 ,1, 李健2, 王峥2, 陈卓1, 何航2, 邓兴旺2, 马力耕,1
,1, 李健2, 王峥2, 陈卓1, 何航2, 邓兴旺2, 马力耕,1Cloning, Expression and Functional Analysis of a Male Fertility Gene ThMs1 in Bread Wheat
HENG YanFang,1, LI Jian2, WANG Zheng2, CHEN Zhuo1, HE Hang2, DENG XingWang2, MA LiGeng,1通讯作者:
责任编辑: 李莉
收稿日期:2020-09-20接受日期:2020-10-30网络出版日期:2020-12-01
| 基金资助: |
Received:2020-09-20Accepted:2020-10-30Online:2020-12-01
作者简介 About authors
衡燕芳,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (7379KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
衡燕芳, 李健, 王峥, 陈卓, 何航, 邓兴旺, 马力耕. 十倍体长穗偃麦草雄性育性基因ThMs1的克隆、表达及功能分析[J]. 中国农业科学, 2020, 53(23): 4727-4737 doi:10.3864/j.issn.0578-1752.2020.23.001
HENG YanFang, LI Jian, WANG Zheng, CHEN Zhuo, HE Hang, DENG XingWang, MA LiGeng.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】小麦是全球范围内广泛种植的农作物,世界上约30%的人口以小麦为主食。随着全球人口的不断增加和耕地面积的不断减少,为了确保未来的全球粮食安全,急需将新一代杂交育种技术(分子设计杂交育种技术)引入小麦育种,利用杂种优势培育产量高、品质优、抗性好的新品种来满足全球人口对粮食的需求[1,2,3,4,5]。小麦是典型的自花授粉作物,实现新一代杂交育种的关键是筛选雄性不育突变体并克隆相关育性调控基因。十倍体长穗偃麦草具有抗病、抗寒、抗旱、耐贫瘠、长穗多花等多种优异性状[6,7,8],是小麦遗传改良中重要的野生近缘种质资源之一,克隆和利用偃麦草中的雄性育性调控基因,可以为远缘杂交育种提供基因资源及理论支撑。【前人研究进展】小麦雄性不育突变体筛选工作始于60年前,目前,人们在普通小麦中发现至少5个核基因突变导致的雄性不育突变体,它们被命名为ms1—ms5[9,10],其中,ms1和ms5是隐性突变体,ms2、ms3和ms4是显性突变体。在水稻中的研究结果表明,克隆导致小麦雄性不育的基因是了解小麦雄性育性调控机制和建立新一代杂交育种体系的关键[4,5]。中国2个团队分别克隆和鉴定了小麦Ms2[11,12],发现普通小麦中Ms2是不表达的,在ms2突变体中,由于启动子区插入一个转座子,导致Ms2在花药中表达,最终引起花粉母细胞和绒毡层降解导致雄性不育。国内马力耕和邓兴旺课题组与国外研究人员同时报道了小麦Ms1的克隆,发现Ms1特异存在于禾本科植物中,在小孢子母细胞特异表达,该基因突变导致雄配子发生障碍[13,14]。最近,Ms5也被克隆[15],其在小麦中有2个等位基因同时不同程度地发挥功能。小麦隐性雄性不育突变体基因的克隆和分子鉴定对了解小麦雄性育性调控机制极为关键,同时这些突变体以及育性基因也为分子设计杂交育种提供了宝贵的小麦基因资源和受体材料。【本研究切入点】小麦核雄性不育突变体ms1稳定的不育特性及其隐性特征使得ms1适合用于小麦分子设计杂交育种实践中,发掘雄性不育突变体ms1的育性恢复基因是实现杂交育种的另一个关键因素。筛选鉴定除Ms1以外的新的育性恢复基因会为分子设计育种提供更多的选择。李振声先生等培育的“蓝粒小麦”携带了4Ag染色体,其着丝点及近着丝点区来自于十倍体长穗偃麦草的E基因组(EeEbEx),而染色体两臂的端部区域来自于St基因组,是一条重组染色体[16]。笔者此前以小麦雄性不育突变体ms1g为母本,以“蓝粒小麦”为父本,从杂交后代中筛选出雄性育性得以恢复的异附加系材料,表明该重组染色体上含有恢复ms1育性的基因。本文以此异附加系为材料,通过同源克隆的方法获得来自十倍体长穗偃麦草、可恢复小麦隐性核雄性不育突变体ms1雄性育性的ThMs1,该工作的结果既可用于小麦杂交育种,又为利用远缘杂交技术发掘近缘种中的有利遗传资源提供了一种模式[17,18,19,20]。【拟解决的关键问题】本研究以小麦隐性核雄性不育突变体ms1与十倍体长穗偃麦草异附加系为材料,克隆调控小麦雄性育性的ThMs1,并进行转基因功能互补验证,进一步的基因表达分析及蛋白功能研究解析ThMs1和Ms1的关系,为该基因在小麦分子设计杂交育种中的应用奠定基础。1 材料与方法
1.1 材料
普通小麦(Triticum aestivum L.,2n=6x=42,AABBDD)、雄性不育突变体小麦ms1g(Triticum aestivum L.,2n=6x=42,AABBDD)、小麦-十倍体长穗偃麦草异附加系、十倍体长穗偃麦草(Thinopyrum ponticum,2n=10x=70,StStEeEbEx)。小麦-十倍体长穗偃麦草异附加系是以小麦隐性核雄性不育突变体ms1g为母本、以李振声先生等培育的“蓝粒小麦”为父本[7,16],在其杂交后代中选育出的稳定雄性可育株系。十倍体长穗偃麦草由中国农业科学院作物科学研究所张学勇研究员提供。1.2 基因组原位杂交
基因组原位杂交流程参考ZHAO等[20]方法。使用Dig Translation Mix(Roche)试剂盒标记十倍体长穗偃麦草基因组DNA探针。基因组原位杂交中探针与封阻DNA的比例为1:300,利用anti-digoxigenin- rhodamine(Roche)试剂盒检测杂交信号。杂交后染色体用含DAPI的抗荧光淬灭封片剂(Vector Laboratories)进行复染。用荧光显微镜(Zeiss Axio Imager M2)拍照保存。1.3 基因克隆、RT-PCR和实时荧光定量PCR
使用TRI试剂(TaKaRa Bio Inc.)提取材料减数分裂期花药的RNA,并用DNaseⅠ(Promega)去除基因组DNA污染。通过First-Strand cDNA Synthesis Kit(Thermo Fisher)合成cDNA。以cDNA为模板,基于小麦及其他物种已知MS1序列设计引物(电子附表1)从小麦-十倍体长穗偃麦草中扩增ThMs1。利用LA Taq(TaKaRa Bio Inc.)进行RT-PCR,并通过2%琼脂糖凝胶电泳检测扩增产物。Primer 5.0软件设计靶向基因的特异性引物,利用SYBR Premix Ex(TaKaRa Bio Inc.),经实时荧光定量PCR检测ThMs1表达特征。用于表达分析的引物见电子附表1。Table S1
附表1
附表1本研究所用引物
Table S1
| 引物 Primer | 序列 Sequence (5'→3') | 目的 Intention |
|---|---|---|
| ThMs1-F | AGATCCCGGCGCCTGCTGCTC | ThMs1扩增引物 PCR primer of ThMs1 |
| ThMs1-R | CGCAGGAGCTGTAGAGCGTGAGGA | |
| BF | TAGCCATCTTTGATCAATGAGC | 4Ag染色体鉴定 Analysis of 4Ag chromosome |
| BR | TGATGAAAGAGCTAGGTGATAGTTG | |
| TF1 | CGCCAGGGTTTTCCCAGTCACGAC | 转基因鉴定 Analysis of transgenic lines |
| TR1 | TACAATGGCTAGTAGAGATTTC | |
| TF2 | ATACGTTTCCTGCTACAGATTTGAGG | |
| TR2 | AAGCAGTGCCGCCAGAGGATCAACGC | |
| TF3 | TTTGCGTTCTGCTGATGATGTG | |
| TR3 | CTCAATTGTCCTTTAGACCATGTCTAAC | |
| TaACTIN-QF | TTCCGTTGCCCTGAGGTCC | RT-PCR和实时荧光定量PCR RT-PCR and real time PCR |
| TaACTIN-QR | TGATCTTCATGCTGCTTGGTGC | |
| Ms1-QF | ACATCATCCTCTGAGTCGCG | |
| Ms1-QR | GACCACGCAAACACGTACG | |
| ThMs1-QF | TCCCGGCGCCTGCTGCTC | |
| ThMs1-QR | CGGCAGAGGCAGGGGACG | |
| Ms1-ISH-F | aagcttGAGCGAGGGAGAGAGAGACC | RNA原位杂交 In situ hybridization |
