 ,1, 解振强2, 诸葛雅贤1, 白云赫1, 管乐1, 吴伟民3, 张培安1, 郑婷1, 房经贵1, 王晨,1
,1, 解振强2, 诸葛雅贤1, 白云赫1, 管乐1, 吴伟民3, 张培安1, 郑婷1, 房经贵1, 王晨,1Function Analysis of GA3 Mediate miR171s and Its Target Genes VvSCLs in Grape Seed Development
WANG WenRan,1, XIE ZhenQiang2, ZHUGE YaXian1, BAI YunHe1, GUAN Le1, WU WeiMin3, ZHANG PeiAn1, ZHENG Ting1, FANG JingGui1, WANG Chen,1通讯作者:
责任编辑: 赵伶俐
收稿日期:2020-04-27接受日期:2020-05-27网络出版日期:2021-01-16
| 基金资助: |
Received:2020-04-27Accepted:2020-05-27Online:2021-01-16
作者简介 About authors
王文然,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (1673KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王文然, 解振强, 诸葛雅贤, 白云赫, 管乐, 吴伟民, 张培安, 郑婷, 房经贵, 王晨. GA3介导miR171s及其靶基因VvSCLs调控葡萄种子发育的作用分析[J]. 中国农业科学, 2021, 54(2): 357-369 doi:10.3864/j.issn.0578-1752.2021.02.011
WANG WenRan, XIE ZhenQiang, ZHUGE YaXian, BAI YunHe, GUAN Le, WU WeiMin, ZHANG PeiAn, ZHENG Ting, FANG JingGui, WANG Chen.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】葡萄(Vitis vinifera L.)是世界四大水果之一,其在鲜食、加工方面均具有极重要的经济价值。无核是鲜食和制干葡萄重要的优良性状,无核因其食用方便,风味口感俱佳,倍受消费者青睐,目前已成为重要的育种目标,也是能够显著提高葡萄商品价值的首要性状。葡萄种子由种皮、胚乳和胚构成,胚是种子的重要组成结构。细胞学水平的研究发现,胚败育是导致葡萄无核化的重要原因之一[1],胚的各部分都是由胚性细胞组成,因此胚性细胞能否正常发育是影响胚发育及无核的重要因素。【前人研究进展】目前在萝卜[2]、百合(Lilium pumilum DC. Fisch)[3]、龙眼(Dimocarpus longan)[4]、落叶松(Larix kaempferi)[5]及柑橘(Citrus reticulata)[1]中均证实miR171介导SCL6转录因子调控植物胚性细胞发育;同时发现杏(Prunus sibirica L.)PsmiR171-SCL是调控杏核发育的重要miRNA[6]。这些研究均表明,miR171介导SCL转录因子调控胚性细胞发育,而miR171家族是功能高度保守的家族[6],因此可推断vvi-miR171s有可能调控VvSCLs影响葡萄胚性细胞发育,进而影响种子发育。2002年在拟南芥中分离得到AtmiR171,并确定AtSCL6-II、AtSCL6-III和AtSCL6-IV为其靶基因[3]。SCL6是GRAS家族SCL亚家族的成员,是植物中特有的转录因子,其功能具有多样性,可参与调控体细胞胚发生[7]、根组织发育[8]、重金属胁迫[9]、光周期的调节[10]、节间形成[11]、叶绿素合成[12]、叶片形成[13]、干旱胁迫[14]及花芽分化[15]等多种生长发育进程。目前,miRBase数据库中已收录了38种植物的miR171家族(http://www.mirbase.org/),但不同物种间家族成员数量不同,大豆的miR171家族成员数量多达21个[16],而在拟南芥[15]中仅发现3个,在酿酒葡萄‘黑比诺’中鉴定出8个成员(http:// www.mirbase.org/),说明miR171家族成员既具有保守性,也具有一定的特异性。据报道,GA可调控miR171-SCL进而影响植物生命活动,拟南芥miR171-SCL调节光照条件下叶绿素合成和叶片生长受GA - DELLA信号途径调控;而龙眼早期体胚发生是基于DlmiR171b对GA3的响应,从而介导DlSCL6表达进而调控细胞形态建成[4]。虽然目前在龙眼[4]和百合[3]等物种的研究中,均已证明某个miR171家族成员可靶向SCL转录因子调控胚性细胞发育。【本研究切入点】尚未阐明miR171家族不同成员是共同还是独立介导靶基因参与胚性细胞发育调控,更不清楚miR171家族能否应答赤霉素介导靶基因参与葡萄种子发育的调控。【拟解决的关键问题】拟通过研究vvi-miR171s应答GA3介导靶基因VvSCLs的作用机制,明确它们在赤霉素诱导葡萄无核过程中的作用,从表观遗传学角度阐释赤霉素介导葡萄无核的分子机制。1 材料与方法
1.1 材料及其处理
以优质的5年生二倍体葡萄‘魏可’为试验材料,于2017年5月2日(花前10 d)用50 mg·L-1 GA3 浸蘸花序30 s,以清水处理为对照,在花后5 d(5 DAF)、10 d(10 DAF)、20 d(20 DAF)、30 d(20 DAF)和45 d(20 DAF)分别采集果实,并将种子(对照组)/种子区(GA3处理后的无核果实中与对照组种子相对应的组织部位)分离后液氮速冻,存于-80℃。1.2 vvi-miR171s成熟体序列克隆及前体鉴定
在miRBase数据库(http://www.mirbase.org/)搜索并下载葡萄miR171家族(vvi-miR171s)的成熟体序列。根据miRBase中下载的vvi-miR171s成熟体序列,设计特异引物,利用miR-RACE技术[17]在‘魏可’葡萄果实组织中克隆及测序鉴定其精确序列。1.3 vvi-miR171s靶基因预测
