 ,1, 罗丁峰1, 柏浩东1, 徐晶晶1, 韩进财1, 徐强2, 王若仲2, 柏连阳,1,2
,1, 罗丁峰1, 柏浩东1, 徐晶晶1, 韩进财1, 徐强2, 王若仲2, 柏连阳,1,2Cloning and Expression Analysis of Light Harvesting Chlorophyll a/b Protein Gene CcLhca-J9 in Conyza canadensis
LI ZuRen,1, LUO DingFeng1, BAI HaoDong1, XU JingJing1, HAN JinCai1, XU Qiang2, WANG RuoZhong2, BAI LianYang,1,2通讯作者:
责任编辑: 岳梅
收稿日期:2020-04-12接受日期:2020-06-28网络出版日期:2021-01-01
| 基金资助: |
Received:2020-04-12Accepted:2020-06-28Online:2021-01-01
作者简介 About authors
李祖任,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (1164KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
李祖任, 罗丁峰, 柏浩东, 徐晶晶, 韩进财, 徐强, 王若仲, 柏连阳. 小飞蓬捕光叶绿素结合蛋白基因CcLhca-J9克隆及表达分析[J]. 中国农业科学, 2021, 54(1): 86-94 doi:10.3864/j.issn.0578-1752.2021.01.007
LI ZuRen, LUO DingFeng, BAI HaoDong, XU JingJing, HAN JinCai, XU Qiang, WANG RuoZhong, BAI LianYang.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】小飞蓬(Conyza canadensis)为菊科(Asteraceae)飞蓬属(Conyza)的一种具较强繁殖和环境适应能力的入侵杂草,主要分布于果、桑、茶园、草坪和荒地[1]。小飞蓬已对草甘膦、草铵膦、莠去津等多种化学除草剂产生不同程度的抗药性,例如浙江宁波的两个抗草甘膦种群抗性指数分别达8.28和7.92[2,3]。据报道,当小飞蓬1 m2发生数为72株时,油菜角果数、角果籽粒数显著下降,产量下降达35.93%,可见恶性杂草小飞蓬给农业生产带来较大的经济损失[4]。植物化感物质的挖掘和利用是开发植物源除草剂的有效途径,是治理小飞蓬等抗药性杂草的热点方向之一[5]。发掘新颖生物源化合物结构、除草作用机制和靶标,阐明活性化合物与杂草相互作用机制,利用生物除草关键基因资源,可最终克服生物除草剂产品发展的成本与环境制约因素[6]。【前人研究进展】目前,以天然产物为基础研发的生物除草剂的作用机制主要有五大类,但是许多天然产物的抑草作用靶点和分子机制仍然是未知的[7]。光合系统是除草剂作用的重要靶标之一,其中光系统I和光系统II是吡啶类和三嗪酮类除草剂的靶标[8]。链霉菌(Streptomyces spp.)产生的多肽L-phosphinothricin抑制谷氨酰胺合成酶,链格孢菌(Alternari alternata)产生的细交链孢菌酮酸(tenuazonic acid,TeA)阻断光合系统II[9,10]。上述抑草靶标的发现,为生物除草剂的开发提供了科学依据。羊脂酸分子式为C8H16O2,油状液体不溶于水,属于有机酸类化合物,主要用于染料、防腐剂、杀菌剂[11]。笔者所在课题组前期从椰子中分离提取出羊脂酸,并验证其为一种具有广谱、高效抑草活性的植物源化合物,具有开发成为植物源除草剂的潜力[12]。采用透射电镜技术观察到羊脂酸处理小飞蓬叶片后,叶绿体和线粒体结构随着作用时间的延长出现了空腔、裂解的现象[13]。采用Lable-free蛋白质组学技术分析羊脂酸处理小飞蓬叶片,发现LHC蛋白(UniProt ID:A0A103Y9J9)响应了羊脂酸处理,并与光合指标测定结果相一致,但该蛋白是否为羊脂酸抑制小飞蓬的靶标蛋白仍需进一步研究[14]。捕光叶绿素a/b结合蛋白(light harvesting chlorophyll a/b-binding protein,LHC)是光系统I和光系统II中获取光能并将其传送至光反应中心的色素蛋白复合体[15]。在高等植物体内,编码LHC(LHCI、LHCII)蛋白是一个多基因家族,定位于细胞核内;LHCI蛋白由Lhcal、Lhca2、Lhca3和Lhca4基因编码,LHCII蛋白由Lhcbl、Lhcb2、Lhcb3、Lhcb4、Lhcb5和Lhcb6基因编码[16]。随着分子克隆和基因测序技术的发展,部分植物的LHC蛋白基因已被克隆出来,例如拟南芥(Arabidopsis thaliana)、橡胶树(Hevea brasiliensis)、籼稻9311(Oryza sativa)[17,18,19]。【本研究切入点】目前的研究大部分集中在各植物LHC蛋白及其基因的表达差异、功能解析等,对于LHC蛋白与抑草活性胁迫相关的功能解析研究较少[20]。【拟解决的关键问题】通过分子克隆和生物信息学等技术全面阐释小飞蓬捕光叶绿素a/b结合蛋白基因(CcLhca-J9)的分子特征,解析羊脂酸处理下该基因表达的特殊性,预测羊脂酸作用LHC蛋白的关键位点,为后续基因功能验证打下基础,同时为植物源羊脂酸产品研发提供科学依据。1 材料与方法
1.1 供试材料
小飞蓬种子2018年9月采集于湖南省长沙市芙蓉区湖南省农业科学院院内荒地并保存于种子超干低湿存储柜(购于北京明日百傲生物科技有限公司),选出饱满种子用0.4%赤霉素溶液催芽后,挑选吐白的种子移入塑料盆钵(内径20 cm),置于光照培养箱(8 h黑暗、温度18℃/16 h光照、光强100—120 μmol·m-2·s-1、温度20℃)培养至三叶期待用。羊脂酸(纯度99%)采用蒸馏萃取法自制,具体制备方法参考文献[12]。1.2 CcLhca-J9的克隆
从NCBI中选择保守性高的Lhca基因同源序列设计简并引物CcJ9-1(表1),进行PCR扩增获取目的基因片段。PCR反应程序:94℃预变性5 min;94℃变性30 s,55℃退火30 s,70℃延伸30 s,循环30次;70℃延伸7 min。凝胶电泳检测PCR产物,切胶回收纯化目的片段,连接到pCambia2301载体,连接转化大肠杆菌DH5α(购于南京诺唯赞生物科技有限公司),挑取阳性克隆送上海生工生物工程有限公司测序[17]。根据已知目的片段设计5′和3′ RACE特异引物(表1),采用TaKaRa公司Clonetech SMARTer RACE 5′/3′ Kit扩增试剂盒说明扩增。获得Lhca的5′ RACE序列、核心序列及3′ RACE序列,根据三者拼接后序列设计引物(表1)确定CcLhca-J9全长。Table 1