| Ms1-ISH-F | gaattcATCACATAGCATCAGTGGTTC | |
| ThMs1-ISH-F | aagcttCTAGCGAGCGAGCGAGAGG | |
| ThMs1-ISH-R | gaattcCAACTGGACGGACCATGGC | |
| ThMs1-SB-F1 | TGAGATATACGGAGCGATTTAG | Southern杂交 Southern blot |
| ThMs1-SB-R1 | AGCACGGCAAGCTTTTGCTCTG | |
| His-F | aattcCATCATCATCATCATCACTGActgca | MBP-His载体构建 Vector construction of MBP-His |
| His-R | gTCAGTGATGATGATGATGATGg | |
| Ms1s-His-F | gagggaaggatttcagaattcCAGCCGGGGGCGCCGTGC | MBP-Ms1-His载体构建Vector construction of MBP-Ms1-His |
| Ms1s-His-R | cagtgccaagcttgcctgcagTCAGTGATGATGATGATGATGGGCCGCCTTGGACGGCG | |
| ThMs1s-His-F | gagggaaggatttcagaattcGCGTTCGGGCCGCAGC | MBP-ThMs1-His载体构建 Vector construction of MBP-ThMs1-His |
| ThMs1s-His-R | cagtgccaagcttgcctgcagtcaGTGATGATGATGATGATGGAAGAAGGCCGCCTTGGACG |
新窗口打开|下载CSV
1.4 RNA原位杂交
按照SHITSUKAWA等[21]方法进行RNA原位杂交。将组织在甲醛-乙酸溶液(3.7%甲醛)中于4℃固定8 h。使用引物ThMs1-ISH-F/R和Ms1-ISH-F/R(电子附表1)扩增探针,并将扩增产物分别以正向和反向插入pEASY-T1载体(TransGen Biotech)。通过HindⅢ(反义探针)和EcoRⅠ(有义探针)消化,使载体线性化,将其用作模板以生成具有T7 RNA聚合酶的探针。把材料切成8 μm厚的部分进行原位杂交,并在42℃杂交过夜。利用BCIP/NBT碱性磷酸酶显色试剂盒显色,镜检拍照保存。1.5 Southern blot
使用改良的CTAB方法从幼叶中提取基因组DNA后,参照ZHAO等[20]方法进行杂交。使用核酸分析仪(NanoDrop 2000,Thermo Scientific)对纯化的DNA进行定量。基因组DNA(每个样品40 μg)在37℃用HindⅢ和EcoRⅠ(New England Biolabs,USA)完全消化,然后在4℃和35 V下经0.8%琼脂糖凝胶电泳。将分离的DNA转移到Hybond-N+尼龙膜(GE Healthcare)并通过UV交联固定。电子附表1中列出了用于探针扩增的引物,并用探针标记试剂盒(Roche)标记探针。用ThMs1的特异性探针进行杂交,同时提高杂交温度(66℃)并进行充分洗脱,从而提高杂交信号的特异性,然后使用试剂盒(GE Healthcare,RPN2106)进行检测分析。1.6 转基因功能互补验证
将ThMs1基因组DNA插入经HindⅢ酶切的pAHC20载体中,构建用于功能互补试验的pAHC20- ThMs1p::ThMs1载体(电子附图1-A),包括该基因ATG上游2 271 bp、基因组序列1 878 bp和TGA下游628 bp。通过基因枪法将载体转入ms1g-4Ag单体异附加系(ms1g纯合突变体附加一条长穗偃麦草4Ag染色体)植株的幼胚,4Ag染色体在后代中存在分离,所以需要对转基因植株进行4Ag染色体的鉴定,用于鉴定的引物BF/BR见电子附表1,不含有4Ag染色体的转基因植株即为ms1g纯合突变基因型背景。附图1
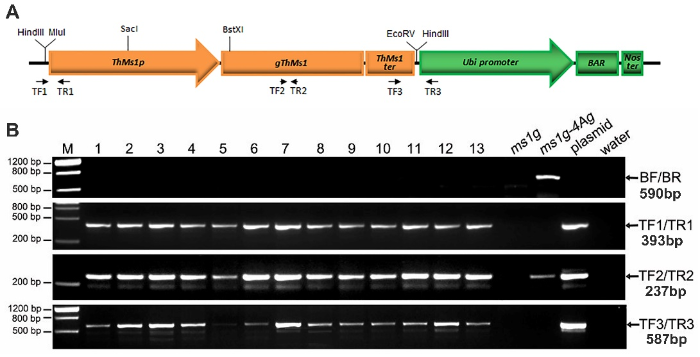 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附图1转基因载体及转基因株系鉴定
A:转基因载体pAHC20-ThMs1p::ThMs1;B:转基因株系的PCR鉴定
Fig. S1The structure of transgenic vector and analysis of transgenic lines
A: Transgenic vector of pAHC20-ThMs1p::ThMs1; B: Analysis of transgenic lines by PCR
在转基因后代鉴定中,同时用TF1/TR1、TF2/TR2和TF3/TR3 3对引物(电子附表1)进行PCR,3对引物均为PCR阳性,说明ThMs1的启动子、基因编码区和终止子都已整合到基因组中,鉴定为转基因阳性植株(电子附图1-B),对检测出的具有ms1g纯合突变且不带有4Ag染色体的转基因阳性植株进行表型鉴定。
1.7 序列比对及系统进化树构建
用NCBI网站上提供的数据信息,获取不同物种中MS1同源性序列,或通过设计引物扩增同源序列。上述序列利用Clustal X 2.1进行多序列比对分析,然后利用MEGA 5.2.1软件通过Neighbour-joining(NJ)方法比对序列构建进化树。1.8 脂质结合活性分析
利用SignalP软件进行蛋白结构预测[22],显示ThMs1蛋白具有8个半胱氨酸残基保守结构的脂结合结构域,将去掉信号肽和跨膜结构域的ThMs1、Ms1与6×His融合构建表达载体,进行原核表达融合蛋白,引物序列见电子附表1。融合蛋白在大肠杆菌BL21菌株中表达并纯化,PIP脂膜P-6001和P-6002购自Echelon Biosciences。参考DOWLER等[23]方法进行蛋白-脂质结合活性检测。2 结果
2.1 十倍体长穗偃麦草ThMs1的克隆
小麦隐性核雄性不育突变体ms1g和普通小麦相比,植株在株型、生育期等农艺性状上无明显差异,但在抽穗扬花期突变体穗部的颖壳呈张开状,而普通小麦的颖壳不张开完成自花授粉(图1-A)。进一步镜检发现,普通小麦的花药发育正常、花粉可育,而雄性不育突变体ms1g与普通小麦相比雌蕊发育正常,早期花药发育也无明显差异,但到花粉成熟时花药稍瘦小不开裂、花粉不育,后期不能正常受精结实(图1-A—图1-D)。通过基因组原位杂交(GISH)证实,在得到的附加系中,附加了十倍体长穗偃麦草4Ag染色体(图1-E),而且附加该染色体后使小麦ms1g雄性育性得到完全恢复(图1-A—图1-D)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1普通小麦、ms1g和ms1g/偃麦草4Ag染色体附加系的表型鉴定及细胞学分析
A:开花期小麦穗,标尺=1 cm;B:花丝伸长之后开裂的花药,标尺=1 mm;C:成熟花粉粒I2-KI染色,标尺=200 μm;D:授粉后15 d的种子,标尺=1 mm,B、C和D图下数字表示统计有此表型的株数及观察的总株数;E:基因组原位杂交分析,红色荧光代表附加染色体;标尺=10 μm;F:花粉DAPI染色,Bar=50 μm
Fig. 1Phenotypic characterization and cytological analysis of bread wheat, ms1g and ms1g/addition line
A: Spikes at flowering, Bar=1 cm; B: Anthers after filament elongation, Bar=1 mm; C: I2-KI staining of mature pollen, Bar=200 μm; D: Seeds at 15 d after pollination, Bar=1 mm, the numbers below the figure in B, C and D indicate the number of plants with the phenotypes and the number of all observed plants; E: Genomic in situ hybridization (GISH) of somatic metaphase chromosomes, Bar=10 μm; F: Microspores from DAPI-stained, Bar=50 μm
花药的组织切片观察显示,在小麦ms1突变体中,花药不同发育时期的药室壁层结构与普通小麦相比无明显差异,绒毡层细胞的发生和降解过程与普通小麦相比也无明显差异,但ms1突变体小孢子发育不正常(电子附图2)。在整个花药及花粉发育过程中,小麦ms1g-4Ag异附加系与普通小麦相比无显著差异(电子附图2),表明Ms1突变没有影响小麦孢子体组织的发育,但影响了小孢子的发育。对小孢子细胞核染色分析,发现ms1g突变体在小孢子发育的单核早期之前与普通小麦相比无明显异常,到单核晚期出现约28%的花粉皱缩现象,而且细胞核也呈凝聚状,到双核早期表型加剧,在突变体中处于双核晚期及三核期的花粉发育完全异常、花粉严重皱缩、细胞核降解,不能产生可育的三核成熟花粉(图1-F);小麦ms1g-4Ag异附加系中花药和花粉发育与普通小麦相比无明显差异(图1-F),表明长穗偃麦草的附加系中含有小麦ms1育性恢复基因。