运用PSRNA Target(http://plantgrn.noble.org/v1_ psRNATarget/),对已鉴定的vvi-miR171s成熟体序列进行靶基因预测。预测时选择User-submitted small RNAs/preloaded transcripts一栏,并将最大期望(maximum expectation)设置为3.0,其他参数为默认设置。1.4 vvi-miR171s及其靶基因的生物信息学分析
分别使用MEGA5.0软件的邻接法(Neighbor-Joining)、Gene Structure Display Server(GSDS)(http://gsds.cbi.pku.edu.cn/index.php)、SMART网站(http://smart.embl-heidelberg.de/)及MEME网站(http://meme.nbcr.net/meme/)进行系统进化树、基因序列结构、保守结构域及模序分析。启动子序列在CRIBI数据库(http://genomes.cribi.unipd.it/gb2/ gbrowse/public/vitis_vinifera_v2/)和Grape Genome Browser(http://www.genoscope.cns.fr/cgi-bin/ggb/vitis/ 12X/gbrowse/vitis/)中查询,取VvSCL15/22起始密码子(ATG)上游1 500 bp的序列或vvi-miR171s上游1 500 bp序列作为该基因的预测启动子,并在NEW PLACE网站(https://www.dna.affrc.go.jp/ PLACE/? action=newplace/)进行启动子序列的顺式作用元件分析。1.5 检测靶裂解位点及3′末端裂解频率
用加ploy(A)尾巴的mRNA反转录合成的cDNA作为PPM-RACE【(Poly(A) polymerase-mediated 3′ RACE)poly(A)聚合酶介导3′RACE RNA】反应的模板,通用引物Gene RACE 3′为:5′-ATTCTAGAG GCCGAGGCGGCCGACATG-3′。用加接头的mRNA反转录成的cDNA作为RLM-RACE【(RNA ligase- mediated 5′ RACE)连接酶介导5′ RACE】反应的模板,通用引物Gene RACE 5′为:5′-GGACACTGACATG GACTGAAGGAGTA-3′。详细步骤参考WANG等[18]的方法。1.6 数据处理与分析
使用Excel 2017进行数据整理,SPSS 19.0进行相关性分析。2 结果
2.1 GA3处理后不同发育时期葡萄种子的发育状态
从图1可以看出,GA3使果实纵向拉长。5 DAF对照组和GA3处理组的果实胚珠均正常发育,无明显差异。10 DAF时,GA3处理的果实胚珠与5 DAF的几乎无差别,无明显地生长,而对照组的果实胚珠已经明显大于处理组。20 DAF、30 DAF和45 DAF均观察到对照组的果实胚珠正常发育,在45 DAF已形成坚硬的种子;而GA3处理组在45 DAF果实中种子区仅存一条细线。因此,GA3在诱导葡萄无核过程中强烈地抑制了葡萄种子的发育。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1GA3对果实种子发育的影响
Fig. 1Effect of GA3 on grape berry seed development
2.2 vvi-miR171s成熟体序列克隆及其序列分析
2.2.1 序列克隆及其序列比对 不同于miRBase数据库中鉴定出10个miR171家族成员,仅在‘魏可’葡萄中鉴定出vvi-miR171a/b/e/f/g/h 6个成员(图2-a),且与miRBase数据库中‘黑比诺’品种的同源序列完全一致(图2-b),表明其序列在葡萄属植物中具有高度的保守性。图2
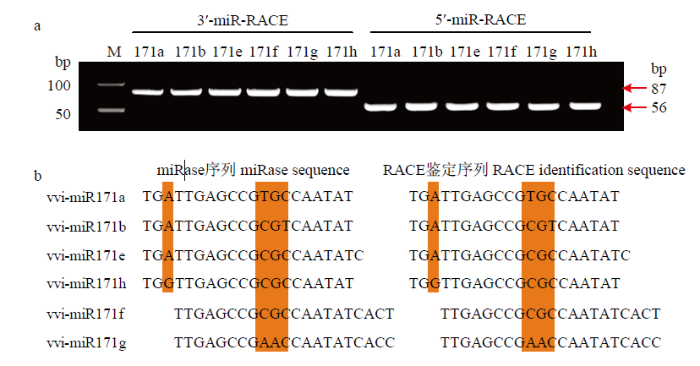 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2vvi-miR171s成熟体片段的PCR产物(a)及Race技术鉴定的vvi-miR171s成熟体序列与miRBase同源基因序列比较(b)
Fig. 2PCR products of vvi-miR171s in grape (a), comparison of cloned vvi-miR171s mature sequences with the homologous sequences in grapes from miRBase (b)
2.2.2 vvi-miR171s成熟体序列进化树分析 为了解miR171s在植物中的进化特性,对比了包括葡萄在内的5个物种39条miR171s的成熟体序列,包括葡萄6条、拟南芥3条、苹果(Monodelphis domestica)14条、柑橘9条、桃(Prunus persica)7条。利用MEGA 5.0构建上述植物miR171s成熟体系统发育树(图3),结果发现这5个物种的miR171s成员进化树被分为3大主枝:其中第二主枝仅有ppe-miR171e一个成员,第三主枝为crt-miR171b-5p和mdm- miR171f-5p,其余成员均在第一主枝。而在第一主枝上的vvi-miR171g和mdm-miR171i与其他成员距离较远,分别位于第二、三分枝,vvi-miR171a、vvi-miR171e、vvi-miR171f、vvi-miR171h和vvi-miR171b集中在第一主枝的第一分枝上,且该分枝的每一小枝均包含5个物种。因此,vvi-miR171a、vvi-miR171e、vvi-miR171f、vvi-miR171h和vvi-miR171b 5个成员亲缘关系较近,与vvi-miR171g的亲缘关系较远。同时发现miR171s在进化速率上有较大差异,而在同一植物中不同成员也呈现出差异性。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3miR171s成熟体的系统发育树(圆点标识为vvi-miR171s)
Fig. 3Phylogeny tree of miR171 family mature sequences (Dot marks denote vvi-miR171s)
2.3 vvi-miR171s的靶基因预测及其序列匹配程度分析