表1
表1CcLhca-J9克隆及实时荧光定量PCR表达引物
Table 1
| 引物名称 Primer name | 引物序列 Primer sequence (5′-3′) | 引物用途 Use of primers |
|---|---|---|
| CcJ9-1 | Forward: GAGYTGCGGYATCBCCGCCGT Reverse: GTTGTGCCATGGRTCAGCCAA | 核心片段扩增 Fragment cloning |
| CcJ9-F2 | Forward: CGGATTTGACCCACTTCGTCT | 5′ RACE扩增 5′ cDNA end amplification |
| CcJ9-R2 | Reverse: TATTGCCAGTATGGTGGGTAGGGT | 5′ RACE扩增 5′ cDNA end amplification |
| CcJ9-R3 | Reverse: GGCAGCCCATTCTTGTGCTTTTAC | 5′ RACE扩增 5′ cDNA end amplification |
| CcJ9-F3 | Forward: GGCAACTGGGTAAAAGCACAAGAAT | 3′ RACE扩增 3′ cDNA end amplification |
| CcJ9-F4 | Forward: AAGACCCAAAAACATTCGCAGAGTA | 3′ RACE扩增 3′ cDNA end amplification |
| qR-J9 | Forward: CATTCTTGTGCTTTTACCCAG Reverse: GAGGTTTAAAGAGTCCGAGCT | 实时荧光定量PCR RT-qPCR |
| ACTIN | Forward: CCGATCCAGACGCTGTATTT Reverse: TGCTGATCGTATGAGCAAGG | 实时荧光定量PCR RT-qPCR |
新窗口打开|下载CSV
1.3 生物信息学分析
采用DNAMAN分析核酸序列并检测编码氨基酸序列,ExPaSy在线预测分子量和等电点(molecular weight/isoelectric point,MW/pI),在NCBI中搜索LHC的高相似度氨基酸序列,选取19个典型的植物蛋白,先用ClustalX软件多序列比对,再用MEGA 4.1软件的邻接法(neighbor-joining,NJ)构建进化树[21]。利用SWISS-MODEL在线搜索与选择模板,再通过Swiss-PdbViewer软件优化模型结构[22]。1.4 分子对接
采用上一步LHC同源建模结果作为模型来寻找羊脂酸与LHCI-J9蛋白的对接方式。在AutoDock 4.2软件上运行,电荷计算采用RESP(Restrained electrostatic potential)法;活性位点采用AutoGrid格式点搜索,网格大小为60?×40?×60?,格点间隔为1.0?。中心设定为配体在LHC中的质心;采用LGA(Lamarckian genetic algorithm)法计算配体与受体间的自由能,其余参数均为缺省值[23]。在最终得到的10个最优构象中,选取能量值最低的构象作为进一步研究对象。1.5 CcLhca-J9响应羊脂酸的表达分析
用羊脂酸(625 μmol·L-1)、阿魏酸(625 μmol·L-1)和清水对照各10 μL,兑水10 mL,喷雾处理小飞蓬叶片0、0.5、1、2、4、8 h后,收集新鲜叶片立即置于液氮。参照Trizol改良法提取小飞蓬叶片总RNA,反转录合成第一链cDNA。RT-qPCR反应体系20 μL:SYBR Premix ExTaqTM 10 μL、引物(R和F)0.5 μL、cDNA 5 μL、ddH2O 4 μL。反应程序:95℃ 30 s,58℃ 20 s,循环40 次。所用引物为目的基因特异性引物qR-J9和内参基因Actin引物(表1)。数据处理运用2-ΔΔCT法,比较目的基因的表达水平[24]。利用SPSS软件中Duncan氏新复极差法进行差异显著性分析。2 结果
2.1 CcLhcIa-J9序列的获得
以反转录未施药的小飞蓬叶片cDNA为模板,用简并引物CcJ9-1进行PCR扩增、回收和测序,获得CcLhca-J9核心序列483 bp(图1-A)。用引物Cc-F2、Cc-R2、Cc-R3、Cc-F3和Cc-F4进行三轮PCR扩增,产物经回收和测序,获得cDNA 5′端序列428 bp和3′端序列253 bp(图1-B)。将5′ RACE序列、核心序列及3′ RACE序列用DNAMAN软件拼接,获得CcLhca-J9 cDNA序列全长(图2)。图1
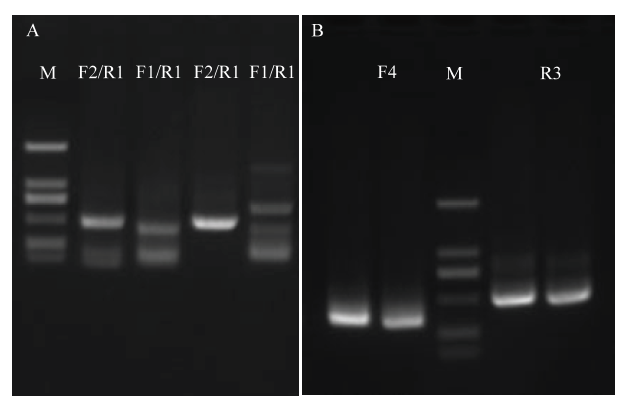 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1CcLhca-J9扩增
A:CcLhca-J9核心序列PCR扩增Amplification of CcLhca-J9 core sequence;B:CcLhca-J9 RACE PCR扩增Amplification of CcLhca-J9 RACE
Fig. 1Amplification of CcLhca-J9
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2CcLhca-J9基因核苷酸及编码的氨基酸序列
Fig. 2CcLhca-J9 gene nucleotide and the encoded amino acid sequences
2.2 CcLhcIa-J9生物信息学分析
