGTGGACGCGGAGCCGAGGATGG
+1(A)
是 Yes
C3-3
AGGTGGACGCGGAGCCG
-9(GGATGGCCG)
否 No
C5-7
AGGTGGACGCGGAGCCGGGATGG
-20(AGGTGGACGCGGAGCCGGGA)
否 No
C6-2
AGGTGGACGCGGAGCCATGG
-3(GGG)
是 Yes
C7-1
AGGTGGACGCGGAGCCG
-6(GGGATG)
是 Yes
加粗字体表示PAM序列,下划线处表示插入的碱基,+表示碱基插入,-表示碱基缺失,---表示野生型,无变化 The bold font represented PAM sequence. The underlined base indicated the insertion base. + represented base insertion, - indicated base deletion, --- represented wild type (B104)
A:ZmFKF1蛋白结构域预测结果;B:ZmFKF1编辑突变体的氨基酸序列预测分析及比对结果 Fig. 4Prediction of ZmFKF1 protein domain and analysis of ZmFKF1 coding amino acid sequence in mutants
A: Prediction results of ZmFKF1 protein domain; B: Amino acid sequence prediction analysis and comparison results of ZmFKF1 in ZmFKF1gene editing mutants
A:C7-1和B104在播种后67 d的表型;B:C7-1和B104在播种后70 d的表型;C:不同基因型材料中开花关键基因的表达情况,其中ZmUbi为内参基因,利用2-ΔΔCT方法计算基因相对表达量结果 Fig. 5Phenotype and flowering related genes expression analysis of ZmFKF1 editing lines
A: The phenotype of C7-1 compared to ‘B104’ at 67 days after sowing; B: The phenotype of C7-1 compared to ‘B104’ at 70 days after sowing; C: Expression analysis of flowering related genes in different genotype lines. ZmUbi primers were used for PCR as an internal control, the result of relative genes expression was calculated by 2-ΔΔCT method
Table 3 表3 表3不同基因型植株玉米抽雄时间、吐丝时间和散粉时间的统计 Table 3Statistical results of flowering time in different genotype lines
基因型 Genotype
抽雄时间 Male phase
散粉时间 Pollen phase
吐丝时间 Female phase
B104
62.70±1.37
67.87±1.04
70.53±1.25
C2-12
63.43±1.04*
69.13±1.36*
72.57±1.65*
C6-2
64.07±0.98*
69.37±1.56*
71.90±1.22*
C7-1
63.96±1.30*
69.04±1.81*
72.33±1.12*
采用随机分区设计种植3个ZmFKF1编辑纯合突变体和野生型B104对照,3个重复,每个小区中各基因型株系种5行,每行6株。随机选取各基因型株系0株进行表型统计分析,取平均值,并进行方差和P值分析。*表示在0.05水平差异显著 Three ZmFKF1gene editing homozygous mutants and wild type B104 were planted in a random area design, with 3 replicates. There were 5 lines of each genotype in each plot, with 6 plants in each row. 10 strains of each genotype were randomly selected for phenotypic statistical analysis. The average value was taken, and the variance and P value were analyzed. * indicate significant difference at 0.05 level
KU LX, SUN ZH, WANG CL, ZHANGJ, ZHANG WQ, CHEN YH. QTL analysis of the photoperiod sensitivity-related traits at different developmental stages in maize (Zea mays L.) Acta Agronomica Sinica, 2010,36(4):602-611. (in Chinese) [本文引用: 1]
OILM, KHARSHIING EV. Photoreceptor mediated plant growth responses: Implications for photoreceptor engineering toward improved performance in crops Frontiers in Plant Science, 2017,8:1181-1194. [本文引用: 1]