基于已鉴定的vvi-miR171a/b/c/d/e/f序列,利用PSRNA Target为其预测到3条靶基因,分别为VvSCL6、VvSCL15、VvSCL22(表1)。其中,vvi-miR171e与靶基因的错配率最低为0,vvi-miR171a、vvi-miR171b和vvi-miR171h的错配率均为0.5,vvi-miR171f的错配率为1.0,vvi-miR171g的错配率最高为4。这表明VvSCL6、VvSCL5和VvSCL22可能为vvi-miR171s的潜在靶基因,且vvi-miR171s不同成员对靶基因的作用强度可能存在差异。Table 1
表1
表1vvi-miR171s预测靶基因结果
Table 1
| miRNA成员 miRNA_Acc. | miRNA 成熟体序列 miRNA_mature sequence | 靶基因基因编号 Target_Acc. | 期望值 Expectation | 未配对时 的能量UPE | 裂解区域 Target region | 抑制作用 Inhibition | 多重性 Multiplicity |
|---|---|---|---|---|---|---|---|
| vvi-miR171a | TGATTGAGCCGTGC CAATATC | VIT_204s0023g01380.1(VvSCL15) | 0.5 | -1 | CDS:656-676 | 裂解 Cleavage | 1 |
| VIT_215s0048g00270.1 (VvSCL6) | 0.5 | -1 | CDS:1594-1614 | 裂解 Cleavage | 1 | ||
| VIT_202s0154g00400.1 (VvSCL22) | 0.5 | -1 | CDS:1724-1744 | 裂解 Cleavage | 1 | ||
| vvi-miR171b | TGATTGAGCCGCGT CAATATC | VIT_204s0023g01380.1 (VvSCL15) | 0.5 | -1 | CDS:656-676 | 裂解 Cleavage | 1 |
| VIT_215s0048g00270.1 (VvSCL6) | 0.5 | -1 | CDS:1594-1614 | 裂解 Cleavage | 1 | ||
| VIT_202s0154g00400.1 (VvSCL22) | 0.5 | -1 | CDS:1724-1744 | 裂解 Cleavage | 1 | ||
| vvi-miR171e | TGATTGAGCCGCGC CAATATC | VIT_204s0023g01380.1 (VvSCL15) | 0 | -1 | CDS:656-676 | 裂解 Cleavage | 1 |
| VIT_215s0048g00270.1 (VvSCL6) | 0 | -1 | CDS:1594-1614 | 裂解 Cleavage | 1 | ||
| VIT_202s0154g00400.1 (VvSCL22) | 0 | -1 | CDS:1724-1744 | 裂解 Cleavage | 1 | ||
| vvi-miR171f | TTGAGCCGCGCCAA TATCACT | VIT_215s0048g00270.1 (VvSCL6) | 1 | -1 | CDS:1591-1611 | 裂解 Cleavage | 1 |
| VIT_202s0154g00400.1 (VvSCL22) | 1 | -1 | CDS:1721-1741 | 裂解 Cleavage | 1 | ||
| VIT_204s0023g01380.1 (VvSCL15) | 1 | -1 | CDS:653-673 | 裂解 Cleavage | 1 | ||
| vvi-miR171g | TTGAGCCGAACCAA TATCACC | VIT_204s0023g01380.1 (VvSCL15) | 4 | -1 | CDS:653-673 | 翻译 Translation | 1 |
| VIT_215s0048g00270.1 (VvSCL6) | 4 | -1 | CDS:1591-1611 | 翻译 Translation | 1 | ||
| VIT_202s0154g00400.1 (VvSCL22) | 4 | -1 | CDS:1721-1741 | 翻译 Translation | 1 | ||
| vvi-miR171h | TGGTTGAGCCGCGC CAATATC | VIT_204s0023g01380.1 (VvSCL15) | 0.5 | -1 | CDS:656-676 | 裂解 Cleavage | 1 |
| VIT_215s0048g00270.1 (VvSCL6) | 0.5 | -1 | CDS:1594-1614 | 裂解 Cleavage | 1 | ||
| VIT_202s0154g00400.1 (VvSCL22) | 0.5 | -1 | CDS:1724-1744 | 裂解 Cleavage | 1 |
新窗口打开|下载CSV
2.4 vvi-miR171s的靶基因验证及其靶裂解位点分析
植物中miRNAs主要通过裂解靶基因进而调控一系列生长发育进程,因此鉴定miRNAs对靶基因的裂解作用是验证miRNAs靶基因的最有效途径。利用5′-RLM-RACE检测到vvi-miR171s对VvSCL6、VvSCL15和VvSClL22的裂解产物,同时发现VvSCL6有2个裂解位点,分别位于第9位和第12位,裂解频率分别为7/20、10/20;VvSCL15有3个裂解位点,分别位于第9位、第12位和第13位,裂解频率分别为13/25、11/25和1/25;VvSCL22有2个裂解位点,分别位于第9位和12位,裂解频率分别为3/20和20/23。这3条靶基因的裂解位点均具有位于miRNA的5′端第9位和第10位碱基之间所匹配的靶点,这些结果不仅证实了这3条基因是miR171s的真实靶基因,而且也表明它们裂解作用位点的保守性。2.5 vvi-miR171s靶基因的染色体分布及基因结构分析