经同源克隆和RACE克隆,获得了CcLhca-J9的全长cDNA序列,该基因阅读框长744 bp,编码247个氨基酸(图2)。编码蛋白分子质量为26.766 kD,理论等电点为6.43;该蛋白中25个丙氨酸,占总数的10.1%;其次是脯氨酸和甘氨酸,分别为24和23个,各占9.7%和9.3%。CcLhca-J9蛋白保守区预测显示,该蛋白属于Chloroa_b-bind家族,具有典型的捕光叶绿素a/b结合蛋白功能(图3)。系统进化结果表明,不同物种来源的Lhca蛋白在分属不同分支,CcLhca-J9蛋白与除虫菊(GEW73959.1,Tanacetum cinerariifolium)和黄花蒿(PWA35049.1, Artemisia annua)Lhca蛋白进化程度最为接近,同处于菊科这一分支(图4)。预测二级结构发现,蛋白具有螺旋(Hh)97个,占39.27%;β转角(Tt)7个,占2.83%;延伸链(Ee)9个,占3.64%;无规则卷曲(Cc)134个,占54.25%(图5)。LHC-J9蛋白以4y28.1.O(2.80?)为模板进行同源建模,序列相似度为89.9%,高度同源结构,GEQE=0.75,QMEAN=-2.20,可见所获得的模型质量是较为可靠的。CcLhca-J9蛋白是单分子物体,具有6个叶绿体a配体,是一个典型的捕光复合物I叶绿素a/b结合蛋白(图6)。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3CcLhca-J9与同源基因编码区氨基酸序列比对
同源氨基酸及其登录号The source and accession number of the amino acids are as follows: TcLhca (Tanacetum cinerariifolium, GEW73959.1), AaLhca (Artemisia annua, PWA35049.1), HaLhca (Helianthus annuus, XP021976973.1), LsLhca (Lactuca sativa, XP023729531.1), CcLhca (Conyza canadensis)
Fig. 3Multiple sequence alignment of CcLhca-J9 and homologous gene coding region
图4
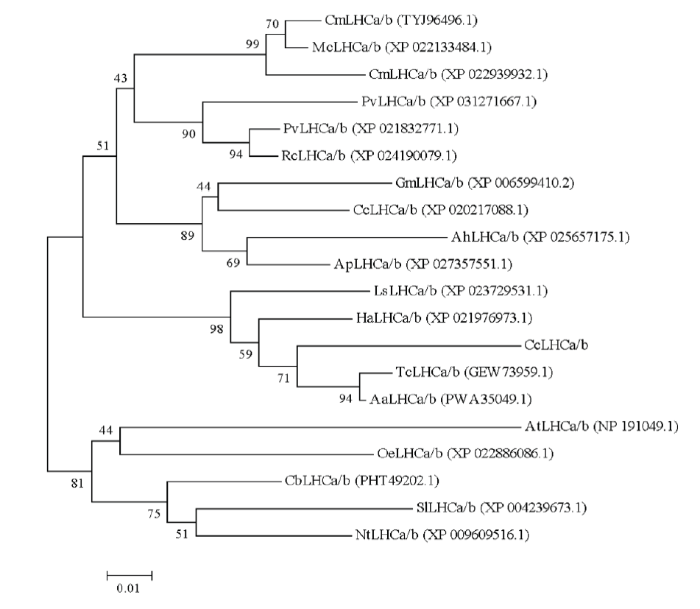 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4CcLhca-J9蛋白与近缘种群的系统进化树(邻接法,1 000次重复)
Fig. 4Phylogenetic tree of CcLhca-J9 protein with homologous proteins from other species (neighbor-joining method, 1 000 replicates)
图5
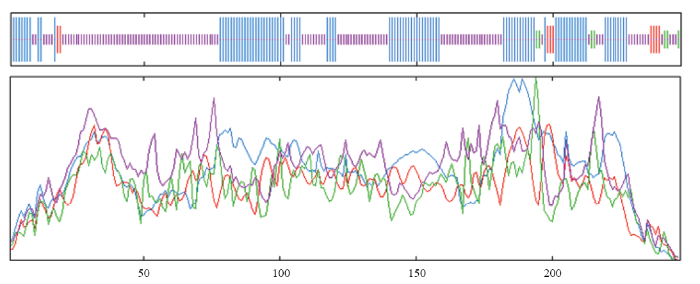 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5CcLhca-J9蛋白二级结构
蓝色Blue:α-螺旋Alpha helix;紫色Purple:无规则卷曲Random coil;红色Red:延伸链Extended strand;绿色Green:β-转角Beta turn
Fig. 5Secondary structure of CcLhca-J9
图6
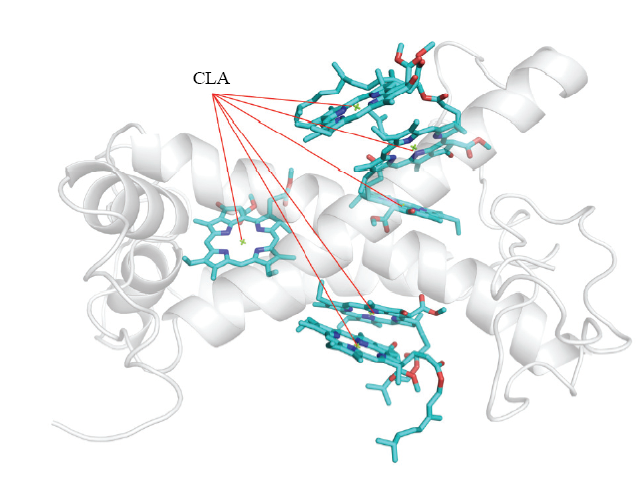 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6CcLhca-J9蛋白同源三级结构预测模型
CLA:叶绿素a配体(6个,带不同颜色)
Fig. 6Homology model structure of CcLhca-J9
6x CLA (chlorophyll a) ligands were shown with different colors
2.3 分子对接
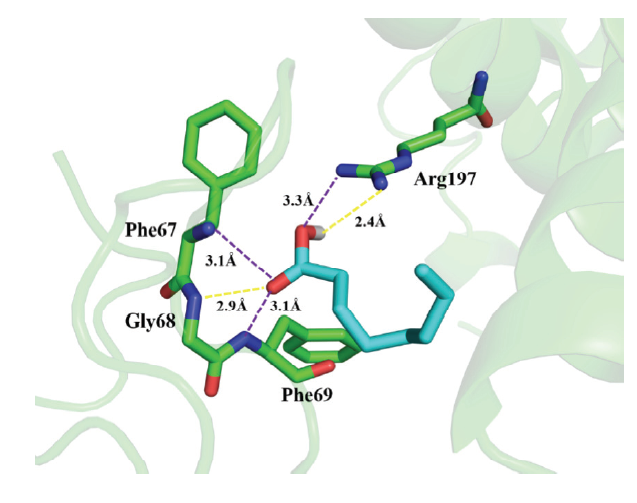
分子对接的计算结果显示(图7),羊脂酸与CcLhca-J9蛋白形成氢键和p-π的作用力共同影响了两者之间的结合,其结合能量为-20.9 kJ·mol-1。羊脂酸的羰基氧原子与Gly68氨基酸残基形成氢键,键长为2.9?;Phe67和Phe69中氮原子与羊脂酸的碳氧双键形成p-π,距离均为3.1?;羊脂酸的羟基氢原子与Arg197氨基酸残基形成氢键,键长为2.4?;羊脂酸的羟基氧原子与Arg197氨基酸残基形成p-π,键长为3.3?。图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7羊脂酸与CcLhca-J9的分子对接模式图
黄色虚线Yellow imaginary line:氢键Hydrogen bond;紫色虚线Purple imaginary line:p-π
Fig. 7Molecular docking schema chart between caprylic acid and CcLha-J9
2.4 羊脂酸处理对CcLhca-J9 表达的影响
RT-qPCR结果表明,在羊脂酸处理小飞蓬叶片后0—8 h,CcLhca-J9表达量明显下降。羊脂酸处理后0.5—1 h,CcLhca-J9的表达量与清水对照相比下降了66.01%;羊脂酸处理后2 h,CcLhca-J9的表达量缓慢下降,与对照相比下降了82.72%;4—8 h 后,CcLhca-J9的表达量趋于稳定,与对照相比下降92%—93%(图8)。羊脂酸处理后,CcLhca-J9表达量与对照相比呈下调趋势,此结果与前期蛋白质组学结果一致。清水对照和另一有机酸类化合物阿魏酸处理小飞蓬叶片,CcLhca-J9表达量总体未出现显著变化。图8
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8羊脂酸处理后不同时间CcLhca-J9的相对表达量
Fig. 8Relative expression level of CcLhca-J9 after caprylic acid treatment
3 讨论