附图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附图2普通小麦、ms1g突变体、ms1g/附加系中不同 时期花药组织石蜡切片观察
E:表皮层;Ar:孢原细胞;Sp:造孢细胞;En:内皮层;ML:中间层;T:绒毡层;MMC:小孢子母细胞;MC:减数分裂细胞;Tds:四分体;BP:双核期花粉;MP:三核期花粉。标尺=50 μm
Fig. S2Paraffin sections of anthers from bread wheat, ms1g, ms1g/addition line at different stages
E: Epidermis; Ar: Archesporial cell; Sp: Sporogenous cell; En: Endothecium; ML: Middle layer; T: Tapetum; MMC: Microspore mother cell; MC: Meiotic cell; Tds: Tetrads; BP: Bicellular pollen; MP: Mature pollen. Bar=50 μm
依据小麦中Ms1的序列及禾本科其他物种中的相似序列比对结果,设计扩增引物,从小麦ms1g-十倍体长穗偃麦草异附加系中,克隆了来自十倍体长穗偃麦草的雄性育性恢复基因ThMs1。ThMs1与小麦4A、4B、4D染色体上3个同源基因Ms1-A、Ms1和Ms1-D(Ms1来自小麦B基因组)的基因组序列进行比对分析,显示ThMs1与小麦的这三个同源基因在基因组序列上相似性分别为88.24%、82.91%和88.94%(电子附图3)。
附图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附图3ThMs1及小麦同源基因序列比对分析
Fig. S3Sequence alignment of ThMs1 and its orthologues in bread wheat
为了进一步检测小麦ms1g-4Ag异附加系和十倍体长穗偃麦草中ThMs1的拷贝数量,设计了ThMs1特异的探针(序列见附图3红框中所示),通过Southern blot在小麦ms1g-4Ag附加系和十倍体长穗偃麦草中分别检测到一条ThMs1的特异杂交信号(图2);而在普通小麦和ms1g雄性不育突变体中都没有检测到特异的杂交信号,表明ms1g-4Ag附加系中含有一个拷贝ThMs1,十倍体长穗偃麦草也含有一个拷贝ThMs1(但可能还含有ThMs1的同源基因)
图2
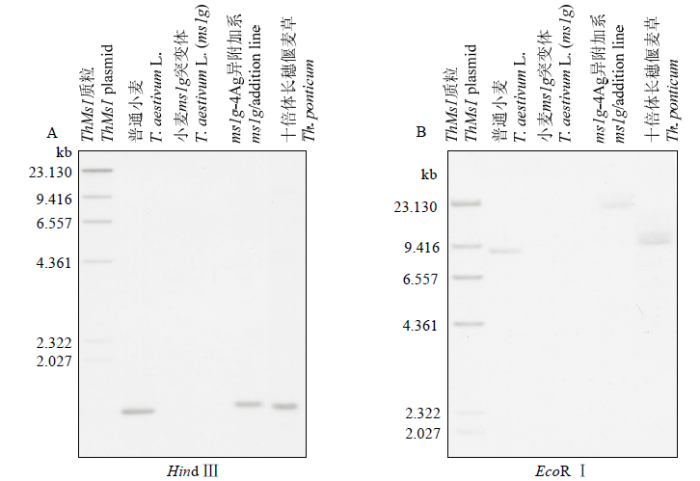 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2ThMs1特异探针与普通小麦、ms1g突变体、ms1g-4Ag附加系、十倍体长穗偃麦草基因组DNA的Southern blot分析
基因组DNA分别用HindⅢ(A)和EcoRⅠ(B)酶切处理
Fig. 2Southern blot of ThMs1 in genomic DNA from bread wheat, ms1g, ms1g-4Ag addition line and Th. Ponticum
Genomic DNA was digested with HindⅢ(A) and EcoRⅠ(B), respectively
2.2 ThMs1转基因功能验证
序列对比结果表明,ThMs1是小麦Ms1的同源基因(电子附图3)。为验证ThMs1的功能,构建用于功能互补试验的pAHC20-ThMs1p::ThMs1载体(电子附图1-A),并将其转入ms1g-4Ag单体异附加系植株的幼胚所制备的愈伤组织中,独立进行2批转化。通过在转化载体上设计3对PCR引物鉴定转基因完整性,获得13株背景为ms1g纯合基因型的T0代转基因阳性植株,后期观察其花粉育性。可育的T0代转基因阳性植株自交结实得到的T1代株系继续PCR鉴定,获得5个独立的、背景为ms1g纯合,且不带有4Ag染色体、转基因可以稳定遗传的株系。上述转基因株系进行穗型、花药形态和花粉育性等观察,结果表明,在小麦ms1g中转入ThMs1能够使ms1g突变体的花粉发育恢复正常,并正常结实(图3),表明ThMs1是小麦Ms1的同源基因,而且在小麦体内能够完全替代小麦Ms1的功能;同时也说明对小麦ms1g的育性表型而言,虽然小麦ms1g-4Ag异附加系转入了一条染色体,但发挥恢复育性功能的只是ThMs1一个基因。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3ThMs1转化小麦ms1的转基因功能互补分析
普通小麦、小麦雄性不育突变体ms1g、背景为ms1g纯合的T1代ThMs1转基因阳性株系。A:开花期小麦穗,标尺=1 cm;B:花丝伸长之后的花药,标尺=1 mm;C:开花后15 d的种子,标尺=1 mm;D:成熟花粉粒I2-KI染色,标尺=200 μm,B、C和D图下数字表示统计有此表型的株数及观察的总株数
Fig. 3Complementation of ms1g by transformation with ThMs1
Ms1, ms1g, T1 transgenic line containing ThMs1 in a ms1g background. A: Spikes at flowering, Bar=1 cm; B: Anthers before filament elongation, Bar=1 mm; C: Seeds at 15 d after pollination, Bar=1 mm; D: I2-KI staining of mature pollen grains, Bar=200 μm, the numbers below the figure in B, C and D indicate the number of plants with the phenotype and the number of observed plants
2.3 ThMs1的表达检测及蛋白聚类分析
基因的表达模式对其发挥功能非常关键,通过以下3种方式检测ThMs1在小麦ms1g-4Ag附加系中的表达模式并与小麦Ms1的表达模式比较:RT-PCR、qRT-PCR和RNA原位杂交。RT-PCR和qRT-PCR表达分析显示ThMs1和Ms1在小麦根、茎、叶中无表达,在内外稃和雌蕊中也无表达;在发育的穗中有表达,尤其在减数分裂期的花药中表达量较高,在单核期的花药中有表达但表达量较低(图4-A—图4-D)。另外,小麦基因组中还含有Ms1的2个同源基因Ms1-A和Ms1-D,这两个基因在所检测的所有组织中均没有表达,表明这两个Ms1同源基因的表达可能被沉默了(图4-A和图4-B)。上述关于Ms1以及它的2个同源基因Ms1-A和Ms1-D表达模式与之前的报道一致[13],同时也表明十倍体长穗偃麦的ThMs1在4Ag附加系中与小麦中的Ms1在小麦中具有非常相似的表达模式。图4
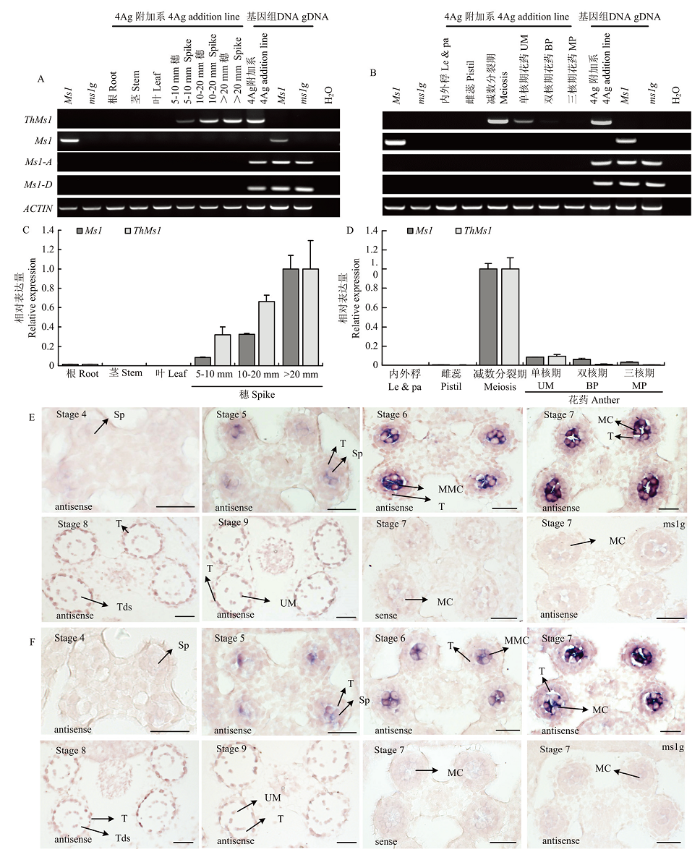 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4Ms1和ThMs1分别在普通小麦以及ms1g-4Ag附加系中的表达检测
A、B:半定量PCR;C、D:实时荧光定量PCR;E、F:RNA原位杂交分析。Ms1在普通小麦中检测,ThMs1在4Ag附加系中检测。Sp:造孢细胞;T:绒毡层;MMC:小孢子母细胞;MC:减数分裂细胞;Tds:四分体;Le&pa:内外稃;UM:单核期花粉;BP:双核期花粉;MP:三核期花粉;标尺=50 μm
Fig. 4Expression analysis of ThMs1 and Ms1 in ms1g-4Ag addition line and bread wheat, respectively
A, B: Expression analysis by RT-PCR; C, D: quantitative RT-PCR; E, F: RNA in situ hybridization analysis. Ms1 in bread wheat and ThMs1in 4Ag addition line anthers. Sp: Sporogenous cell; T: Tapetum; MMC: Microspore mother cell; MC: Meiotic cell; Tds: Tetrads; Le&pa: Lemma and palea; UM: Unicellular microspore; BP: Bicellular pollen; MP: Mature pollen; Bar = 50 μm