为了进一步了解SCL家族的基因结构及功能区域,对其进行了染色体定位(图5-a)、构建进化树(图5-b)、基因结构(图5-c)、motif(图5-d)、domain分析(图5-e),发现6个miR171s成员及3个靶基因分别定位在9条不同的染色体上。为了研究SCL家族成员间的进化关系,利用邻接法构建了VvSCL6/15/22及其他物种中同源基因蛋白序列的进化树(图5-b)。进化树显示,5个物种的15个SCLs基因共分为两个主枝,其中所有SCL6和SCL22分布在第一分枝上,而所有SCL15聚集在第二主枝上,这说明在进化过程中SCL6和SCL22较为相似;VvSCL6/22与桃、樱桃(Prunus avium)和苹果中的同源基因亲缘关系较近,而VvSCL15则与拟南芥、桃、樱桃和苹果中的同源基因进化距离相同,这说明SCL家族的不同成员在进化过程中具有特异性。仅VvSCL22中包含1个内含子区,其他成员的基因序列中只包含UTR区和外显子区。同时发现VvSCL6/22与其亲缘关系较近的PpSCL6/22、PaSCL6/22和MdSCL6/22的外显子区域长度较为一致,明显长于亲缘关系较远的拟南芥同源基因;而5个SCL15的外显子区长度也基本相同。结合motif和domain分析发现,这15个SCL家族成员motif类型及排列顺序较为保守,且均具有GRAS结构域,因此可推测该家族功能较为保守,并在motif分析中找到15个SCL蛋白中该结构域对应的保守基序,分别为motif 5/6/8/9/10(图5-f)。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4vvi-miR171s对靶基因的裂解位点示意图
Fig. 4The diagram of cleavage site between vvi-miR171s and its targets
图5
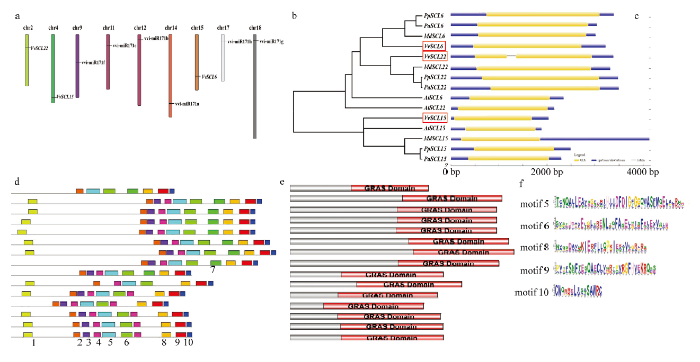 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5vvi-miR171s及VvSCLs染色体定位;SCLs的基因结构、motif、及domain分析
a:vvi-miR171s及VvSCLs在葡萄染色体的定位;b:邻接法构建5个物种SCLs蛋白的系统发育树;c:与进化树基因序列对应的SCLs基因的结构分析;d:SCLs蛋白保守基序分析;e:保守结构域分析;f:SCLs蛋白保守结构域具有的共同保守基序
Fig. 5The distribution of vvi-miR171s and VvSCLs on grapevine chromosomes, and analysis of SCLs, motif and domain
a: The distribution of vvi-miR171s and VvSCLs on grapevine chromosomes; b: The unrooted tree of SCLs protein of 5 species was generated by neighbor-joining method; c: The exon-intron composition of SCL genes; d: The conserved motifs of SCL proteins; e: Conserved domains; f: SCLs protein has common conserved motifs of the conserved domain
2.6 vvi-miR171s启动子的顺式作用元件分析
在vvi-miR171s及其靶基因的启动子中,富含5类顺式作用元件分别是激素、光信号、低温和干旱胁迫、组织特异性和昼夜周期节律相关作用元件(图6-a)。对比6个miR171s成员的作用元件发现,数量上,vvi-miR171h最多,而vvi-miR171g的元件数量最少;类型上,除vvi-miR171a/g含有昼夜节律元件外,其余成员的启动子均仅含有其余4类作用元件,其中激素、逆境和组织特异性相关响应元件占总数量的比重较大,而光响应元件数量较少。与vvi-miR171s类似,靶基因中仅VvSCL6启动子中发现1个昼夜节律相关元件,光响应元件数量最少,其他3种类型的元件数量较多。同时发现miR171s及靶基因启动子均含有大量与胚乳和花粉发育有关的顺式作用元件,这些调控元件说明vvi-miR171s及其靶基因不仅对光、低温和干旱胁迫等外界环境信号做出响应,而且还能应答激素,可能参与葡萄无核果实的生长发育。图6
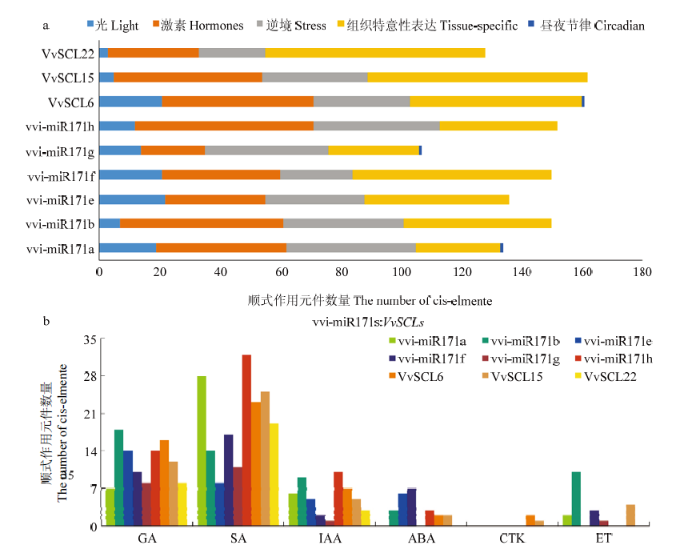 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6vvi-miR171s及靶基因启动子顺式作用元件分析
a:vvi-miR171s启动子中各类响应元件的数量;b:响应不同激素的作用元件数量
Fig. 6Motif analysis of the promoters from the vvi-miR171s precursor and their target VvSCL genes
a: The total number of diverse types of motifs derived from vvi-miR171s and VvSCLs promoters; b: The proportion of each type of hormones related elements of vvi-miR171s and its targeted genes VvSCLs, respectively
为进一步认识vvi-miR171s及其靶基因在激素信号途径中的作用,对激素相关顺式作用元件进行分类(图6-b)。所有启动子均含有激素相关响应元件,主要包括GA、SA、生长素(Auxin,IAA)、脱落酸(Abscisic acid,ABA)、细胞分裂素(Cytokinin,CTK)及乙烯(Ethylene,ET)等激素响应元件,表明它们可应答多种激素信号进而参与调控葡萄生长发育。其中,GA、SA和IAA相关元件在9个基因启动子中均存在,而ABA响应元件在vvi-miR171a/g和VvSCL22中未发现,CTK相关顺式作用元件仅在VvSCl6/15中发现,ET元件则在vvi-miR171e/h和VvSCL6/22中未发现。另外,在众多激素元件中,GA和SA作用元件的数量明显多于其他激素,而GA和SA是葡萄种子发育的关键激素,特别是与无核葡萄果实的发育密切相关[19,20],表明miR171s及靶基因可能通过响应激素信号参与无核葡萄的发育过程。
2.7 vvi-miR171s及靶基因在葡萄种子发育不同时期的时空表达特征