植物LHC蛋白家族基因是细胞核内的光合系统基因,其编码的蛋白与色素形成的复合物能捕获光能并传递能量至光化学反应中心[16]。本研究采用RACE技术克隆获得小飞蓬叶片LHC蛋白家族基因成员CcLhca-J9 cDNA序列,其OFR全长为744 bp,编码247个氨基酸,分子量为26.766 kD,理论等电点为6.43;二级结构具有螺旋、β转角、延伸链、无规则卷曲;以4y28.1.O(2.80?)为模板进行同源建模,三级结构是单分子物体,具有6个叶绿体a配体,是一个典型的捕光复合物I叶绿素a/b结合蛋白。系统进化分析表明,与除虫菊和黄花蒿Lhca蛋白进化程度最为接近,同处于菊科这一分支,一致性超过85%,表明该基因家族保守性较强。比对已报道的植物LHC蛋白,发现它们均无信号肽,存在跨膜区,含有结构功能域,为亲水性非分泌蛋白[25,26]。植物LHC表达与抗逆性密切相关。YANG等[27]研究表明,与野生型相比,稀脉浮萍(Lemna perpusilla)的细胞色素b6f突变体在低光强下可维持LHC II在一个恒定的低水平上;李真等[28]研究发现,东南景天(Sedum alfredii)的SaLhcb2在镉、铜、铅处理后,根、茎、叶中表达量相比对照均发生显著上调或者降低;YANG等[29]研究发现,菠菜(Spinacia oleracea)N-端LHCII可识别蛋白降解酶,通过降解LHCII的含量可达到响应不同光强。本研究qPCR结果显示,羊脂酸胁迫处理小飞蓬叶片后,CcLhca-J9的表达量在处理后0—8 h随时间延长表现出下降的趋势。与对照清水和阿魏酸处理相比,羊脂酸抑制CcLhca-J9的表达存在着一定的特异性,预示着LHC可作为抑草靶标用来开发除草剂。
植物LHC是一类潜在的植物源除草剂作用靶标。光合作用抑制剂的开发一直是农药学家创制新型除草剂的热门方向,但是建立在充分研究受体蛋白结构与抑制剂相互作用的前提下[30]。去草净(terburyn)是一种典型三嗪类PSII抑制剂,与D1蛋白氨基酸残基Ile224和Ser233形成氢键,与Glu212、Val220和Ile229也有反应[31]。敌草隆(diuron,DCUM)的芳香取代基与Phe255形成氢键,提高了其抑制活性[32]。TeA是一种从链格孢菌里分离提取出来的抑制PSII的除草化合物,其作用机制也是与靶蛋白D1的相互作用有关[33]。本研究采用分子对接的手段,发现羊脂酸与植物LHC中氨基酸残基Gly68、Phe67、Phe69和Arg197形成氢键和p-π键,其结合能量为-20.9 kJ·mol-1,两者存在较强的作用力,羊脂酸具有开发成光合作用抑制剂类除草剂的潜力。
4 结论
CcLhca-J9是一个典型的捕光复合物I叶绿素a/b结合蛋白,与羊脂酸结合形成氢键和p-π键,同时CcLhca-J9表达量在羊脂酸处理小飞蓬叶片后存在明显的特异性。因而,推测CcLhca-J9在羊脂酸抑制小飞蓬叶片生长过程中起了较为关键的作用,但是该基因是否为植物源羊脂酸的靶标基因有待更进一步的功能验证。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
DOI:10.1017/wet.2019.20URL [本文引用: 1]
DOI:10.1016/S1671-2927(11)60093-XURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1002/ps.2333URL [本文引用: 1]
[本文引用: 1]
DOI:10.1104/pp.16.01009URLPMID:27663408 [本文引用: 1]

State transitions in photosynthesis provide for the dynamic allocation of a mobile fraction of light-harvesting complex II (LHCII) to photosystem II (PSII) in state I and to photosystem I (PSI) in state II. In the state I-to-state II transition, LHCII is phosphorylated by STN7 and associates with PSI to favor absorption cross-section of PSI. Here, we used Arabidopsis (Arabidopsis thaliana) mutants with defects in chlorophyll (Chl) b biosynthesis or in the chloroplast signal recognition particle (cpSRP) machinery to study the flexible formation of PS-LHC supercomplexes. Intriguingly, we found that impaired Chl b biosynthesis in chlorina1-2 (ch1-2) led to preferentially stabilized LHCI rather than LHCII, while the contents of both LHCI and LHCII were equally depressed in the cpSRP43-deficient mutant (chaos). In view of recent findings on the modified state transitions in LHCI-deficient mutants (Benson et al., 2015), the ch1-2 and chaos mutants were used to assess the influence of varying LHCI/LHCII antenna size on state transitions. Under state II conditions, LHCII-PSI supercomplexes were not formed in both ch1-2 and chaos plants. LHCII phosphorylation was drastically reduced in ch1-2, and the inactivation of STN7 correlates with the lack of state transitions. In contrast, phosphorylated LHCII in chaos was observed to be exclusively associated with PSII complexes, indicating a lack of mobile LHCII in chaos Thus, the comparative analysis of ch1-2 and chaos mutants provides new evidence for the flexible organization of LHCs and enhances our understanding of the reversible allocation of LHCII to the two photosystems.