进一步通过RNA原位杂交发现ThMs1和Ms1在小孢子母细胞特异表达,两者具有相似的表达模式,它们在花药发育的第6期和第7期小孢子母细胞发育时表达量比较高,到花药发育的第8期四分体时期表达量明显下调(图4-E和图4-F),该表达模式与小麦Ms1的功能以及ThMs1在小麦ms1g-4Ag附加系中的功能一致,同时也很好地解释了为何ThMs1在小麦ms1g-4Ag附加系中可以替代小麦Ms1的功能(二者有非常相似的表达模式)。
Western blot检测发现Ms1和ThMs1蛋白分别在普通小麦和4Ag附加系的减数分裂期花药中表达,而雄性不育突变体ms1g中没有Ms1表达(电子附图4)。上述结果一方面说明制备的小麦Ms1抗体能够特异识别Ms1及其同源蛋白,同时也表明ThMs1和Ms1蛋白也有相似的表达模式。
附图4
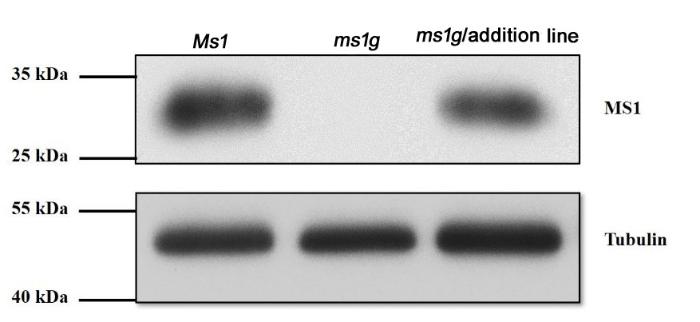 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附图4Ms1和ThMs1分别在普通小麦、4Ag附加系的减数分裂期花药中表达检测
Fig. S4Analysis of expression of Ms1 and ThMs1 in developing anther during microspore meiosis from bread wheat and ms1g-4Ag addition line
为了解小麦Ms1和偃麦草ThMs1的演化关系,将不同物种中MS1同源序列进行比对(电子附图5)。通过聚类分析发现,MS1只存在于禾本科植物中(电子附图6)。十倍体长穗偃麦草的ThMs1蛋白与普通小麦、四倍体小麦、二倍体小麦祖先种中的MS1蛋白亲缘关系较近,属于同一分支(电子附图6)。上述结果表明MS1是禾本科植物进化过程形成的一个新基因,同时也说明禾本科植物雄性育性调控可能与双子叶植物如拟南芥以及禾本科植物以外的单子叶植物有不同之处。
附图5
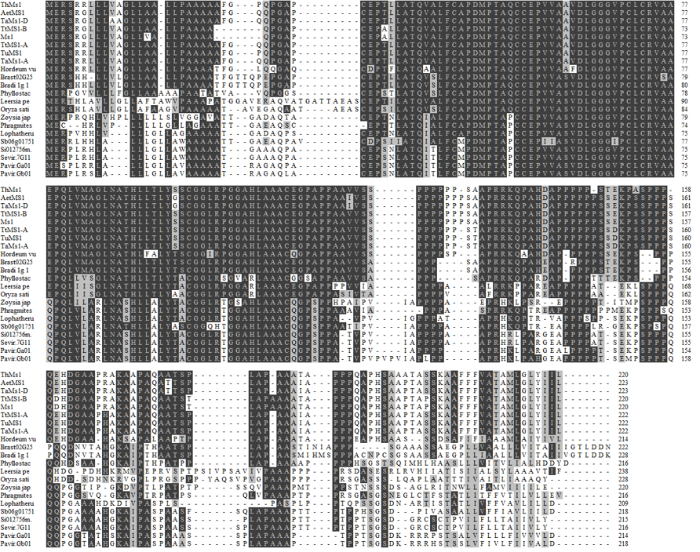 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附图5ThMs1及其同源蛋白序列的比对分析
Fig. S5Sequence alignment of ThMs1 and its orthologues in Poaceae plants
Th:十倍体长穗偃麦草Thinopyrum ponticum;Aet:节节麦Aegilops tauschii;Ta:普通小麦Triticum aestivum;Tt:圆锥小麦Triticum turgidum;Tu:乌拉尔图小麦Triticum Urartu;Bradi:二穗短柄草Brachypodium distachyon;Brast:短柄藓Brachypodium stacei;Hordeum vulgare:大麦; Phyuostachys heterocycla:毛竹;Leersia perrieri:假稻属;Oryza sativa:水稻;Zoysia japonica:结缕草;Phragmites australis:芦苇;Lophtherum gracile:淡竹叶;Sb:高粱Sorghum bicolor;Si:谷子Setaria italica;Sevir:狗尾草Setaria viridis;Pavir:柳枝稷Panicum virgatum。下同The same as below
附图6
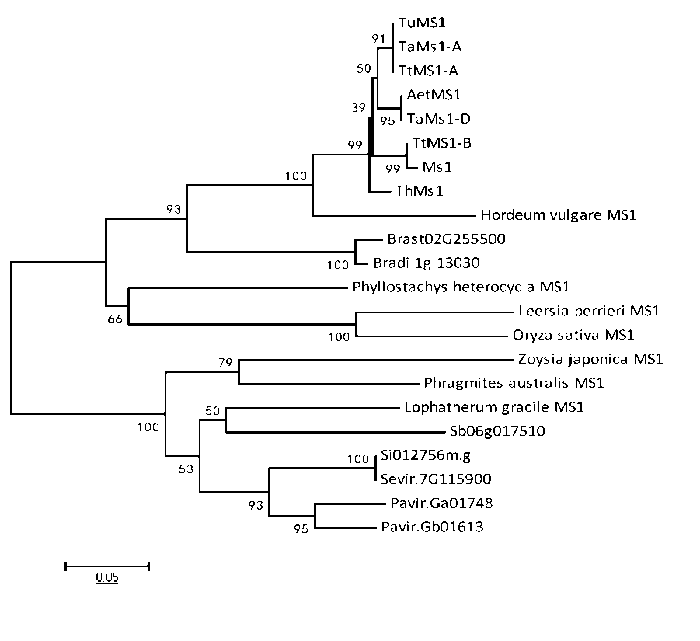 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附图6ThMs1及其同源蛋白的聚类分析
聚类分析的序列同
Fig. S6Phylogenetic tree of ThMs1 and its orthologs in Poaceae plants
Sequences were used to produce the phylogenetic tree shown in Fig. S4
2.4 ThMs1蛋白具有脂结合活性
ThMs1蛋白结构预测显示中间有一个具有8个半胱氨酸残基保守结构的脂结合结构域(电子附图7)。鉴于ThMs1与Ms1同样具有预测的脂结合(lipid transfer protein,LTP)结构域,试图鉴定ThMs1是否具有脂分子结合活性,并以Ms1作对照。为此,首先在大肠杆菌中表达Ms1和ThMs1蛋白,蛋白可溶性检测发现经过IPTG诱导后能够在细菌中表达MBP-His、MBP-Ms1-His和MBP-ThMs1-His蛋白,进一步纯化了这三种融合蛋白(图5)。利用纯化的MBP-His、MBP-Ms1-His、MBP-ThMs1-His蛋白的脂结合试验表明,与Ms1类似,ThMs1蛋白可以结合磷脂酸(phosphatidic acid,PA)和磷酸化的磷脂酰肌醇(phosphatidylinositols,PIs),包括:磷脂酰肌醇-3-磷酸(PI(3)P)、磷脂酰肌醇-4-磷酸(PI(4)P)、磷脂酰肌醇-5-磷酸(PI(5)P)、磷脂酰肌醇-3,4-二磷酸(PI(3,4)P2)、磷脂酰肌醇-3,5-二磷酸(PI(3,5)P2)、磷脂酰肌醇-4,5-二磷酸(PI(4,5)P2)和磷脂酰肌醇-3,4,5-三磷酸(PI(3,4,5)P3),但不结合测试的其他脂类分子(图5),表明Ms1和ThMs1蛋白均特异结合磷脂酸和磷酸化的磷脂酰肌醇。附图7
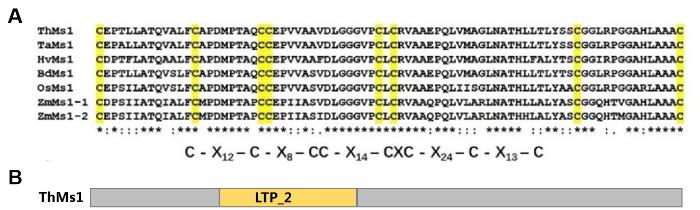 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附图7ThMs1蛋白结构预测
A:8个半胱氨酸残基保守结构;B:预测的脂结合序列
Fig. S7The predicted structures of ThMs1
A: The conserved domain with 8 Cysteine residues; B: Predicted lipid binding domain
图5
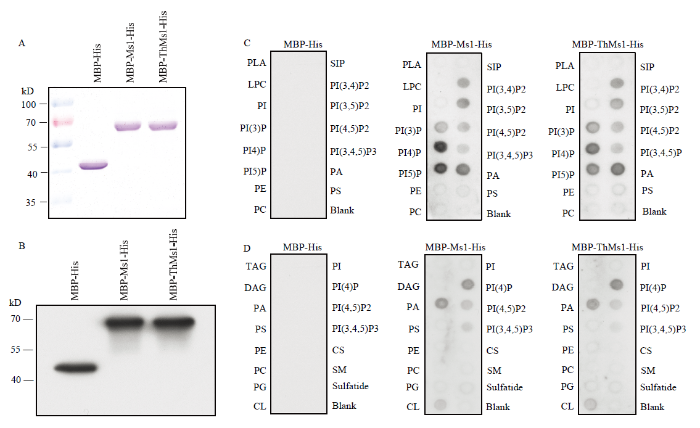 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5Ms1和ThMs1蛋白体外结合磷脂类分子
A、B:MBP-His、MBP-Ms1-His、MBP-ThMs1-His蛋白的纯化和检测;MBP-His、MBP-Ms1-His、MBP-ThMs1-His蛋白覆膜:脂膜P-6001(C)和脂膜P-6002(D)。CL:心磷脂;CS:胆固醇;DAG:二酰甘油;LPA:溶血磷脂酸;LPC:溶血磷脂酰胆碱;PE:磷脂酰乙醇胺;PG:磷脂酰甘油;PI:磷脂酰肌醇;PI(3)P:磷脂酰肌醇-3-磷酸;PI(4)P:磷脂酰肌醇-4-磷酸;PI(5)P:磷脂酰肌醇-5-磷酸;PI(3,4)P2:磷脂酰肌醇-3,4-二磷酸;PI(3,5)P2:磷脂酰肌醇-3,5-二磷酸酯;PI(4,5)P2:磷脂酰肌醇-4,5-二磷酸;PI(3,4,5)P3:磷脂酰肌醇-3,4,5-三磷酸;PS:磷脂酰丝氨酸;SM:鞘磷脂;S1P:鞘氨醇-1-磷酸;TAG:三酰甘油
Fig. 5Both Ms1 and ThMs1 bind phospholipids in vitro
A, B: Purified MBP-His, MBP-Ms1-His and MBP-ThMs1-His; MBP-His, MBP-Ms1-His and MBP-ThMs1-His were overlaid in an Echelon P-6001 lipid strip (C) and Echelon P-6002 membrane strip (D). CL: Cardiolipin. Cs: Cholesterol. DAG: Diacylglycerol. LPA: Lysophosphatidic acid. LPC: Lysophos-phocholine; PC: Phosphatidylcholine; PE: Phosphatidylethanolamine; PG: Phosphatidylglycerol; PI: Phosphatidy-linositol; PI(3)P: Phosphatidylinositol-3-phosphate; PI(4)P: Phosphatidylinositol-4-phosphate; PI(5)P: Phosphatidylinositol-5-phosphate; PI(3,4)P2: Phosphatidylinositol-3,4-bisphosphate; PI(3,5)P2: Phospha- tidylinositol-3,5-bisphosphate; PI(4,5)P2: Phosphatidylinositol-4,5-bisphosphate; PI(3,4,5)P3: Phospha- tidylinositol-3,4,5-triphosphate; PS: Phosphatidylserine; SM: Sphingomyelin; S1P: Sphingosine-1 -phosphate; TAG: Triacylglycerol