2.7.1 在葡萄种子不同发育时期时空表达分析 如图7所示,根据表达量水平可将vvi-miR171s分为3类,vvi-miR171a为最高,其次是vvi-miR171b/e/f,vvi-miR171g几乎不表达,未检测到vvi-miR171g;靶基因中仅检测到VvSCL15/VvSCL22,未检测到VvSCL6。VvSCL15/VvSCL22呈现完全相反的表达模式,VvSCL15在20 DAF检测到最高水平,在30 DAF最低;而VvSCL22则在20 DAF未检测到表达量,而在30 DAF最高。vvi-miR171s中除vvi-miR171b在20 DAF检测到最高水平外,其他成员均在45 DAF检测到最高水平。vvi-miR171s与VvSCL15/VvSCL22相关性分析发现(图7),vvi-miR171a/f-VvSCL15(r=-0.82/-0.98)、vvi-miR171b-VvSCL22(r=-1.00)具有显著负相关性,而vvi-miR171b-VvSCL15(r=1.00)和vvi-miR171a/f-VvSCL22则为显著正相关(r=0.83/0.98),这一结果也为vvi-miR171s成员以不同的途径和作用方式调节靶基因进而调控种子发育提供了证据;vvi-miR171a/f-VvSCL15、vvi-miR171b-VvSCL22在葡萄种子发育过程中均存在负调控关系,结合表达水平认为vvi-miR171a-VvSCL15为调控种子发育过程的主要成员。图7
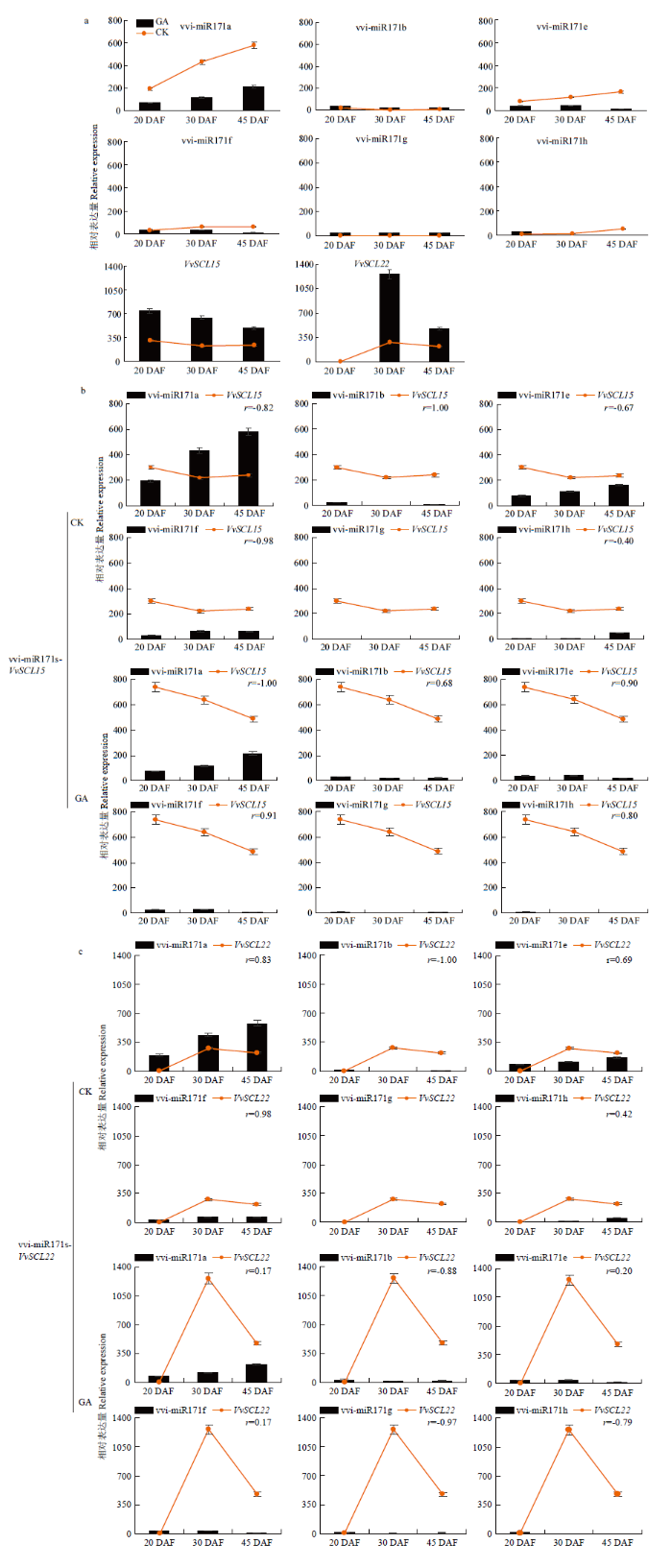 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7vvi-miR171s在葡萄种子/种子区的时空表达分析(a)、应答GA3的表达模式(b)及vvi-miR171s对靶基因的作用模式(c)
Fig. 7Spatio-temporal expression analysis of vvi-miR171s in grape seed/seed region (a), expression pattern in response to GA3 (b) and action pattern of vvi-miR171s on target gene (c)
2.7.2 vvi-miR171s及靶基因应答GA3的模式分析 为进一步认识葡萄vvi-miR171s及其靶基因在种子区对外源GA3的时空响应模式,利用对照和GA3处理的种子区样品cDNA为模板,进行定量分析。图7-a为对照与GA3处理后的葡萄种子区vvi-miR171s及VvSCL15/VvSCL22的表达水平。GA3处理后种子区vvi-miR171s响应GA3的模式可分为2类,vvi- miR171a/e/f/h的表达量显著下调,而vvi-miR171b/g则在GA3处理后上调;与CK相比,VvSCL15/22的表达量均显著升高。同时相关性分析表明(图7-b、c),GA3处理后,vvi-miR171a-VvSCL15(r=-1.00)、vvi- miR171b/g/h-VvSCL22(r=-0.88/-0.97/-0.79)呈显著负相关;而GA3处理后种子区vvi-miR171f-VvSCL15由显著负相关性(r=-0.98)变为显著正相关(r=0.91),同时vvi-miR171/f/h-VvSCL15(r=0.91、0.80)为显著正相关,这表明vvi-miR171s在种子发育过程中以不同的模式响应赤霉素信号。
由于miRNA家族中各个成员的作用可能存在冗余现象,因此利用vvi-miR171s的累计表达量与靶基因的表达量进行分析(图8)。值得注意的是,GA3强烈抑制了种子区vvi-miR171s的表达(图8),其中vvi-miR171a(图7-b)的下调程度远超过其他成员,表明在葡萄种子区,vvi-miR171a可能是该家族中应答GA3的主要成员。同时发现,GA3显著上调种子区的VvSCL15和VvSCL22,且GA3处理后,vvi-miR171a-VvSCL15呈负调控关系,因而推测vvi-miR171a负调控VvSCL15是vvi-miR171s响应赤霉素信号调控种子发育的主要途径。
图8
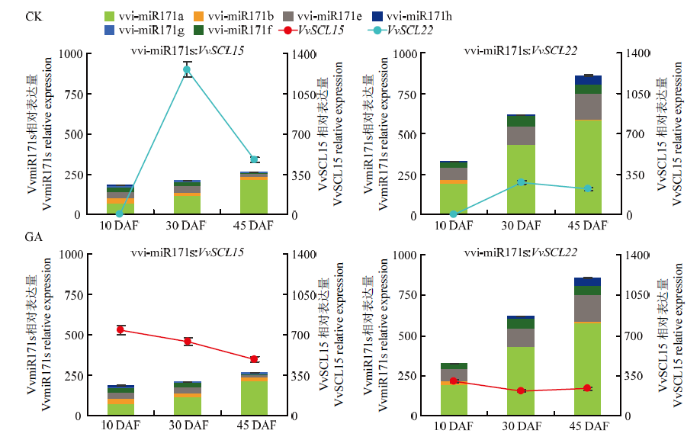 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8种子/种子区中vvi-miR171s累计表达量与靶基因的时空表达分析、作用模式及应答GA3的模式分析
Fig. 8Total expression of vvi-miR171s and their VvSCL target genes during seed/seed region development and their patterns in response to GA3
3 讨论
据报道,不同物种中miR171s的功能不完全一致。在拟南芥[15]、大麦[13]和烟草[21]中发现miR171c调控腋生分生组织形成;落叶松LxmiR171在胚性细胞中过表达后,可促进诱导胚性愈伤组织[22],同时LxmiR171a/b和LxmiR171c还可以调控原胚和体胚诱导阶段[23];百合Lpu-miR171a[24]在胚性愈伤组织阶段表达量最高,而Lpu-miR171b、Lda-miR171a和Lda-miR171c的表达量均在鱼雷形胚阶段出现最高值;甜橙miR171在胚性愈伤组织中有高表达,主要在柑橘细胞胚性诱导和保持过程中发挥作用[1];vvi-miR171s对葡萄花芽分化具有一定作用[25];利用STTM和CRISPR/Cas9技术,发现水稻miR171a参与叶绿素、脯氨酸的合成过程,并对植物生长势和抗旱能力有调节作用[26];SlmiR171b不仅在番茄花粉萌发过程中起负调控作用,也参与了番茄的呼吸跃变过程,并负调控果实中葡萄糖、果糖和番茄红素等营养物质的积累[27];而LIN等[6]发现杏(Prunus sibirica L.)PsmiR171-SCL与种子发育相关,且不同成员在种子不同发育时期发挥重要作用。本研究的靶裂解位点结果表明,VvSCL6、VvSCL15和VvSCL22均为vvi-miR171s的靶基因,但对不同时期种子区cDNA进行定量检测,未检测到VvSCL6的表达量,这可能是由于VvSCL6表达量很弱或具有组织、发育时期特异性。