[本文引用: 1]
DOI:10.1007/s00204-016-1721-zURLPMID:27136897 [本文引用: 1]
Phosphinotricin (L-PPT) is the active compound of a broad-spectrum herbicide. Acute poisoning with L-PPT has various clinical manifestations, including seizures and convulsions. However, the exact mechanism of L-PPT toxicity remains unclear. The present study addressed the role of L-PPT, in the excitability of striatal medium-sized spiny neurons (MSNs). In whole-cell current-clamp experiments, L-PPT increased the input resistance (Ri), decreased the rheobase and increased the firing frequency of action potentials. In voltage-clamp experiments, L-PPT inhibited the inward-rectifying potassium (Kir) currents. Finally, the effects of L-PPT mimicked the inhibition of Kir channels with Ba(2+) on neuronal excitability. Altogether, these results suggest that the herbicide L-PPT is a modulator of Kir channels in MSNs. Thereby, Kir channels are potent regulators of the excitability of MSNs and reduced open probability of these channels would generate a powerful upregulation of neuronal output. This effect may represent a possible mechanism for L-PPT dependent neuronal toxicity.
DOI:10.1016/j.arabjc.2010.08.007URL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
DOI:10.1002/pmic.201800294URLPMID:30865362 [本文引用: 1]
Caprylic acid (CAP) is anticipated to be a potential biocontrol herbicide in the control of weeds, however the molecular mechanism of how CAP affects weeds is poorly understood. Here, the physiological and biochemical (protein-level) changes in horseweed (Conyza canadensis L.) are studied under CAP treatment, with infrared gas analyzer and label-free quantitative proteomics methods. In total, 112 differentially-accumulated proteins (DAPs) (>1.5 fold change, p < 0.05) are present between treated horseweed and control samples, with 46 up-regulated and 66 down-regulated proteins. These DAPs are involved in 28 biochemical pathways, including photosynthesis pathways. In particular, six photosynthesis proteins show significant abundance changes in the CAP-treated horseweed. The qRT-PCR results confirm three of the six genes involved in photosynthesis. Moreover, by measuring photosynthesis characteristics, CAP was shown to decrease photosynthetic rate, stomatal conductance, intercellular CO2 concentration, and the transpiration rate of horseweed. These results suggest that photosystem I is one of the main biological processes involved in the response of horseweed to CAP.
URLPMID:7748263 [本文引用: 1]
URL [本文引用: 2]
[本文引用: 2]
DOI:10.1104/pp.17.00505URLPMID:28432258 [本文引用: 2]
The LIL3 protein of Arabidopsis (Arabidopsis thaliana) belongs to the light-harvesting complex (LHC) protein family, which also includes the light-harvesting chlorophyll-binding proteins of photosystems I and II, the early-light-inducible proteins, PsbS involved in nonphotochemical quenching, and the one-helix proteins and their cyanobacterial homologs designated high-light-inducible proteins. Each member of this family is characterized by one or two LHC transmembrane domains (referred to as the LHC motif) to which potential functions such as chlorophyll binding, protein interaction, and integration of interacting partners into the plastid membranes have been attributed. Initially, LIL3 was shown to interact with geranylgeranyl reductase (CHLP), an enzyme of terpene biosynthesis that supplies the hydrocarbon chain for chlorophyll and tocopherol. Here, we show another function of LIL3 for the stability of protochlorophyllide oxidoreductase (POR). Multiple protein-protein interaction analyses suggest the direct physical interaction of LIL3 with POR but not with chlorophyll synthase. Consistently, LIL3-deficient plants exhibit substantial loss of POR as well as CHLP, which is not due to defective transcription of the POR and CHLP genes but to the posttranslational modification of their protein products. Interestingly, in vitro biochemical analyses provide novel evidence that LIL3 shows high binding affinity to protochlorophyllide, the substrate of POR. Taken together, this study suggests a critical role for LIL3 in the organization of later steps in chlorophyll biosynthesis. We suggest that LIL3 associates with POR and CHLP and thus contributes to the supply of the two metabolites, chlorophyllide and phytyl pyrophosphate, required for the final step in chlorophyll a synthesis.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/php.12919URLPMID:29569330 [本文引用: 1]
We present the detailed characterization on the reconstitution of two cyclic tetrapyrrole pigments that have the same substituents but differ in the degree of hydrogenation in the macrocycles from bacteriochlorophyll (BChl) a (7,8,17,18-tetrahydroporphyrin) into the binding sites of B800 BChl a in light-harvesting complex 2 (LH2) of purple photosynthetic bacteria. Both 3-acetyl chlorophyll (Chl) a (17,18-dihydroporphyrin) and 3-acetyl protochlorophyll (PChl) a (porphyrin) were inserted into the B800-binding pockets in LH2, indicating that these pockets allow alteration of the degree of hydrogenation in the cyclic tetrapyrroles. Redshifts of the Qy peak positions of 3-acetyl (P)Chl a by insertion into the B800-binding sites were smaller than that of BChl a. The relative Qy absorbance of 3-acetyl (P)Chl a to B850 BChl a in the reconstituted proteins was significantly smaller than that of B800 BChl a in native LH2 in spite of their high occupancy in the B800-binding sites. These are ascribable to the smaller dipole strengths of 3-acetyl (P)Chl a. We also performed the coreconstitution of both 3-acetyl Chl a and BChl a into the nine B800-binding sites in LH2, indicating that the affinity of 3-acetyl Chl a to the B800-cavity was slightly higher than that of BChl a.
