 ,, 陈青, 邵冰豪, 郭钰君, 张向丽, 杜鹏飞, 朱瑶, 黄艳群,, 陈文河南农业大学牧医工程学院,郑州 450002
,, 陈青, 邵冰豪, 郭钰君, 张向丽, 杜鹏飞, 朱瑶, 黄艳群,, 陈文河南农业大学牧医工程学院,郑州 450002Effect of the Heterozygous Sex-Linked Dwarf Gene on Fat Deposition in Normal Type Chickens
ZHU XingHao,, CHEN Qing, SHAO BingHao, GUO YuJun, ZHANG XiangLi, DU PengFei, ZHU Yao, HUANG YanQun,, CHEN WenCollege of Livestock Husbandry and Veterinary Engineering, Henan Agricultural University, Zhengzhou 450002通讯作者:
责任编辑: 林鉴非
收稿日期:2020-01-2接受日期:2020-11-16网络出版日期:2021-01-01
| 基金资助: |
Received:2020-01-2Accepted:2020-11-16Online:2021-01-01
作者简介 About authors
朱星浩,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (846KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
朱星浩, 陈青, 邵冰豪, 郭钰君, 张向丽, 杜鹏飞, 朱瑶, 黄艳群, 陈文. 杂合型伴性矮小基因对正常体型鸡脂肪沉积的影响[J]. 中国农业科学, 2021, 54(1): 213-223 doi:10.3864/j.issn.0578-1752.2021.01.016
ZHU XingHao, CHEN Qing, SHAO BingHao, GUO YuJun, ZHANG XiangLi, DU PengFei, ZHU Yao, HUANG YanQun, CHEN Wen.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】通过矮小型母本向正常体型公鸡导入杂合状态的dw基因,以探究杂合型伴性矮小基因对正常体型鸡的脂肪沉积及肌肉品质的影响。【前人研究进展】性连锁矮小型(sex-linked dwarf gene,dw基因)鸡以其肉质优良、低饲料消耗、低基础代谢率、高产蛋率和经济效益等育种价值,在当代家禽业中得到了广泛的研究和应用[1,2,3,4]。固始鸡和广西瑶鸡均是我国著名的地方优良品种。固始鸡具有肉质优良、脂肪含量适中等特点,但性成熟较晚;广西瑶鸡具有耐粗饲、皮脂薄、肉质细嫩的特点,是培育低脂优质肉鸡的良好遗传素材[5]。在生产上通常选择用产蛋性能高的性连锁型矮小鸡作为母本,地方鸡种作为父本进行杂交配套,后代公鸡表现为体型正常的杂合型,且能保持地方鸡种的外貌特征,是保护和利用地方鸡种的很好的途径[6]。性连锁型矮小鸡,是由于生长激素受体(growth hormone receptor,GHR)基因发生了错义突变,导致生长激素受体功能丧失,从而影响生长激素(growth hormone,GH)的分泌[7,8,9,10,11]。dw基因呈隐性伴性遗传,是目前发现的唯一对鸡健康无害的矮小基因[12]。dw基因在1935年首次被发现[13],在8—10周龄时,纯合矮小鸡与正常鸡在表型上有显著区别[14]。纯合dw基因可缩短成年鸡只的跖骨长度[15]、减轻体重[12],降低母鸡产蛋期的采食量,从而降低规模化、产业化过程中的饲料成本,并有效地提高养殖效率[16]。有研究报道纯合矮小鸡在生长阶段脂肪沉积能力远远高于正常体型鸡[4, 17],其体脂含量约为正常鸡的1.5倍[18],其肌内脂肪(IMF)含量也得到提高[19]。IMF在纯合矮小鸡体内的更多沉积可能受到多种信号通路和昼夜节律相关基因的部分调控[19]。有研究报道杂合型伴性矮小鸡虽表现为正常体型,但体重略低于正常型[20]。LILBURN等发现2—8周龄的杂合型正常鸡比纯合型正常肉鸡有更多的腹部脂肪[21]。也有报道杂合伴性矮小公鸡胴体特征和生长速率与正常型公鸡无异,但正常型公鸡与杂合型公鸡的脂酶活性存在一定的差异[22]。【本研究切入点】目前关于杂合型伴性矮小基因对鸡的脂肪沉积效应尚没有系统的研究。本研究将正常型母鸡(ZDWW)和矮小型母鸡(ZdwW)分别与正常体型的地方品种公鸡(ZDWZDW)杂交,将杂交后代在同一条件下饲养。对杂交后代进行体尺指标、血清生化指标、肌纤维特性、胸肌、腿肌IMF含量的测定,并对体脂含量及其动态变化进行研究。【拟解决的关键问题】揭示矮小型母本导入杂合状态的dw基因对后代的脂肪沉积及肌肉品质的影响,掌握杂合型正常鸡的脂肪沉积动态变化的规律,从而为利用母本矮小型生产优质肉鸡和地方鸡种研究奠定基础。1 材料与方法