3 讨论
十倍体长穗偃麦草是小麦的野生近缘种,具有抗病、抗旱、长穗多花等多种优良性状,对小麦的遗传改良具有重要作用[6,7,8]。本研究发现来自十倍体长穗偃麦草基因组中的小麦雄性育性基因Ms1的同源基因ThMs1具有与小麦Ms1相似的基因结构(电子附图3和电子附图7),而且ThMs1在小麦中与小麦Ms1具有相似的组织细胞特异性表达模式(图4),ThMs1和Ms1蛋白都具有脂结合活性(图6),最终导致这两个基因也具有相似的生物学功能,ThMs1可以替代Ms1的功能恢复小麦雄性不育突变体ms1的花粉育性缺陷(图1和图2)。利用杂种优势制备杂交种是作物遗传育种的重要方向之一,玉米等主要农作物已经在杂交育种方面取得了成功并产生巨大的经济效益,目前种植的玉米品种主要是通过杂交育种制备的品种[24,25]。FREEMAN[26]首次报道了小麦的杂种优势,之后研究人员也证实杂交小麦的生产潜力[27,28],但由于小麦是自花授粉作物,无法实现通过人工去雄进行大规模杂交制种,所以利用雄性不育进行小麦杂交制种是实现小麦杂种优势利用的最佳选择[29,30]。过去几十年建立的小麦杂交育种体系各有缺陷:以细胞质雄性不育为基础的第一代“三系法”系统,受恢保关系的制约导致资源利用率低[31,32];以环境敏感型雄性不育为基础的“两系法”系统受光温条件影响较大、无法满足杂交种纯度要求[28, 33-34],导致目前全球杂交小麦种植面积比例很低;化学杀雄法制备杂交种,存在杀雄专一性不够、成本高、易污染环境等缺点[28,35]。分子设计杂交育种技术具有不育性稳定、控制不育性状的基因遗传行为简单易于开展优良性状的聚合育种等优点[4-5,36]。目前,利用此方法已成功实现了杂交水稻育种技术的新突破[4,5]。分子设计杂交育种技术也应该适用于小麦杂交育种实践,而实现小麦分子设计杂交育种技术的关键是筛选稳定的小麦雄性不育突变体及对应的育性恢复基因。本研究获得了一个新的小麦雄性育性恢复基因ThMs1,该基因在ms1g突变体中能够完全替代小麦Ms1的功能恢复雄性育性(图1和图2)。因此,ThMs1可以替代小麦Ms1作为育性恢复基因,应用于利用小麦隐性核雄性不育突变体ms1制备小麦新一代杂交育种体系中。同时,由于十倍体长穗偃麦草以及该附加系中含有植物育性以及一些抗性基因,该附加系可以应用于小麦杂交育种体系中。为此,本文也提供了把十倍体长穗偃麦草控制重要农艺性状基因用于小麦育种的一种模式,即可以通过杂交把十倍体长穗偃麦草的某一条染色体导入小麦细胞或把某个控制重要农艺性状的基因转入到小麦基因组中。
4 结论
克隆了十倍体长穗偃麦草4Ag染色体上携带的ThMs1,偃麦草中的ThMs1在小麦体内与小麦Ms1具有相似的时空表达模式,2个蛋白也具有相似的生化活性和生物学功能。(责任编辑 李莉)
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1038/nature01014URLPMID:12167873 [本文引用: 1]

A doubling in global food demand projected for the next 50 years poses huge challenges for the sustainability both of food production and of terrestrial and aquatic ecosystems and the services they provide to society. Agriculturalists are the principal managers of global usable lands and will shape, perhaps irreversibly, the surface of the Earth in the coming decades. New incentives and policies for ensuring the sustainability of agriculture and ecosystem services will be crucial if we are to meet the demands of improving yields without compromising environmental integrity or public health.
DOI:10.1038/nature10452URLPMID:21993620 [本文引用: 1]
Increasing population and consumption are placing unprecedented demands on agriculture and natural resources. Today, approximately a billion people are chronically malnourished while our agricultural systems are concurrently degrading land, water, biodiversity and climate on a global scale. To meet the world's future food security and sustainability needs, food production must grow substantially while, at the same time, agriculture's environmental footprint must shrink dramatically. Here we analyse solutions to this dilemma, showing that tremendous progress could be made by halting agricultural expansion, closing 'yield gaps' on underperforming lands, increasing cropping efficiency, shifting diets and reducing waste. Together, these strategies could double food production while greatly reducing the environmental impacts of agriculture.
DOI:10.1126/science.1251788URLPMID:25035500 [本文引用: 1]
An ordered draft sequence of the 17-gigabase hexaploid bread wheat (Triticum aestivum) genome has been produced by sequencing isolated chromosome arms. We have annotated 124,201 gene loci distributed nearly evenly across the homeologous chromosomes and subgenomes. Comparative gene analysis of wheat subgenomes and extant diploid and tetraploid wheat relatives showed that high sequence similarity and structural conservation are retained, with limited gene loss, after polyploidization. However, across the genomes there was evidence of dynamic gene gain, loss, and duplication since the divergence of the wheat lineages. A high degree of transcriptional autonomy and no global dominance was found for the subgenomes. These insights into the genome biology of a polyploid crop provide a springboard for faster gene isolation, rapid genetic marker development, and precise breeding to meet the needs of increasing food demand worldwide.