DOI:10.3864/j.issn.0578-1752.2019.19.012URL [本文引用: 1]
【Objective】 In this study, the full-length sequence of gibberellin insensitive gene (GAI2) was obtained from the leaf of Nantongxiaofangshi (Diospyros kaki Linn. cv. nantongxiaofangshi), named as DkGAI2. The subcellular localization and expression characteristics of DkGAI2 gene were analyzed. Then DkGAI2 gene was transformed into plant tobacco, and the morphological and physiological indicators of the transgenic tobacco plants were determined. Our study would provide a theoretical basis for the future research of GAI gene.【Method】 The full-length sequence of DkGAI2 gene was cloned from Nantongxiaofangshi by RT-PCR, 3’ RACE and 5’ RACE using the labeled GAI2 gene as the original sequence of high-throughput transcriptome sequencing of Nantongxiaofangshi (Unpublished). Sequence characteristics were analyzed by bioinformatics. The expression characteristics of DkGAI2 gene in five different phenological stages of Dafangshi and Nantongxiaofangshi, and the expression level of DkGAI2 gene in different tissues of Nantongxiaofangshi was detected by real-time quantitative PCR (qRT-PCR). The transient expression vector pCAMBIA-GFP-1302-DkGAI2 was constructed and was transiently infected with tobacco to analyze the subcellular localization of DkGAI2 gene. DkGAI2 gene driven by the 35S promoter was constructed and delivered into plant tobacco by Agrobacterium-mediated transformation approach. Transgenic plants were identified by using GUS staining and RT-PCR. After transplanting, the plant height, internode length, leaf aspect ratio and the content of GA1 and GA4 in transgenic tobacco plants were measured at the first flowering stage.【Result】 1 827 bp of DkGAI2 was cloned from Nantongxiaofangshi, the nucleotide sequence of DkGAI2 gene shared 72%-80% in homology compared with kiwifruit (KF588651.1), light skin (MF149049.1), pear (KX078214.1), apple (FJ535245.1) and grape (MG738718.1). DkGAI2 gene containing DELLA and GRAS conserved special domain, belonging to DELLA protein gene family. DkGAI2 gene encoding a putative protein about 608 amino acids, the relative molecular mass of DkGAI2 gene was 66.5 KD, the theoretical isoelectric point was 5.54, and the instability coefficient iwas 50.41, without obvious hydrophobic region, without transmembrane domain and signal peptide. Phylogenetic analysis showed that the DkGAI2 gene had close relationship with grape. qRT-PCR result showed that the expression level of DkGAI2 gene in five different phenological stages of Nantongxiaofangshi was higher than that of Dafangshi. DkGAI2 gene showed tissue expression specificity in different tissues of Nantongxiaofangshi, which had the highest expression in old leaves, followed by young leaves and shoot tips, while it had the lowest expression in fruitlet. The green fluorescent signal of the pCAMBIA-GFP- 1302-DkGAI2 fusion protein was located in the nucleus, indicating that DkGAI2 gene localized in the nucleus. After GUS staining and RT-PCR detection, 5 transgenic tobacco lines of DkGAI2 gene were obtained. The GA1 content of transgenic tobacco leaves were increased, the GA4 content were decreased, the total content of GA1 and GA4 were decreased, and the transgenic tobacco plants showed plant dwarfing phenotypes with shortened internodes, reduced leaf aspect ratio, and delayed flowering.【Conclusion】 DkGAI2 gene had tissue expression specificity; DkGAI2 gene localized in the nucleus, the expression level of DkGAI2 gene in Nantongxiaofangshi was higher than that of Dafangshi at the five different phenological periods. It was speculated that DkGAI2 may cause plant dwarfing by reducing GA4 content.
DOI:10.3864/j.issn.0578-1752.2019.19.012URL [本文引用: 1]
【Objective】 In this study, the full-length sequence of gibberellin insensitive gene (GAI2) was obtained from the leaf of Nantongxiaofangshi (Diospyros kaki Linn. cv. nantongxiaofangshi), named as DkGAI2. The subcellular localization and expression characteristics of DkGAI2 gene were analyzed. Then DkGAI2 gene was transformed into plant tobacco, and the morphological and physiological indicators of the transgenic tobacco plants were determined. Our study would provide a theoretical basis for the future research of GAI gene.【Method】 The full-length sequence of DkGAI2 gene was cloned from Nantongxiaofangshi by RT-PCR, 3’ RACE and 5’ RACE using the labeled GAI2 gene as the original sequence of high-throughput transcriptome sequencing of Nantongxiaofangshi (Unpublished). Sequence characteristics were analyzed by bioinformatics. The expression characteristics of DkGAI2 gene in five different phenological stages of Dafangshi and Nantongxiaofangshi, and the expression level of DkGAI2 gene in different tissues of Nantongxiaofangshi was detected by real-time quantitative PCR (qRT-PCR). The transient expression vector pCAMBIA-GFP-1302-DkGAI2 was constructed and was transiently infected with tobacco to analyze the subcellular localization of DkGAI2 gene. DkGAI2 gene driven by the 35S promoter was constructed and delivered into plant tobacco by Agrobacterium-mediated transformation approach. Transgenic plants were identified by using GUS staining and RT-PCR. After transplanting, the plant height, internode length, leaf aspect ratio and the content of GA1 and GA4 in transgenic tobacco plants were measured at the first flowering stage.【Result】 1 827 bp of DkGAI2 was cloned from Nantongxiaofangshi, the nucleotide sequence of DkGAI2 gene shared 72%-80% in homology compared with kiwifruit (KF588651.1), light skin (MF149049.1), pear (KX078214.1), apple (FJ535245.1) and grape (MG738718.1). DkGAI2 gene containing DELLA and GRAS conserved special domain, belonging to DELLA protein gene family. DkGAI2 gene encoding a putative protein about 608 amino acids, the relative molecular mass of DkGAI2 gene was 66.5 KD, the theoretical isoelectric point was 5.54, and the instability coefficient iwas 50.41, without obvious hydrophobic region, without transmembrane domain and signal peptide. Phylogenetic analysis showed that the DkGAI2 gene had close relationship with grape. qRT-PCR result showed that the expression level of DkGAI2 gene in five different phenological stages of Nantongxiaofangshi was higher than that of Dafangshi. DkGAI2 gene showed tissue expression specificity in different tissues of Nantongxiaofangshi, which had the highest expression in old leaves, followed by young leaves and shoot tips, while it had the lowest expression in fruitlet. The green fluorescent signal of the pCAMBIA-GFP- 1302-DkGAI2 fusion protein was located in the nucleus, indicating that DkGAI2 gene localized in the nucleus. After GUS staining and RT-PCR detection, 5 transgenic tobacco lines of DkGAI2 gene were obtained. The GA1 content of transgenic tobacco leaves were increased, the GA4 content were decreased, the total content of GA1 and GA4 were decreased, and the transgenic tobacco plants showed plant dwarfing phenotypes with shortened internodes, reduced leaf aspect ratio, and delayed flowering.【Conclusion】 DkGAI2 gene had tissue expression specificity; DkGAI2 gene localized in the nucleus, the expression level of DkGAI2 gene in Nantongxiaofangshi was higher than that of Dafangshi at the five different phenological periods. It was speculated that DkGAI2 may cause plant dwarfing by reducing GA4 content.
DOI:10.1093/nar/gkw1132URL [本文引用: 1]
DOI:10.1007/s40484-019-0179-4URL [本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2020.01.014URL [本文引用: 1]
【Objective】AtRPL is an important gene regulating Arabidopsis fruit development and mediating replum formation. Using homologous cloning, the RPL homolog in cucumber was identified, the biological function of CsRPL by expression analysis and ectopic transformation in Arabidopsis was explored.【Method】We cloned the CsRPL gene by performing a BLAST search of AtRPL in cucumber genome. Then we performed amino acid sequence alignment of CsRPL and RPL homologs from other species by MEGA5.2. CsRPL expression pattern in cucumber was detected by real-time quantitative PCR (qRT-PCR) and in situ hybridization. Expression and phenotypic analysis of transgenic Arabidopsis upon ectopic expression of CsRPL were performed as well.【Result】Two RPL genes, named CsRPL1 and CsRPL2, were identified in cucumber with the conserved BELL domain and Homeodomain, and two EAR-Motifs. CsRPL1 was expressed in all organs of cucumber, with the highest expression level in male flowers at anthesis. During early stages of fruit development, the CsRPL1 expression decreased gradually. The expression level of CsRPL2 was significantly lower than that of CsRPL1. In situ hybridization revealed that signals of CsRPL1/2 were detected in the placenta of fruits and the central zone (CZ) of the shoot apical meristem (SAM). Ectopic overexpression of CsRPL1/2 into Arabidopsis resulted in shorter siliques, reduced pollen fertility, and inhibited seed development.【Conclusion】CsRPL1/2 were involved in the development of reproductive organs and might have functional redundancy in cucumber. CsRPL1 might play the primary role under normal growth condition. The function of CsRPL1/2 was not fully conserved as compared to that of AtRPL.