1.1 试验动物
在35周龄,选用正常体型的固始公鸡(ZDWZDW)分别与正常型母鸡(固始鸡,G;广西瑶鸡,Y;ZDWW)和矮小型母鸡(快大型矮小鸡,A;ZdwW)进行杂交,另选取正常体型的广西瑶鸡(ZDWZDW)分别与正常型母鸡(固始鸡,G;含有12.5%的海兰粉血统的固始鸡与含浅芦花基因的高产蛋鸡的杂交后代,MH;ZDWW)和矮小型母鸡(矮小型黄麻肉鸡,R;矮小型黄麻肉鸡与含浅芦花基因的高产蛋鸡的杂交后代,RH;ZdwW)进行杂交,杂交均以人工授精的方式进行。分别从不同杂交群体中各收集种蛋200枚进行同批孵化,将同批孵化出的1日龄雏鸡在同一饲养条件下分开笼养,自由采食,自由饮水,执行正常的免疫程序,实施规范的饲养管理。试验地点在河南三高农牧股份有限公司,试验期为120 d,其中0—7周龄、8—17周龄分别饲喂育雏料(810饲料)和育成料(813饲料),日粮组成及营养水平见表1。Table 1
表1
表1日粮组成及营养水平
Table 1
| 营养指标 Nutrition indicators | 0-7周龄 0-7 weeks | 8-17周龄 8-17 weeks | 营养水平 Nutrient level | 0-7周龄 0-7 weeks | 8-17周龄 8-17 weeks | |
|---|---|---|---|---|---|---|
| 玉米 Corn grain (%) | 33.65 | 55.17 | 代谢能 ME(MJ·Kg-1) | 11.28 | 12.17 | |
| 43%豆粕 43% soybean meal (%) | 6.50 | — | 粗蛋白 CP (%) | 14.94 | 13.31 | |
| 46%豆粕 46% soybean meal (%) | 13.65 | 9.91 | 钙 Ca (%) | 1.00 | 0.74 | |
| 米糠油 Rice bran oil (%) | — | 2.00 | 总磷 TP (%) | 0.60 | 0.47 | |
| 玉米蛋白饲料 Corn protein feed (%) | 16.50 | 2.21 | 有效磷 A-P (%) | 0.09 | 0.10 | |
| 玉米蛋白粉 Corn gluten meal (%) | 1.50 | 0.87 | 赖氨酸 Lysine (%) | 0.87 | 0.62 | |
| 次粉 Wheat middling and reddog (%) | 24.00 | 20.00 | 蛋氨酸+胱氨酸 Methionine + Cystine (%) | 0.68 | 0.59 | |
| 石粉 Stone powder (%) | 1.53 | 1.44 | ||||
| 干酒槽及其可溶物 DDGs (%) | — | 6.00 | ||||
| 磷酸氢钙 NaHCo3 (%) | 1.20 | 0.57 | ||||
| 99%蛋氨酸 99% Methionine (%) | 0.15 | 0.13 | ||||
| 98%赖氨酸 98% Lysine (%) | 0.30 | — | ||||
| 预混料 Premix 1) (%) | 1.02 | 1.70 | ||||
| 合计 Total | 100.00 | 100.00 |
新窗口打开|下载CSV
1.2 体尺指标的测量
于60日龄、90日龄和120日龄从固始鸡和广西瑶鸡的杂交后代中分别随机选取100只后代(公母各半)进行体尺指标的测定。主要测定指标包括:胫长、胫围。测定方法如下:胫长:从胫部上关节到第三、四趾间的直线距离。胫围:胫骨中部的周长。1.3 样品的采集
分别于60日龄和90日龄从固始公鸡与正常型和矮小型母鸡杂交的每个群体中分别随机选取10只后代(公母各半)进行屠宰。另于120日龄从固始鸡和广西瑶鸡的杂交后代中各选取鸡只10只(公母各半)进行屠宰并收集颈静脉血样至促凝管中。取同侧同一位置胸肌组织块(长2 cm,厚0.5 cm,宽0.5 cm)放置4%多聚甲醛溶液保存以备石蜡切片制作,同时分离胸肌、腿肌用保鲜膜包裹放置-20℃冰箱保存以备进行肌内脂肪含量分析。1.4 体脂指标的测定
屠宰后,分离需测组织并称重,主要测定指标包括:活重、全净膛重、腹脂重、皮下脂肪厚、肌间脂肪宽。测定方法如下:活重:禁食12 h后的体重。全净膛重:屠体重为减去内脏器官、头、脚,保留肺脏、肾脏的重量。腹脂重:包括腹脂(板油)及肌胃外脂肪的重量。皮下脂肪厚:从尾根部切线向上沿第一切线剥离两侧皮肤,用游标卡尺测量此处的皮脂厚。肌间脂肪宽:将胸部的皮掀开,在胸骨侧突的部位用游标卡尺测量脂肪带的宽度。1.5 血清生化指标的测定
收集颈静脉血液10 mL于促凝离心管中倾斜静置,以4℃,3 000 r/min离心10 min,取上部血清于-20℃冰箱保存,然后用全自动生化分析仪分析测试血清生化指标。主要测定指标为总胆固醇(TC)、甘油三酯(TG)、高密度脂蛋白(HDL)、低密度脂蛋白(LDL)含量。1.6 肌内脂肪含量的测定