NELSON DC, LASSWELLJ, ROGG LE, COHEN MA, BARTELB. FKF1, a clock-controlled gene that regulates the transition to flowering in Arabidopsis Cell, 2000,101(3):331-340. [本文引用: 5]
SAWAM, NUSINOW DA, KAY SA, IMAIZUMIT. FKF1 and GIGANTEA complex formation is required for day-length measurement in Arabidopsis Science, 2007,318(5848):261-265. [本文引用: 3]
IMAIZUMIT. FKF1 F-Box protein mediates cyclic degradation of a repressor of CONSTANS in Arabidopsis Science, 2005,309(5732):293-297. [本文引用: 3]
GORALOGIA GS, LIUT, ZHAOL, PANIPINTO PM, IMAIZUMIT. CYCLING DOF FACTOR 1 represses transcription through the TOPLESS co-repressor to control photoperiodic flowering in Arabidopsis The Plant Journal, 2017,92(2):244-262. [本文引用: 2]
KOBAYASHIY, WEIGELD. Move on up, it’s time for change- mobile signals controlling photoperiod-dependent flowering Genes & Development, 2007,21(19):2371-2384. [本文引用: 2]
HWANG DY, PARKS, LEES, IMAIZUMIT, SONG YH. GIGANTEA regulates the timing stabilization of CONSTANS by altering the interaction between FKF1 and ZEITLUPE Molecular Cells, 2019,42(10):693-701. [本文引用: 2]
CORBESIERL, VINCENTC, JANGS, FORNARA, F, FAN QZ, SEARLEI, GIAKOUNTISA, FARRONAS, GISSOTL, TURNBULLC, COUPLANDG. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis Science, 2007,316:1030-1033. [本文引用: 2]
D’ALOIAM, BONHOMMED, BOUCHéF, TAMSEDDAKK, ORMENESES, TORTIS, COUPLANDG, PERILLEUXC. Cytokinin promotes flowering of Arabidopsis via transcriptional activation of the FT paralogue TSF The Plant Journal, 2011,65(6):972-979. [本文引用: 2]
SONG YH, SMITH RW, TO BJ, MILLAR AJ, IMAIZUMIT. FKF1 conveys timing information for CONSTANS stabilization in photoperiodic flowering Science, 2012,336(6084):1045-1049. [本文引用: 2]
HAN SH, YOO SC, LEE BD, ANG. PAEKN C. Rice FLAVIN-BINDING, KELCH REPEAT, F-BOX1 (OsFKF1) promotes flowering independent of photoperiod Plant Cell Environ, 2015,38(12):2527-2540. [本文引用: 3]
MOUTIQR, RIBAUT JM, EDMEADESG, KRAKOWSKY MD, LEEM. Quantitative trait loci for photoperiod response in maize Maize Genetics Conference Abstracts, 2002(43):147. [本文引用: 2]
WANG CL, CHENG FF, SUN ZH, TANG JH, WU LC, KU LX, CHEN YH. Genetic analysis of photoperiod sensitivity by molecular markers in a tropical by temperate maize recombinant in bred population Theoretical and Applied Genetics, 2008,117:1129-1139. [本文引用: 2]
WANG XT, WU LJ, ZHANG SF, WU LC, KU LX, WEI XM, XIE LL, CHEN YH. Robust expression and association of ZmCCA1 with circadian rhythms in maize Plant Cell Report, 2011,30:1261-1272. [本文引用: 2]