DOI:10.1073/pnas.1613792113URL [本文引用: 4]
DOI:10.1360/052013-299URL [本文引用: 4]
DOI:10.1360/052013-299URL [本文引用: 4]
DOI:10.1007/s00122-017-2939-8URLPMID:28656363 [本文引用: 2]
KEY MESSAGE: A major locus for resistance to different Fusarium diseases was mapped to the most distal end of Th. elongatum 7EL and pyramided with Th. ponticum beneficial genes onto wheat 7DL. Perennial Triticeae species of the Thinopyrum genus are among the richest sources of valuable genes/QTL for wheat improvement. One notable and yet unexploited attribute is the exceptionally effective resistance to a major wheat disease worldwide, Fusarium head blight, associated with the long arm of Thinopyrum elongatum chromosome 7E (7EL). We targeted the transfer of the temporarily designated Fhb-7EL locus into bread wheat, pyramiding it with a Th. ponticum 7el1L segment stably inserted into the 7DL arm of wheat line T4. Desirable genes/QTL mapped along the T4 7el1L segment determine resistance to wheat rusts (Lr19, Sr25) and enhancement of yield-related traits. Mapping of the Fhb-7EL QTL, prerequisite for successful pyramiding, was established here on the basis of a bioassay with Fusarium graminearum of different 7EL-7el1L bread wheat recombinant lines. These were obtained without resorting to any genetic pairing promotion, but relying on the close 7EL-7el1L homoeology, resulting in 20% pairing frequency between the two arms. Fhb-7EL resided in the telomeric portion and resistant recombinants could be isolated with useful combinations of more proximally located 7el1L genes/QTL. The transferred Fhb-7EL locus was shown to reduce disease severity and fungal biomass in grains of infected recombinants by over 95%. The same Fhb-7EL was, for the first time, proved to be effective also against F. culmorum and F. pseudograminearum, predominant agents of crown rot. Prebreeding lines possessing a suitable 7EL-7el1L gene/QTL assembly showed very promising yield performance in preliminary field tests.
DOI:10.1007/BF00226688URLPMID:24202744 [本文引用: 3]
By crossing fertile nullisomics-4D of common wheat (nullisomics 72180 and 'Tianxuan' no. 15) with three octoploid Agrotriticum lines [partial amphiploids (2n=56) 78829 from the offsprings of wheat with Agropyron intermedium (=Thinopyrum intermedium, 2n=42) and 784 and 7631 from those of wheat with Ag. elongatum (Thinopyrum elongatum, 2n=70)], backcrossing the hybrid with 4D nullisomics as recurrent parent for one to two generations, and then selfing the BC individuals, we obtained three different alien substitution lines, 4Ei (4D), 4Ee(4D) blue grain, and 4Fe(4D) hairy leaf. The F1 hybrids of 784x78829 had the basic chromosome pairing configuration of 27'' + 2', indicating that 784 and 78829 differed by only one pair of chromosomes. This means that Ag. intermedium and Ag. elongatum have a genome (or genomes) in common. The formation of 22'' +12' in the F1 of 784x7631 suggested that there was only one pair of homologous chromosomes of Ag. elongatum in 784 and 7631. Because F1 plants of 7631x78829 generally formed 21'' +14' at metaphase I, the Agropyron genomes in the two octoploid Agrotriticums were presumed to differ from each other. Upon intercrossing the three alien substitution lines, we found that all of the pollen mother cells of the F1 plants had 20'' + 2' or more univalents at metaphase I. We concluded that the three Agropyron chromosomes were not homologous but belonged to the same homoeologous group. These results were confirmed by reciprocal crosses of the three octoploids. The relative transmission rates of 4Ee, 4Fe, 4Ei, 4R and 4D were calculated using blue grain and hairy leaf as genetic markers; these were 4D>4Ee>4Ei, 4Fe>4R. The four alien chromosomes could compensate for the 4D of bread wheat, the substitution lines being vigorous and fertile.
DOI:10.1007/s11032-019-1053-9URL [本文引用: 2]
DOI:10.2135/cropsci2002.1447URL [本文引用: 1]
//
[本文引用: 1]
DOI:10.1038/ncomms15121URLPMID:28452349 [本文引用: 1]
Male sterility is a valuable trait for plant breeding and hybrid seed production. The dominant male-sterile gene Ms2 in common wheat has facilitated the release of hundreds of breeding lines and cultivars in China. Here, we describe the map-based cloning of the Ms2 gene and show that Ms2 confers male sterility in wheat, barley and Brachypodium. MS2 appears as an orphan gene within the Triticinae and expression of Ms2 in anthers is associated with insertion of a retroelement into the promoter. The cloning of Ms2 has substantial potential to assemble practical pipelines for recurrent selection and hybrid seed production in wheat.
DOI:10.1038/ncomms15407URLPMID:28497807 [本文引用: 1]
The male-sterile ms2 mutant has been known for 40 years and has become extremely important in the commercial production of wheat. However, the gene responsible for this phenotype has remained unknown. Here we report the map-based cloning of the Ms2 gene. The Ms2 locus is remarkable in several ways that have implications in basic biology. Beyond having no functional annotation, barely detectable transcription in fertile wild-type wheat plants, and accumulated destructive mutations in Ms2 orthologs, the Ms2 allele in the ms2 mutant has acquired a terminal-repeat retrotransposon in miniature (TRIM) element in its promoter. This TRIM element is responsible for the anther-specific Ms2 activation that confers male sterility. The identification of Ms2 not only unravels the genetic basis of a historically important breeding trait, but also shows an example of how a TRIM element insertion near a gene can contribute to genetic novelty and phenotypic plasticity.
DOI:10.1073/pnas.1715570114URL [本文引用: 2]
DOI:10.1038/s41467-017-00945-2URL [本文引用: 1]
DOI:10.1111/tpj.14350URLPMID:31009129 [本文引用: 1]
Nuclear male-sterile mutants with non-conditional, recessive and strictly monogenic inheritance are useful for both hybrid and conventional breeding systems, and have long been a research focus for many crops. In allohexaploid wheat, however, genic redundancy results in rarity of such mutants, with the ethyl methanesulfonate-induced mutant ms5 among the few reported to date. Here, we identify TaMs5 as a glycosylphosphatidylinositol-anchored lipid transfer protein required for normal pollen exine development, and by transgenic complementation demonstrate that TaMs5-A restores fertility to ms5. We show ms5 locates to a centromere-proximal interval and has a sterility inheritance pattern modulated by TaMs5-D but not TaMs5-B. We describe two allelic forms of TaMs5-D, one of which is non-functional and confers mono-factorial inheritance of sterility. The second form is functional but shows incomplete dominance. Consistent with reduced functionality, transcript abundance in developing anthers was found to be lower for TaMs5-D than TaMs5-A. At the 3B homoeolocus, we found only non-functional alleles among 178 diverse hexaploid and tetraploid wheats that include landraces and Triticum dicoccoides. Apparent ubiquity of non-functional TaMs5-B alleles suggests loss-of-function arose early in wheat evolution and, therefore, at most knockout of two homoeoloci is required for sterility. This work provides genetic information, resources and tools required for successful implementation of ms5 sterility in breeding systems for bread and durum wheats.
[本文引用: 2]
DOI:10.3864/j.issn.0578-1752.2020.07.001URL [本文引用: 1]
Wheat alien species are vast reservoir of diversity for disease and pest resistance as well as stress tolerance, which are excellent gene sources for wheat breeding. Through wide hybridization, the genes of alien species could be transferred to wheat to create wheat-alien chromosome lines such as amphiploids or partial amphiploids, additions, substitutions and translocation lines. These genetic stocks could be utilized to study chromosome behavior and genome evolution, mapping genes, and diversifying the genetic basis of wheat for diseases and pest resistance, as well as yield and quality improvement. In order to understand the progress of wheat wide hybridization and useful gene transfer from alien species to wheat, in this paper, the classification of the tribe Triticeae, the definition and significance of wheat wide hybridization, alien transfers progress from species belonging to genera Aegilops, Secale, Thinopyrum, Dasypyrum, Agropyron, Hordeum, Elymus, Leymus, Psathyrostachys and Eremopyrum to wheat have been summarized and discussed. To date, the official designated genes originated from wheat alien species include 17 stripe rust resistance genes, 35 leaf rust resistance gens, 30 stem rust resistance genes, 41 powdery mildew resistance genes, 3 Fusarium head blight-resistance genes, one wheat blast resistance gene, one Septoria tritici blotch resistance genes, one Septoria nodorum blotch resistance gene, 4 tan spot resistance genes, 2 eyespot resistance genes, one wheat spindle streak mosaic virus resistance gene, 2 wheat streak mosaic virus resistance genes and 2 cereal yellow dwarf resistance genes. Names and the chromosomal locations of these disease resistance genes were inducted. Moreover, the utilization of these genes in wheat breeding has also been reviewed and summarized. In the history of world wheat breeding, disease resistant germplasms such as wheat-rye 1RS·1BL translocation, 1RS·1AL translocation and wheat-Aegilops ventricosa 2NS/2AS translocation have made outstanding contributions. However, this only benefited from the utilization of a few disease resistant genes. Compared to the number of the designated genes, relatively few disease-resistant genes have been used in wheat breeding. In this paper, the limiting factors for the underutilization are discussed. Suggestions on how to use these disease-resistant genes in the future are put forward. Meanwhile, the cloned disease-resistant genes from wheat alien species are listed. The methods of cloning these genes and the possible research hotspots in the future are also analyzed. It is believed that the development and application of wheat-wild species translocation lines without genetic drag may be an important driving force for material innovation and variety breeding in the future.