与葡萄中的其他microRNAs类似[28,29],miR171s也具有时空特异的表达模式。本研究发现,在葡萄硬核期种子区vvi-miR171a相比其他成员表达量较高,vvi-miR171a可能作为主要成员影响种子发育,说明葡萄miR171s的功能可能与拟南芥、落叶松和柑橘中的存在差异性,葡萄中极可能存在特异性的调控途径。同时,类似于拟南芥AtmiR171c[22,30]及百合Lpu-miRl71a/b[24],葡萄vvi-miR171a与靶基因VvSCL15之间也存在明显的负调控的关系。本研究利用3′-PPM-RACE和5′-RLM-RACE技术验证了vvi-miR171a/b/e/f/g/h对VvSCL6/15/22的裂解作用,这与miR171s在拟南芥[31]、落叶松[5]和龙眼[4]中通过对VvSCL15和VvSCL22的转录产物切割而进行调控的结论一致,验证了这3条靶基因的真实性;同时确定了VvSCL22的裂解位点均位于miR171a/b/e/f/g/h的5′端第9位和第10位碱基之间所匹配的靶点上,与拟南芥[31]中miR171靶基因的裂解位点一致,说明miR171s对靶基因SCL裂解作用位点的保守性。
研究发现miR171及靶基因SCL6可参与GA信号转导,GA3可上调龙眼miR171b和SCL6的表达进而影响胚性细胞的发育;MA等[12]发现miR171-SCL受拟南芥GA-DELLA信号途径调节,在叶绿素合成中起关键调控作用,这些研究均证明miR171-SCL可能通过对GA信号的响应调控植物生长进程。与龙眼[4]不同,GA3显著下调了葡萄种子区vvi-miR171a的表达,而上调了VvSCL15和VvSCL22;同时,vvi-miR171a与VvSCL15存在负调控关系,结合GA3高效诱导葡萄无核的效果,及vvi-miR171a可能是miR171s在硬核期调控种子发育的主要成员,推测赤霉素信号通过下调vvi-miR171a影响靶基因VvSCL15的表达,最终影响葡萄种子的发育。本研究初步鉴定了vvi-miR171s-VvSCL15可能是vvi-miR171s调控靶基因,响应赤霉素信号参与葡萄种子发育的主要途径,这将为进一步研究vvi-miR171s及靶基因响应赤霉素信号调控葡萄种子发育的分子机制研究奠定重要基础。
4 结论
在‘魏可’葡萄中鉴定到vvi-miR171a/b/e/f/g/h 6个成员,并利用3′-PPM-RACE和5′-RLM-RACE技术验证了它们对3个靶基因VvSCl6/15/22的裂解作用及裂解位点。通过对vvi-miR171s和VvSCLs在葡萄种子中的时空表达模式分析,发现vvi-miR171a-VvSCL15可能作为主要的调控途径响应赤霉素信号并参与调控葡萄种子发育。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1007/s00425-010-1312-9URL [本文引用: 3]
DOI:10.1007/s11105-014-0700-xURL [本文引用: 1]

microRNAs (miRNAs), endogenous non-coding RNAs of approximately 21-24 nucleotides, are important regulators of transcriptional and post-transcriptional gene expression. These regulators play a key role during plant growth and development, including embryogenesis, which is crucial to the life cycle of most plant species. However, although embryogenesis-associated miRNAs have been isolated in a few species, the diversity of these regulatory miRNAs remains largely unexplored, especially in radish. In this study, two small RNA libraries were constructed from radish ovules before and after fertilization. Both libraries were sequenced by next generation sequencing (NGS) technology. This analysis identified 144 conserved and 34 non-conserved miRNAs (representing 43 known miRNA families) and 38 novel miRNAs (representing 28 miRNA families). Comparative analysis revealed that 29 known and 10 novel miRNA families were differentially expressed during embryogenesis. QRT-PCR analysis confirmed miRNA expression patterns and revealed tissue-specific and/or developmental stage-dependent expression for some miRNAs. Moreover, potential target predictions indicated that most of these targets were transcription factors involved in regulating plant growth, development and metabolism. Notably, target transcripts such as squamosa promoter-binding protein, auxin response factor and agamous-like MADS-box protein participated in radish embryogenesis.