CHARDONF, VIRLONB, MOREAUL, FALQUEM, JOETSJ, DECOUSSETL, MURIGNEUXA, CHARCOSSETA. Genetic architecture of flowering time in maize as inferred from QTL meta-analysis and synteny conservation with the rice genome Genetics, 2004,162:2169-2185. [本文引用: 2]
COLES ND, MCMULLEN MD, BALINT-KURTI PJ, PRATT R C, HOLLAND J B. Genetic control of photoperiod sensitivity in maize revealed by joint multiple population analysis Genetics, 2010,184(3):799-812. [本文引用: 2]
DUCROCQS, GIAUFFRETC, MADURD, COMBESV, DUMASF, JOUANNES, COUBRICHED, JAMINP, MOREAUL, CHARCOSSETA. Fine mapping and haplotype structure analysis of a major flowering time quantitative trait locus on maize chromosome 10 Genetics, 2009,183(4):1555-1563. [本文引用: 2]
MU GQ, LIANGY, ZHANG ZM, WU YQ, LIU SJ, PENGH, ZHANG SZ, PAN GT. Mapping quantitative trait loci associated with photoperiod sensitivity in maize (Zea mays L.) Agricultural Sciences in China, 2009,8(1):24-30. [本文引用: 2]
HUNG HY, SHANNON LM, TIANF, BRADBURY PJ, HOLLAND JB. ZmCCT and the genetic basis of day-length adaptation underlying the post domestication spread of maize Proceedings of the National Academy of Sciences of the United States of America, 2012,109(28):1913-1921. [本文引用: 2]
YANGQ, LIZ, LIW. CACTA-like transposable element in ZmCCT attenuated photoperiod sensitivity and accelerated the post domestication spread of maize Proceedings of the National Academy of Sciences of the United States of America, 2013,110:16969-16974. [本文引用: 2]
ZHAO SJ. Gene cloning and expression analysis of pseudo-response regulator 73(ZmPRR73) in maize [D]. Luoyang: Henan University of Science and Technology, 2018. (in Chinese) [本文引用: 2]
LIUL. Molecular evolution and association analysis of ZmFKF1, a clock-controlled gene in maize [D]. Yaan: Sichuan Agricultural University, 2014. (in Chinese) [本文引用: 1]
LI JF, NORVILLE JE, AACHJ, MCCORMACKM, ZHANG DD, BUSHJ, CHURCH GM, SHEENJ. Multiplex and homologous recombination-mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9 Nature Biotechnology, 2013,31(8):688-691. [本文引用: 1]
MAO YF, ZHANGH, XU NF, ZHANG BT, GOUF, ZHU JK. Application of the CRISPR-Cas system for efficient genome engineering in plants Molecular Plant, 2013,6(6):2008-2011. [本文引用: 1]
WANG YP, CHENGX, SHAN QW, ZHANGY, LIU JX, GAOCX, QIU JL. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew Nature Biotechnology, 2014,32(9):947-951. [本文引用: 1]
LIC, UNVERT, ZHANG BH. A high-efficiency CRISPR/Cas9 system for targeted mutagenesis in cotton (Gossypium hirsutum L.) Scientific Reports, 2017,7:43902. [本文引用: 1]
ITOY, NISHIZAWA-YOKOIA, ENDOM, MIKAMIM, TOKIS. CRISPR/Cas9-mediated mutagenesis of the RIN locus that regulates tomato fruit ripening Biochemical & Biophysical Research Communications, 2015,467(1):76-82. [本文引用: 1]