DOI:10.3864/j.issn.0578-1752.2020.07.001URL [本文引用: 1]
Wheat alien species are vast reservoir of diversity for disease and pest resistance as well as stress tolerance, which are excellent gene sources for wheat breeding. Through wide hybridization, the genes of alien species could be transferred to wheat to create wheat-alien chromosome lines such as amphiploids or partial amphiploids, additions, substitutions and translocation lines. These genetic stocks could be utilized to study chromosome behavior and genome evolution, mapping genes, and diversifying the genetic basis of wheat for diseases and pest resistance, as well as yield and quality improvement. In order to understand the progress of wheat wide hybridization and useful gene transfer from alien species to wheat, in this paper, the classification of the tribe Triticeae, the definition and significance of wheat wide hybridization, alien transfers progress from species belonging to genera Aegilops, Secale, Thinopyrum, Dasypyrum, Agropyron, Hordeum, Elymus, Leymus, Psathyrostachys and Eremopyrum to wheat have been summarized and discussed. To date, the official designated genes originated from wheat alien species include 17 stripe rust resistance genes, 35 leaf rust resistance gens, 30 stem rust resistance genes, 41 powdery mildew resistance genes, 3 Fusarium head blight-resistance genes, one wheat blast resistance gene, one Septoria tritici blotch resistance genes, one Septoria nodorum blotch resistance gene, 4 tan spot resistance genes, 2 eyespot resistance genes, one wheat spindle streak mosaic virus resistance gene, 2 wheat streak mosaic virus resistance genes and 2 cereal yellow dwarf resistance genes. Names and the chromosomal locations of these disease resistance genes were inducted. Moreover, the utilization of these genes in wheat breeding has also been reviewed and summarized. In the history of world wheat breeding, disease resistant germplasms such as wheat-rye 1RS·1BL translocation, 1RS·1AL translocation and wheat-Aegilops ventricosa 2NS/2AS translocation have made outstanding contributions. However, this only benefited from the utilization of a few disease resistant genes. Compared to the number of the designated genes, relatively few disease-resistant genes have been used in wheat breeding. In this paper, the limiting factors for the underutilization are discussed. Suggestions on how to use these disease-resistant genes in the future are put forward. Meanwhile, the cloned disease-resistant genes from wheat alien species are listed. The methods of cloning these genes and the possible research hotspots in the future are also analyzed. It is believed that the development and application of wheat-wild species translocation lines without genetic drag may be an important driving force for material innovation and variety breeding in the future.
DOI:10.2135/cropsci2016.09.0819URL [本文引用: 1]
DOI:10.3389/fpls.2019.01324URLPMID:31695716 [本文引用: 1]
Prompted by recent changes in climate trends, cropping areas, and management practices, Fusarium head blight (FHB), a threatening disease of cereals worldwide, is also spreading in unusual environments, where bread wheat (BW) and durum wheat (DW) are largely cultivated. The scarcity of efficient resistance sources within adapted germplasm is particularly alarming for DW, mainly utilized for human consumption, which is therefore at high risk of kernel contamination by health-dangerous mycotoxins (e.g., deoxynivalenol = DON). To cope with this scenario, we looked outside the wheat primary gene pool and recently transferred an exceptionally effective FHB resistance QTL (Fhb-7EL) from Thinopyrum elongatum 7EL chromosome arm onto a Thinopyrum ponticum 7el1L arm segment, containing additional valuable genes (including Lr19 for leaf rust resistance and Yp for yellow pigment content), distally inserted onto 7DL of BW lines. Two such lines were crossed with two previously developed DW-Th. ponticum recombinants, having 7el1L distal portions on 7AL arms. Genomic in situ hybridization (GISH) analysis showed homologous pairing, which is enabled by 7el1L segments common to the BW and DW recombinant chromosomes, to occur with 42-78% frequency, depending on the shared 7el1L amount. Aided by 7EL/7el1L-linked markers, 7EL+7el1L tetraploid recombinant types were isolated in BC1 progenies to DW of all cross combinations. Homozygous 7EL+7el1L recombinant plants and null segregates selected in BC2F2 progenies were challenged by Fusarium graminearum spike inoculation to verify the Fhb-7EL efficacy in DW. Infection outcomes confirmed previous observations in BW, with >90% reduction of disease severity associated with Fhb-7EL presence vs. its absence. The same differential effect was detected on seed set and weight of inoculated spikes, with genotypes lacking Fhb-7EL having approximately 80% reduction compared with unaffected values of Fhb-7EL carriers. In parallel, DON content in flour extracts of resistant recombinants averaged 0.67 ppm, a value >800 times lower than that of susceptible controls. Furthermore, as observed in BW, the same Fhb-7EL also provided the novel DW recombinants with resistance to Fusarium crown rot ( approximately 60% symptom reduction) as from seedling infection with Fusarium culmorum. Through alien segment stacking, we succeeded in equipping DW with a very effective barrier against different Fusarium diseases and other positive attributes for crop security and safety.
DOI:10.1111/tpj.14444URLPMID:31259444 [本文引用: 3]
Centromeres mediate chromosome attachment to microtubules and maintain the integrity of chromosomes for proper segregation of the sister chromatids during cell division. Advances in the assembly of Triticeae genome sequences combined with the capacity to recover hybrid species derived from very distantly related species provides potential experimental systems for linking retrotransposon amplification and repositioning of centromeres via non-mendelian inheritance in partial amphiploid breeds. The decaploid tall wheatgrass (Thinopyrum ponticum) is one of the most successfully used perennial species in wheat breeding for generating translocation lines with valuable agronomic traits. We found that wheat centromere retrotransposons CRW and Quinta widely occur within the tall wheatgrass genome. In addition, one of the genome donors to Th. ponticum, Pseudoroegneria stipifolia (StSt), has been shown to have Abigail and a satellite repeat, CentSt. We also found two other centromeric retrotransposons, Abia and CL135 in Th. ponticum by ChIP-seq. Examination of partial amphiploid lines that were generated in the 1970s demonstrated extensive modification in centromere sequences using CentSt, Abigail and Abia as probes. We also detected that St-genome chromosomes were more enriched with Abigail and CentSt, whereas E-genome chromosomes were enriched with CRW and Quinta in tall wheatgrass and its closer relatives. It can be concluded that bursts of transposition of retrotransposons and repositioning of centromeres via non-mendelian segregation are common in partial amphiploids derived from interspecific hybrids. Practically speaking, our study reveals that the existence of homologous centromere functional sequences in both a donor and its receptor can substantially contribute to the successful transfer of alien genes into crop species. OPEN RESEARCH BADGES: This article has earned an Open Data Badge for making publicly available the digitally-shareable data necessary to reproduce the reported results. The data is available at https://www.ncbi.nlm.nih.gov/sra/SRR9089557; https://www.ncbi.nlm.nih.gov/sra/SRR9089558; https://www.ncbi.nlm.nih.gov/sra/SRR9089559; https://www.ncbi.nlm.nih.gov/sra/SRR9089560; https://www.ncbi.nlm.nih.gov/sra/SRR9089561; https://www.ncbi.nlm.nih.gov/sra/SRR9089562; https://www.ncbi.nlm.nih.gov/sra/SRR9089563; https://www.ncbi.nlm.nih.gov/sra/SRR9089564; https://www.ncbi.nlm.nih.gov/nuccore/MK999394; https://www.ncbi.nlm.nih.gov/nuccore/MK999395; https://www.ncbi.nlm.nih.gov/nuccore/MK999396.