[D].
[本文引用: 3]
[D].
[本文引用: 3]
[本文引用: 5]
[本文引用: 5]
URL [本文引用: 2]
URLPMID:27762296 [本文引用: 3]
DOI:10.1007/s11240-017-1249-9URL [本文引用: 1]
DOI:10.1016/S0092-8674(00)80115-4URL [本文引用: 1]
URLPMID:18662674 [本文引用: 1]
DOI:10.1016/j.febslet.2009.02.024URLPMID:19236868 [本文引用: 1]
MicroRNAs (miRNAs) are small RNAs acting as regulators of eukaryotic gene expression at the post-transcriptional level. Plant miRNAs have been implicated in developmental processes and adaptation to the environment. We show that the accumulation of four Arabidopsis miRNAs (miR171, miR398, miR168 and miR167) oscillates during the diurnal cycle, their accumulation increasing during the light period of the daytime and decreasing in darkness. This oscillatory pattern of miRNA accumulation is not governed by the circadian clock. These results suggest a potential role of light in controlling miRNA accumulation while defining a new level of regulation of miRNA gene expression in Arabidopsis.
DOI:10.1007/s11105-014-0787-0URL [本文引用: 1]
DOI:10.1371/journal.pgen.1004519URLPMID:25101599 [本文引用: 2]
An extraordinarily precise regulation of chlorophyll biosynthesis is essential for plant growth and development. However, our knowledge on the complex regulatory mechanisms of chlorophyll biosynthesis is very limited. Previous studies have demonstrated that miR171-targeted scarecrow-like proteins (SCL6/22/27) negatively regulate chlorophyll biosynthesis via an unknown mechanism. Here we showed that SCLs inhibit the expression of the key gene encoding protochlorophyllide oxidoreductase (POR) in light-grown plants, but have no significant effect on protochlorophyllide biosynthesis in etiolated seedlings. Histochemical analysis of beta-glucuronidase (GUS) activity in transgenic plants expressing pSCL27::rSCL27-GUS revealed that SCL27-GUS accumulates at high levels and suppresses chlorophyll biosynthesis at the leaf basal proliferation region during leaf development. Transient gene expression assays showed that the promoter activity of PORC is indeed regulated by SCL27. Consistently, chromatin immunoprecipitation and quantitative PCR assays showed that SCL27 binds to the promoter region of PORC in vivo. An electrophoretic mobility shift assay revealed that SCL27 is directly interacted with G(A/G)(A/T)AA(A/T)GT cis-elements of the PORC promoter. Furthermore, genetic analysis showed that gibberellin (GA)-regulated chlorophyll biosynthesis is mediated, at least in part, by SCLs. We demonstrated that SCL27 interacts with DELLA proteins in vitro and in vivo by yeast-two-hybrid and coimmunoprecipitation analysis and found that their interaction reduces the binding activity of SCL27 to the PORC promoter. Additionally, we showed that SCL27 activates MIR171 gene expression, forming a feedback regulatory loop. Taken together, our data suggest that the miR171-SCL module is critical for mediating GA-DELLA signaling in the coordinate regulation of chlorophyll biosynthesis and leaf growth in light.
DOI:10.1186/1471-2229-13-6URLPMID:23294862 [本文引用: 2]
BACKGROUND: The transitions from juvenile to adult and adult to reproductive phases of growth are important stages in the life cycle of plants. The regulators of these transitions include miRNAs, in particular miR156 and miR172 which are part of a regulatory module conserved across the angiosperms. In Arabidopsis miR171 represses differentiation of axillary meristems by repressing expression of SCARECROW-LIKE(SCL) transcription factors, however the role of miR171 has not been examined in other plants. RESULTS: To investigate the roles of mir171 and its target genes in a monocot, the Hvu pri-miR171a was over-expressed in barley (Hordeum vulgare L. cv. Golden promise) leading to reduced expression of at least one HvSCL gene. The resulting transgenic plants displayed a pleiotropic phenotype which included branching defects, an increased number of short vegetative phytomers and late flowering. These phenotypes appear to be the consequence of changes in the organisation of the shoot meristem. In addition, the data show that miR171 over-expression alters the vegetative to reproductive phase transition by activating the miR156 pathway and repressing the expression of the TRD (THIRD OUTER GLUME) and HvPLA1 (Plastochron1) genes. CONCLUSIONS: Our data suggest that some of the roles of miR171 and its target genes that have been determined in Arabidopsis are conserved in barley and that they have additional functions in barley including activation of the miR156 pathway.