每只鸡胸肌、腿肌样品各取10 g,采用索式提取法提取其肌内脂肪并测定其含量,测定方法参考肉与肉制品游离脂肪含量的测定(国标GB/T-6434-2006)。每个样品测定3次,以平均值作为样品的肌内脂肪含量。根据公式:肌内脂肪含量IMF(100%)=(提取前总重-提取后的总重)/提取干物质总重×100%,计算干物质中IMF的含量。1.7 肌纤维特性分析
常规方法制备石蜡切片(厚度为2 μm),进行H.E.染色,将石蜡切片置于10×10荧光倒置显微镜下,随机选取3个视野并保存图像,用图像分析软件(Image J)测量单个图像总面积(S),并统计其中肌纤维数目(N),计算肌纤维密度(d)。利用Image J进行图像分析,分别随机抽测50根肌纤维横截面积Si,然后用公式计算出肌纤维横截面的直径Di,再计算个体胸肌的肌纤维平均直径。d=S/N;Di=2(Si/π)0.5;平均肌纤维直径D=ΣD i/50。1.8 数据统计分析
试验数据采用 Excel 2016软件进行整理后,使用SPSS 22.0生物统计软件进行双因素和单因素方差分析,并用Duncan多重比较进行显著性检验。结果以平均值±标准差(mean ± SD)表示。2 结果
2.1 F1代表型特征
正常型父本与正常型母本(后代简称母本正常型群体)和矮小型母本(后代简称母本矮小型群体)杂交的F1后代,均表现为正常体型。其中母本矮小型群体中的公鸡(ZDWZdw)虽表现为正常型,但其含有隐性的伴性矮小基因(简称dw杂合子),而母本正常型群体中公鸡基因型为ZDWZDW(简称DW纯合子)。其杂交遗传模式见图1。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1杂交遗传模式示意图
Fig. 1Schematic diagram of hybrid genetic pattern
2.2 不同日龄不同群体鸡只体尺指标的比较
整合母本正常型和母本矮小型群体的体尺数据(包含来自固始鸡和广西瑶鸡为父本的杂交群体)发现:在60日龄、90日龄和120日龄时,公鸡的胫长、胫围均显著大于母鸡(P<0.01,表2),母本矮小型群体的胫长、胫围均大于母本正常型群体。在60日龄的公鸡中,母本矮小型公鸡的胫长显著大于母本正常型公鸡(P<0.05,表2),母本矮小型公鸡的胫围显著大于母本正常型公鸡(P<0.01,表2);在母鸡中,母本矮小型母鸡的胫长、胫围显著大于母本正常型母鸡(P<0.01,表2)。Table 2
表2
表2不同日龄不同群体鸡只体尺性状的比较
Table 2
| 日龄 Age | 项目 Project | 性别 Gender | 群体 Population | 显著性Significant | |||
|---|---|---|---|---|---|---|---|
| 母本矮小型 The dwarf type of female parent | 母本正常型 The normal type of female parent | 群体 Population | 性别 Gender | 群体×性别 Population×Gender | |||
| 60 | 胫长 Shank length (mm) | 公Male | 69.19±4.81*## | 67.91±4.83## | 0.000 | 0.000 | 0.128 |
| 母Female | 64.26±4.72** | 61.73±4.66 | |||||
| 胫围 Shank circumference (cm) | 公Male | 3.60±0.26**## | 3.43±0.29## | 0.000 | 0.000 | 0.766 | |
| 母Female | 3.27±0.22** | 3.12±0.21 | |||||
| 90 | 胫长 Shank length (mm) | 公Male | 87.59±7.42## | 86.79±6.25## | 0.008 | 0.000 | 0.325 |
| 母Female | 75.88±4.82** | 74.13±4.91 | |||||
| 胫围 Shank circumference (cm) | 公Male | 4.11±0.25*## | 3.95±0.25## | 0.000 | 0.000 | 0.996 | |
| 母Female | 3.67±0.20** | 3.51±0.23 | |||||
| 120 | 胫长 Shank length (mm) | 公Male | 94.99±5.49## | 94.00±5.79## | 0.004 | 0.000 | 0.520 |
| 母Female | 77.55±4.96** | 76.00±5.15 | |||||