DOI:10.1105/tpc.107.051813URLPMID:17586655 [本文引用: 1]
Bread wheat (Triticum aestivum) is a hexaploid species with A, B, and D ancestral genomes. Most bread wheat genes are present in the genome as triplicated homoeologous genes (homoeologs) derived from the ancestral species. Here, we report that both genetic and epigenetic alterations have occurred in the homoeologs of a wheat class E MADS box gene. Two class E genes are identified in wheat, wheat SEPALLATA (WSEP) and wheat LEAFY HULL STERILE1 (WLHS1), which are homologs of Os MADS45 and Os MADS1 in rice (Oryza sativa), respectively. The three wheat homoeologs of WSEP showed similar genomic structures and expression profiles. By contrast, the three homoeologs of WLHS1 showed genetic and epigenetic alterations. The A genome WLHS1 homoeolog (WLHS1-A) had a structural alteration that contained a large novel sequence in place of the K domain sequence. A yeast two-hybrid analysis and a transgenic experiment indicated that the WLHS1-A protein had no apparent function. The B and D genome homoeologs, WLHS1-B and WLHS1-D, respectively, had an intact MADS box gene structure, but WLHS1-B was predominantly silenced by cytosine methylation. Consequently, of the three WLHS1 homoeologs, only WLHS1-D functions in hexaploid wheat. This is a situation where three homoeologs are differentially regulated by genetic and epigenetic mechanisms.
DOI:10.1146/annurev.cellbio.042308.113414URLPMID:19572810 [本文引用: 1]
Chloroplasts are the defining organelle of photoautotrophic plant cells. Photosynthetic light reactions and electron transport are the functions of an elaborate thylakoid membrane system inside chloroplasts. The lipid composition of photosynthetic membranes is characterized by a substantial fraction of nonphosphorous galactoglycerolipids reflecting the need of sessile plants to conserve phosphorus. Lipid transport and assembly of glycerolipids play an essential role in the biogenesis of the photosynthetic apparatus in developing chloroplasts. During chloroplast biogenesis, fatty acids are synthesized in the plastid and are exported to the endoplasmic reticulum, where they are incorporated into membrane lipids. Alternatively, lipids can also be assembled de novo at the inner envelope membrane of plastids in many plants. A rich repertoire of lipid exchange mechanisms involving the thylakoid membranes, the chloroplast inner and outer envelope membranes, and the endoplasmic reticulum is emerging. Studies of thylakoid biogenesis provide new insights into the general mechanisms of intermembrane lipid transfer.
[本文引用: 1]
DOI:10.1126/science.1183700URLPMID:20150489 [本文引用: 1]
To feed the several billion people living on this planet, the production of high-quality food must increase with reduced inputs, but this accomplishment will be particularly challenging in the face of global environmental change. Plant breeders need to focus on traits with the greatest potential to increase yield. Hence, new technologies must be developed to accelerate breeding through improving genotyping and phenotyping methods and by increasing the available genetic diversity in breeding germplasm. The most gain will come from delivering these technologies in developing countries, but the technologies will have to be economically accessible and readily disseminated. Crop improvement through breeding brings immense value relative to investment and offers an effective approach to improving food security.
DOI:10.1111/j.1744-7909.2007.00513.xURL [本文引用: 1]
The hybrid rice (Oryza sativa L.) breeding that was initiated in China in the 1970s led to a great improvement in rice productivity. In general, it increases the grain yield by over 20% to the inbred rice varieties, and now hybrid rice has been widely introduced into Africa, Southern Asia and America. These hybrid varieties are generated through either three-line hybrid and two-line hybrid systems; the former is derived from cytoplasmic male sterility (CMS) and the latter derived from genic male sterility (GMS). There are three major types of CMS (HL, BT and WA) and two types of GMS (photoperiod-sensitive (PGMS) and temperature-sensitive (TGMS)). The BT- and HL-type CMS genes are characterized as orf79 and orfH79, which are chimeric toxic genes derived from mitochondrial rearrangement. Rf3 for CMS-WA is located on chromosome 1, while Rf1, Rf4, Rf5 and Rf6 correspond to CMS-BT, CMS-WA and CMS-HL, located on chromosome 10. The Rf1 gene for BT-CMS has been cloned recently, and encodes a mitochondria-targeted PPR protein. PGMS is thought to be controlled by two recessive loci on chromosomes 7 and 12, whereas nine recessive alleles have been identified for TGMS and mapped on different chromosomes. Attention is still urgently needed to resolve the molecular complexity of male sterility to assist rice breeding.
URLPMID:17245919 [本文引用: 1]
[本文引用: 1]
DOI:10.1093/jxb/ert333URL [本文引用: 3]
Global food security demands the development and delivery of new technologies to increase and secure cereal production on finite arable land without increasing water and fertilizer use. There are several options for boosting wheat yields, but most offer only small yield increases. Wheat is an inbred plant, and hybrids hold the potential to deliver a major lift in yield and will open a wide range of new breeding opportunities. A series of technological advances are needed as a base for hybrid wheat programmes. These start with major changes in floral development and architecture to separate the sexes and force outcrossing. Male sterility provides the best method to block self-fertilization, and modifying the flower structure will enhance pollen access. The recent explosion in genomic resources and technologies provides new opportunities to overcome these limitations. This review outlines the problems with existing hybrid wheat breeding systems and explores molecular-based technologies that could improve the hybrid production system to reduce hybrid seed production costs, a prerequisite for a commercial hybrid wheat system.
DOI:10.1038/srep11274URLPMID:26073981 [本文引用: 1]
Hybrid seeds are used for stimulated crop production, as they harness heterosis. The achievement of complete male-sterility in the female-parent and the restored-fertility in F1-hybrids are the major bottlenecks in the commercial hybrid seed production. Here, we report a male sterility-fertility restoration system by engineering the in most nutritive anther wall layer tapetum of female and male parents. In the female parent, high-level, and stringent expression of Arabidopsis autophagy-related gene BECLIN1 was achieved in the tapetum, which altered the tapetal degeneration program, leading to male sterility. This works on our previously demonstrated expression cassette based on functional complementation of TATA-box mutant (TGTA) promoter and TATA-binding protein mutant3 (TBPm3), with modification by conjugating Long Hypocotyle in Far-Red1 fragment (HFR1(NT131)) with TBPm3 (HFR1(NT131)-TBPm3) to exercise regulatory control over it. In the male parent, tapetum-specific Constitutive photo-morphogenesis1 (COP1) was expressed. The F1 obtained by crossing these engineered parents showed decreased BECLIN1 expression, which was further completely abolished when COP1-mutant (COP1(L105A)) was used as a male parent, leading to normal tapetal development and restored fertility. The system works on COP1-HFR1 interaction and COP1-mediated degradation of TBPm3 pool (HFR1(NT131)-TBPm3). The system can be deployed for hybrid seed production in agricultural crops.
DOI:10.1016/j.tplants.2017.10.001URLPMID:29126789 [本文引用: 1]
In many plant species, male-sterile female lines with cytoplasmic male sterility (CMS) or nuclear-controlled environment-sensitive genic male sterility (EGMS) have long been used to efficiently produce hybrids that harness hybrid vigor or heterosis. However, the underlying molecular mechanisms for these applications have only recently been uncovered in a few species. We provide here an update on the understanding of cytoplasmic-nuclear communication based on the discovery of mitochondrial CMS genes and their corresponding nuclear fertility determinants. Recent findings that uncover diverse mechanisms such as epigenetic, transcriptional, and post-transcriptional controls of EGMS by temperature and photoperiod signals are also reviewed. Furthermore, translational research that applies basic knowledge of plant male fertility control to hybrid seed production practice is highlighted.
DOI:10.2135/cropsci1976.0011183X001600050033xURL [本文引用: 1]
[本文引用: 1]
DOI:10.1016/s1360-1385(02)02252-5URLPMID:11992824 [本文引用: 1]
The introduction of hybrid crop varieties has enabled spectacular increases in productivity owing to hybrid vigor and increased uniformity. To produce hybrid seeds, a pollination control system is required to prevent unwanted self-pollination. In crop species with hermaphrodite flowers, this can be a major challenge. Over the past decade, new pollination control systems have been developed with the aid of genetic engineering, mainly based on the generation of nuclear-encoded male sterility. The successful application of these systems for large-scale hybrid seed production depends on whether the male-sterile female parent line can be multiplied efficiently and economically. In spite of its relevance, the propagation of the male-sterile line has often been overlooked in the development of pollination control systems.
DOI:10.7606/j.issn.1009-1041.2005.04.168URL [本文引用: 1]
利用C49S温光型不育系选育两系杂种小麦为小麦杂种优势的利用开辟了一条新途径。多年研究表明,由于受江汉平原气候影响及C49S不育系本身的一些缺点,限制了该类型两系杂种小麦在江汉平原生产上的应用。其主要问题是不育系育性不稳定,制种产量低而成本高、优质抗病组合少。并针对这些问题,提出了一定的解决方法,旨在为该类型杂种小麦的生产、应用提供一定的参考。
DOI:10.7606/j.issn.1009-1041.2005.04.168URL [本文引用: 1]
利用C49S温光型不育系选育两系杂种小麦为小麦杂种优势的利用开辟了一条新途径。多年研究表明,由于受江汉平原气候影响及C49S不育系本身的一些缺点,限制了该类型两系杂种小麦在江汉平原生产上的应用。其主要问题是不育系育性不稳定,制种产量低而成本高、优质抗病组合少。并针对这些问题,提出了一定的解决方法,旨在为该类型杂种小麦的生产、应用提供一定的参考。
DOI:10.1007/s11738-014-1713-7URL [本文引用: 1]
DOI:10.1111/j.1467-7652.2008.00392.xURLPMID:19121104 [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}