DOI:10.3864/j.issn.0578-1752.2020.01.014URL [本文引用: 1]
【Objective】AtRPL is an important gene regulating Arabidopsis fruit development and mediating replum formation. Using homologous cloning, the RPL homolog in cucumber was identified, the biological function of CsRPL by expression analysis and ectopic transformation in Arabidopsis was explored.【Method】We cloned the CsRPL gene by performing a BLAST search of AtRPL in cucumber genome. Then we performed amino acid sequence alignment of CsRPL and RPL homologs from other species by MEGA5.2. CsRPL expression pattern in cucumber was detected by real-time quantitative PCR (qRT-PCR) and in situ hybridization. Expression and phenotypic analysis of transgenic Arabidopsis upon ectopic expression of CsRPL were performed as well.【Result】Two RPL genes, named CsRPL1 and CsRPL2, were identified in cucumber with the conserved BELL domain and Homeodomain, and two EAR-Motifs. CsRPL1 was expressed in all organs of cucumber, with the highest expression level in male flowers at anthesis. During early stages of fruit development, the CsRPL1 expression decreased gradually. The expression level of CsRPL2 was significantly lower than that of CsRPL1. In situ hybridization revealed that signals of CsRPL1/2 were detected in the placenta of fruits and the central zone (CZ) of the shoot apical meristem (SAM). Ectopic overexpression of CsRPL1/2 into Arabidopsis resulted in shorter siliques, reduced pollen fertility, and inhibited seed development.【Conclusion】CsRPL1/2 were involved in the development of reproductive organs and might have functional redundancy in cucumber. CsRPL1 might play the primary role under normal growth condition. The function of CsRPL1/2 was not fully conserved as compared to that of AtRPL.
DOI:10.3724/SP.J.1006.2016.01332URL [本文引用: 1]
The Lhca gene family in green plants encodes several light-harvesting chlorophyll a/b-binding proteins that harvest and transfer light energy to the reaction center of photosystem I (PSI) in photosynthesis. The cDNA sequence of Lhca3 gene was firstly obtained from sugarcane leaf full-length cDNA library through sequencing and validated by homology comparison. It was designated ScLhca3 and submitted to the GenBank (accession number: KU215669). ScLhca3 contains an 804 bp open reading frame (ORF) and encodes a deduced protein of 267 amino acids, with a molecular weight and pI of 28.91 kD and 8.96, respectively. Bioinformatics analysis showed that ScLhca3 is a hydrophilic non-secretory protein with three transmembrane domains and a chlorophyll a/b binding domain. Sequence multi-alignment and phylogenetic analysis demonstrated that the ScLhca3 protein sequence shared a high identity with Lhca3 from other plants, with the specificity of species. The protokaryotic expression vector pGEX-6P-1-ScLhca3 was constructed and expressed in E. coli cells under the induction of IPTG. The subcellular localization experiment showed that the ScLhca3 fused with GFP, a reporter protein, was located on chloroplast. Real-time quantitative PCR analysis showed the expression of ScLhca3 had a clear tissue specificity, and was upregulated by CdCl2, ABA, and H2O2, but downregulated by darkness, NaCl, and PEG.
DOI:10.3724/SP.J.1006.2016.01332URL [本文引用: 1]
The Lhca gene family in green plants encodes several light-harvesting chlorophyll a/b-binding proteins that harvest and transfer light energy to the reaction center of photosystem I (PSI) in photosynthesis. The cDNA sequence of Lhca3 gene was firstly obtained from sugarcane leaf full-length cDNA library through sequencing and validated by homology comparison. It was designated ScLhca3 and submitted to the GenBank (accession number: KU215669). ScLhca3 contains an 804 bp open reading frame (ORF) and encodes a deduced protein of 267 amino acids, with a molecular weight and pI of 28.91 kD and 8.96, respectively. Bioinformatics analysis showed that ScLhca3 is a hydrophilic non-secretory protein with three transmembrane domains and a chlorophyll a/b binding domain. Sequence multi-alignment and phylogenetic analysis demonstrated that the ScLhca3 protein sequence shared a high identity with Lhca3 from other plants, with the specificity of species. The protokaryotic expression vector pGEX-6P-1-ScLhca3 was constructed and expressed in E. coli cells under the induction of IPTG. The subcellular localization experiment showed that the ScLhca3 fused with GFP, a reporter protein, was located on chloroplast. Real-time quantitative PCR analysis showed the expression of ScLhca3 had a clear tissue specificity, and was upregulated by CdCl2, ABA, and H2O2, but downregulated by darkness, NaCl, and PEG.
DOI:10.1007/s10811-006-9116-9URL [本文引用: 1]
Genes encoding proteins of the major light-harvesting complex of photosystem II (LHCII) in higher plants are well studied. However, little is known about the corresponding genes in the green alga Dunaliella salina, although this knowledge might provide valuable information about the respective roles of each LHCII protein at the molecular level under extreme environmental conditions. Here, we describe an additional LhcII gene from D. salina. An LhcII cDNA cloned by screening a D. salina cDNA library contains an open reading frame encoding a protein of 261 amino acids with a calculated molecular mass of 27.8kDa. The deduced amino acid sequence shows high homology with other LHCII proteins. Genomic DNA—obtained by PCR using a specific primer set corresponding to the 5′ and 3′ untranslated regions—was used to determine the intron-exon structure. Short-term changes in mRNA levels after a shift from low-light to high-light or dark conditions were analyzed by real-time quantitative PCR, and indicated that this gene expresses different mRNA levels under different light conditions.