ZHOU XH, JACOBS TB, XUE LJ, HARDING SA, TSAI CJ. Exploiting SNPs for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-coumarate: CoA ligase specificity and redundancy New Phytologist, 2015,208(2):298-301. [本文引用: 1]
MALNOYM, VIOLAR, JUNG MH, KOO OJ, KIMS, KIM JS, VELASCOR, KANCHISWAMY CN. DNA-free genetically edited grapevine and apple protoplast using CRISPR/Cas9 ribonucleoproteins Frontiers in Plant Science, 2016,7:1904. [本文引用: 1]
YANG LS, GUOY, HUY, WEN YQ. CRISPR/Cas9-mediated mutagenesis of VviEDR2 results in enhanced resistance to powdery mildew in grapevine (Vitis vinifera) Acta Horticulturae Sinica, 2020,47(4):623-634. (in Chinese) [本文引用: 1]
ZHENG AH, ZHANGF, JIANGM, YUANQ, JIANG LY, CHENQ, TANG HR, SUNB. Targeted editing of BoaZDS by CRISPR/Ca9 technology in Chinese kale Acta Horticulturae Sinica, 2019,46(1):57-64. (in Chinese) [本文引用: 1]
ZOU XP, FAND, PENG AH, HE YR, XU LZ, LEI TG, YAO LX, LIQ, LUO KM, CHEN SC. CRISPR/Cas9-mediated editing of multiple sites in the citrus CsLOB1 promoter Acta Horticulturae Sinica, 2019,46(2):337-344. (in Chinese) [本文引用: 1]
FENGC, YUANJ, WANGR, LIUY, BIRCHLER JA, HAN FP. Efficient targeted genome modification in maize using CRISPR/Cas9 system Journal of Genetics & Genomics, 2016,43(1):37-43. [本文引用: 3]
ZENG DC, MA XL, XIE XR, ZHU QL, LIU YG. A protocol for CRISPR/Cas9-based multi-gene editing and sequence decoding of mutant sites in plants Scientia Sinica Vitae, 2018,48(7):783-794. (in Chinese) [本文引用: 1]
WANG XT. Cloning and characterization of photoperiod sensitive gene ZmGI in maize Zhengzhou: Henan Agricultural University, 2009. (in Chinese) [本文引用: 1]
MILLER TA, MUSLIN EH, DORWEILER JE. A maize CONSTANS-like gene, conz1, exhibits distinct diurnal expression patterns in varied photoperiods Planta, 2008,227(6):1377-1388. [本文引用: 1]
MENGX, MUSZYNSKI MG, DANILEVSKAYA ON. The FT-like ZCN8 gene functions as a floral activator and is involved in photoperiod sensitivity in maize The Plant Cell, 2011,23(3):942-960. [本文引用: 1]
LEBLANCC, ZHANGF, MENDEZJ, LOZANOY, CHATPARK, IRISH VF, JACOBY. Increased efficiency of targeted mutagenesis by CRISPR/Cas9 in plants using heat stress The Plant Journal, 2018,93(2):377-386. [本文引用: 1]
 ,1, 胥华伟2, 王翠玲2, 杨护1, 魏岳荣
,1, 胥华伟2, 王翠玲2, 杨护1, 魏岳荣

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT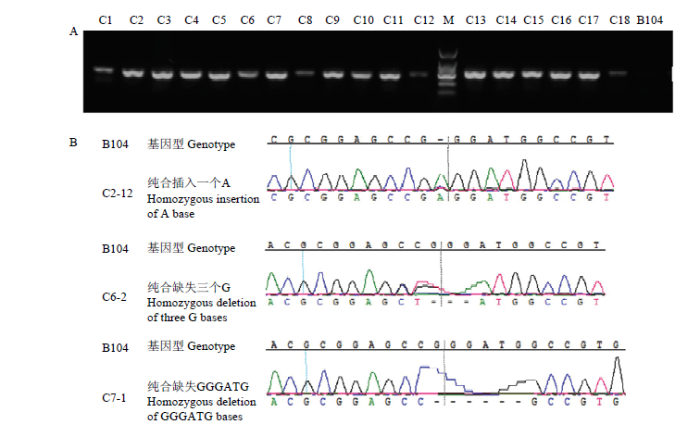 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT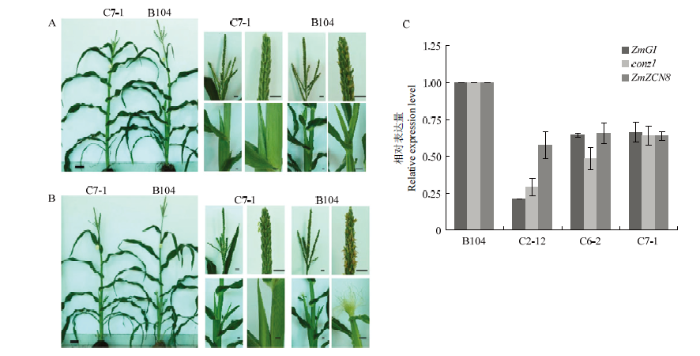 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}