[本文引用: 1]
DOI:10.1093/mp/ssq042URLPMID:20720155 [本文引用: 3]
MicroRNAs (miRNAs) are approximately 21-nucleotide noncoding RNAs that play critical roles in regulating plant growth and development through directing the degradation of target mRNAs. Axillary meristem activity, and hence shoot branching, is influenced by a complicated network that involves phytohormones such as auxin, cytokinin, and strigolactone. GAI, RGA, and SCR (GRAS) family members take part in a variety of developmental processes, including axillary bud growth. Here, we show that the Arabidopsis thaliana microRNA171c (miR171c) acts to negatively regulate shoot branching through targeting GRAS gene family members SCARECROW-LIKE6-II (SCL6-II), SCL6-III, and SCL6-IV for cleavage. Transgenic plants overexpressing MIR171c (35Spro-MIR171c) and scl6-II scl6-III scl6-IV triple mutant plants exhibit a similar reduced shoot branching phenotype. Expression of any one of the miR171c-resistant versions of SCL6-II, SCL6-III, and SCL6-IV in 35Spro-MIR171c plants rescues the reduced shoot branching phenotype. Scl6-II scl6-III scl6-IV mutant plants exhibit pleiotropic phenotypes such as increased chlorophyll accumulation, decreased primary root elongation, and abnormal leaf and flower patterning. SCL6-II, SCL6-III, and SCL6-IV are located to the nucleus, and show transcriptional activation activity. Our results suggest that miR171c-targeted SCL6-II, SCL6-III, and SCL6-IV play an important role in the regulation of shoot branch production.
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1399-3054.2011.01481.xURLPMID:21496033 [本文引用: 1]
MicroRNAs (miRNAs) are a class of non-coding RNA molecules which have significant gene regulation roles in organisms. The advent of new high-throughput sequencing technologies has enabled the discovery of novel miRNAs. Although there are two recent reports on high-throughput sequencing analysis of small RNA libraries from different organs of two wine grapevine varieties, there was a significant divergence in the number and kinds of miRNAs sequenced in these studies. More sequencing of small RNA libraries is still important for the discovery of novel miRNAs in grapevine. In this study, a total of 130 conserved grapevine Vitis vinifera miRNA (Vv-miRNA) belonging to 28 Vv-miRNA families were validated, other 80 unconserved Vv-miRNAs including 72 novel potential and 8 known but unconserved ones were found. Fifty-two (52.5%) of these 80 unconserved Vv-miRNAs exhibited differential poly(A)-tailed reverse transcriptase-polymerase chain reaction expression profiles in various grapevine tissues that could further confirm their existence in grapevine, among which 20 were expressed only in grapevine berries, indicating a degree of fruit-specificity. One hundred thirty target genes for 56 unconserved miRNAs could be predicted. The locations of these potential target genes on grapevine chromosomes and their complementary levels with the corresponding miRNAs were also analyzed. These results point to a regulatory role of miRNAs in grapevine berry development and response to various environments.
DOI:10.1016/j.jplph.2013.02.005URLPMID:23582890 [本文引用: 1]
MicroRNAs (miRNAs) regulate target gene expression by mediating target gene cleavage or inhibition of translation at transcriptional and post-transcriptional levels in higher plants. Until now, many grapevine microRNAs (Vv-miRNAs) have been identified and quite a number of miRNA target genes were also verified by various analysis. However, global interaction of miRNAs with their target genes still remained to perform more research. We reported experimental validation of a number of miRNA target genes in table grapevine that had been previously identified by bioinformatics in our earlier studies. To verify more predicted target genes of Vv-miRNAs and elucidate the modes by which these Vv-miRNAs work on their target genes, 31 unverified potential target genes for 18 Vv-miRNAs were experimentally verified by a new integrated strategy employing a modified 5'-RLM-RACE (RNA ligase-mediated 5' rapid amplification of cDNA ends), 3'-PPM-RACE (poly(A) polymerase-mediated 3' rapid amplification of cDNA ends) and qRT-PCRs of cleavage products. The results showed that these Vv-miRNAs negatively regulated expression of their target messenger RNAs (mRNAs) through guiding corresponding target mRNA cleavage, of which about 94.4% Vv-miRNAs cleaved their target mRNAs mainly at the tenth nucleotide of 5'-end of miRNAs. Expression levels of both miRNAs and their target mRNAs in eight tissues exhibited inverse relationships, and expressions both of cleaved targets and miRNAs indicated a cleavage mode of Vv-miRNAs on their target genes. Our results confirm the importance of Vv-miRNAs in grapevine growth and development, and suggest more study on Vv-miRNAs and targets can enrich the knowledge of miRNA mediated-regulation in grapevine.
URLPMID:25888129 [本文引用: 1]
DOI:10.1371/journal.pone.0080044URLPMID:24224035 [本文引用: 1]
BACKGROUND: The phytohormone gibberellic acid (GA3) is widely used in the table grape industry to induce seedlessness in seeded varieties. However, there is a paucity of information concerning the mechanisms by which GAs induce seedlessness in grapes. METHODOLOGY/PRINCIPAL FINDINGS: In an effort to systematically analyze the cause of this GA3-induced seed abortion, we conducted an in depth characterization of two seeded grape cultivars ('Kyoho' and 'Red Globe'), along with a seedless cultivar ('Thompson Seedless'), following treatment with GA3. In a similar fashion to the seedless control, which exhibited GA3-induced abortion of the seeds 9 days after full bloom (DAF), both 'Kyoho' and 'Red Globe' seeded varieties exhibited complete abortion of the seeds 15 DAF when treated with GA3. Morphological analyses indicated that while fertilization appeared to occur normally following GA3 treatment, as well as in the untreated seedless control cultivar, seed growth eventually ceased. In addition, we found that GA3 application had an effect on redox homeostasis, which could potentially cause cell damage and subsequent seed abortion. Furthermore, we carried out an analysis of antioxidant enzyme activities, as well as transcript levels from various genes believed to be involved in seed development, and found several differences between GA3-treated and untreated controls. CONCLUSION: Therefore, it seems that the mechanisms driving GA3-induced seedlessness are similar in both seeded and seedless cultivars, and that the observed abortion of seeds may result at least in part from a GA3-induced increase in cell damage caused by reactive oxygen species, a decrease in antioxidant enzymatic activities, and an alteration of the expression of genes related to seed development.
URL [本文引用: 1]
URL [本文引用: 1]
URLPMID:20599742 [本文引用: 2]
URLPMID:22526500 [本文引用: 1]
[D].
[本文引用: 2]
[D].
[本文引用: 2]
[D].
[本文引用: 1]
URL [本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
[本文引用: 2]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}