DOI:10.1023/A:1011849919438URL [本文引用: 1]
A cytochrome b 6 f deficient mutant of Lemna perpusilla maintains a constant and lower level of the light-harvesting chl a/b-binding protein complex II (LHC II) as compared to the wild type plants at low-light intensities. Inhibition of the plastoquinone pool reduction increases the LHC II content of the mutant at both low- and high-light intensities but only at high-light intensity in the wild type plants. Proteolytic activity against LHC II appears during high-light photoacclimation of wild type plants. However, the acclimative protease is present in the mutant at both light intensities. These and additional results suggest that the plastoquinone redox state serves as the major signal-transducing component in the photoacclimation process affecting both, synthesis and degradation of LHC II and appearance of acclimative LHC II proteolysis. The plastoquinol pool cannot be oxidized by linear electron flow in the mutant plants which are locked in a ‘high light’ acclimation state. The cytochrome b 6 f complex may be involved indirectly in the regulation of photoacclimation via 1) regulation of the plastoquinone redox state; 2) regulation of the redox-controlled thylakoid protein kinase allowing exposure of the dephosphorylated LHC II to acclimative proteolysis.
DOI:10.11833/j.issn.2095-0756.2014.06.003URL [本文引用: 1]
LHCB基因对植物适应各种环境胁迫的过程中起着重要作用。序列分析显示:东南景天Sedum alfredii的SaLhcb2基因全长929 bp,其中开放阅读框(ORF)为798 bp,含有1个74 bp的内含子。通过蛋白序列比对,SaLhcb2基因编码的蛋白与多种植物的蛋白序列同源性都很高(92%以上)。分析400 μmol·L-1 镉离子(Cd2+),铜离子(Cu2+)和铅离子(Pb2+)胁迫处理后的东南景天,结果显示:镉处理后SaLhcb2基因在茎、叶中表达量快速上升(12.0 h内),根中表达量到48.0 h后才上调。铜处理后0.5 h根中表达显著上调,胁迫6.0 h后茎中表达量显著上调,随后一直降低,叶片中该基因表达量一直较低。铅处理后,根中表达量降低,96.0 h左右比对照略微上调,而茎中96.0 h内表达量相比对照都上调,叶片中96.0 h内都降低。研究结果表明:SaLhcb2与东南景天的镉、铜、铅胁迫抗性有密切的相关性。图6表1参30
DOI:10.11833/j.issn.2095-0756.2014.06.003URL [本文引用: 1]
LHCB基因对植物适应各种环境胁迫的过程中起着重要作用。序列分析显示:东南景天Sedum alfredii的SaLhcb2基因全长929 bp,其中开放阅读框(ORF)为798 bp,含有1个74 bp的内含子。通过蛋白序列比对,SaLhcb2基因编码的蛋白与多种植物的蛋白序列同源性都很高(92%以上)。分析400 μmol·L-1 镉离子(Cd2+),铜离子(Cu2+)和铅离子(Pb2+)胁迫处理后的东南景天,结果显示:镉处理后SaLhcb2基因在茎、叶中表达量快速上升(12.0 h内),根中表达量到48.0 h后才上调。铜处理后0.5 h根中表达显著上调,胁迫6.0 h后茎中表达量显著上调,随后一直降低,叶片中该基因表达量一直较低。铅处理后,根中表达量降低,96.0 h左右比对照略微上调,而茎中96.0 h内表达量相比对照都上调,叶片中96.0 h内都降低。研究结果表明:SaLhcb2与东南景天的镉、铜、铅胁迫抗性有密切的相关性。图6表1参30
DOI:10.1016/S0014-5793(00)01107-8URL [本文引用: 1]
URL [本文引用: 1]
According to the pharmacophore model, a series of new type photosystem II (PS II) inhibitors were designed, synthesized and characterized with element analysis, UV, IR and ~1H NMR spectra. The biological activities of the new PS II inhibitors by Hill reaction showed that they all presented the biological activities of blocking electron transport in photosystem II. The biological activities of some compounds were so high that they could be applied in the future as PS II herbicides.
URL [本文引用: 1]
According to the pharmacophore model, a series of new type photosystem II (PS II) inhibitors were designed, synthesized and characterized with element analysis, UV, IR and ~1H NMR spectra. The biological activities of the new PS II inhibitors by Hill reaction showed that they all presented the biological activities of blocking electron transport in photosystem II. The biological activities of some compounds were so high that they could be applied in the future as PS II herbicides.
URLPMID:16453713 [本文引用: 1]
An X-ray structure analysis of the photosynthetic reaction centre from the purple bacterium Rhodopseudomonas viridis provides structural details of the pigment-binding sites. The photosynthetic pigments are found in rather hydrophobic environments provided by the subunits L and M. In addition to apolar interactions, the bacteriochlorophylls of the primary electron donor (;special pair') and the bacteriopheophytins, but not the accessory bacteriochlorophylls, form hydrogen bonds with amino acid side chains of these protein subunits. The two branches of pigments which originate at the primary electron donor, and which mark possible electron pathways across the photosynthetic membrane, are in different environments and show different hydrogen bonding with the protein: this may help to understand why only one branch of pigments is active in the light-driven electron transfer. The primary electron acceptor, a menaquinone (Q(A)), is in a pocket formed by the M subunit and interacts with it by hydrophobic contacts and hydrogen bonds. Competitive inhibitors of the secondary quinone Q(B) (o-phenanthroline, the herbicide terbutryn) are bound into a pocket provided by the L subunit. Apart from numerous van der Waals interactions they also form hydrogen bonds to the protein.
DOI:10.1007/s00425-002-0886-2URLPMID:12569405 [本文引用: 1]
The relationship between the redox state of the photosystem (PS) I primary donor, P700, and thermal energy dissipation in PSI were examined in intact leaves using simultaneous measurements of absorbance changes at 830 nm and variations of thermal emission monitored by photoacoustic (PA) spectroscopy, respectively. A strict proportionality (close to a 1:1 ratio) was found between the magnitudes of P700 oxidation and a positive variable PA signal induced by far-red light of various irradiances under conditions favoring effective electron donation from PSII to PSI. The proportionality was observed also between the ratio of reduced P700 to the total P700 content and the ratio of the variable component to the total PA signal measured with modulated light of 695 nm. Those findings clearly revealed that in intact leaves, variable thermal dissipation in PSI is determined by the fraction of P700 in the reduced state. Diuron-treated leaves exposed to 45 degrees C in which PSI received electrons not from PSII, but from soluble reductants localized in the chloroplast stroma were also used. In such leaves, the linear relationship between the ratio of reduced P700 to the total P700 content and the ratio of the variable component to the total PA signal measured with modulated light of 700 nm has been found as well, but its slope was twice smaller than in untreated leaves. This is probably related to an increased contribution of thermal emission from inactive PSII to the steady-state level of the PA signal in diuron-treated leaves exposed to high temperatures. The results demonstrated that the yield of variable thermal dissipation is strictly dependent on the redox pressure applied to the photosystem. The above illustrates the strong photochemical energy quenching occurring when the reaction centers are in open state (reduced P700).
DOI:10.1016/j.envexpbot.2014.11.009URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}