| 胫围 Shank circumference (cm) | 公Male | 4.57±0.24*## | 4.41±0.28## | 0.000 | 0.000 | 0.684 | |
| 母Female | 3.98±0.22** | 3.80±0.21 | |||||
新窗口打开|下载CSV
在90和120日龄时,在公鸡中,母本矮小型公鸡与母本正常型公鸡间的胫长无显著性差异,母本矮小型公鸡的胫围显著大于母本正常型公鸡(P<0.05,表2);在母鸡中,母本矮小型母鸡的胫长、胫围显著大于母本正常型母鸡(P<0.01,表2)。
2.3 固始鸡杂交后代脂肪沉积的动态变化
选择固始鸡杂交后代进行母本正常型和母本矮小型杂交后代的脂肪沉积规律的分析发现:在母本正常型群体中,公鸡的腹脂重和腹脂率一直处于低水平,甚至120日龄时的腹脂重和腹脂率均显著低于60日龄 (P<0.05,图2-A、B);公鸡的皮下脂肪厚、肌间脂肪宽在不同日龄间差异不显著(图2-C、D);而母鸡的体脂总体呈现随日龄的增加而上升的趋势,120日龄时母鸡的体脂指标(腹脂重、腹脂率、皮下脂肪厚和肌间脂肪宽)较60日龄、90日龄均有显著的提升(P<0.05,图2);母鸡60日龄和90日龄间的体脂指标差异不显著。而在母本矮小型群体中,公、母鸡均呈现了与母本正常型群体中的母鸡相似的脂肪沉积变化特性。母本矮小型群体的公、母鸡腹脂重、皮下脂肪厚和肌间脂肪宽均随日龄逐渐升高,特别在120日龄时显著升高。120日龄的腹脂重、皮下脂肪厚和肌间脂肪宽均显著高于60或90日龄(P<0.05,图2-A、C、D);此外,母本矮小型基因也一定程度上提高了后代鸡只90日龄的脂肪沉积,特别是90日龄的公鸡肌间脂肪宽显著高于60日龄(P<0.05)。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2固始鸡杂交后代不同时期的体内脂肪沉积动态变化情况
A. 腹脂重;B. 腹脂率;C. 皮下脂肪厚;D. 肌间脂肪宽;相同日龄、相同性别不同群体间,*表示差异显著(P<0.05),**表示差异极显著(P<0.01);相同群体、相同性别不同日龄间,不同小写字母表示差异显著(P<0.05);相同日龄、相同群体不同性别间,##表示差异极显著(P<0.01);腹脂率=腹脂重/活重×100%
Fig. 2Dynamic changes of body fat deposition in different periods of hybrid offspring of Gushi chickens
A. Abdominal fat weight; B. Percentage of abdominal fat; C. Subcutaneous fat thickness; D. Intermuscular fat width; In different groups of the same age and sex, * means significant difference (P<0.05), ** means extremely significant difference (P<0.01); In different age of the same population and sex, different lowercase letters means significant difference (P<0.05); In different sex of the same age and population, ## means extremely significant difference (P<0.01). Percent of abdominal fat= abdominal fat weight/ body weight×100%
进行同一群体、同一日龄不同性别鸡只间的比较,发现在母本正常型群体中,120日龄时母鸡腹脂重和腹脂率极显著高于公鸡(P<0.01,图2-A、B);皮下脂肪厚、肌间脂肪宽在三个时间点公、母间均无显著差异(图2-C、D)。而在母本矮小型群体中,60日龄、90日龄、120日龄公、母鸡间的体脂指标(包括腹脂重、腹脂率、皮下脂肪厚和肌间脂肪宽)均无显著差异(P>0.05,图2)。
进行同一日龄、同一性别不同群体鸡只的比较发现,母本矮小型群体公鸡(dw杂合子)在90日龄和120日龄时各项体脂指标(腹脂重、腹脂率、皮下脂肪厚和肌间脂肪宽)均显著或极显著高于同日龄的母本正常型群体公鸡(DW纯合子),而仅120日龄母本矮小型母鸡的腹脂重显著高于母本正常型群体中的母鸡(P<0.05,图2-A)。
2.4 120日龄不同群体鸡只体脂指标的比较
进一步整合120日龄固始鸡和广西瑶鸡杂交后代的体脂数据,进行双因素方差分析(表3),发现群体因素和性别因素对活重和脂肪沉积相关指标(包括腹脂重、腹脂率、肌间脂肪宽和皮下脂肪厚)的效应均达到显著水平,此外皮下脂肪厚的群体和性别的互作效应也达到显著水平,故对皮下脂肪厚这一指标分别以群体和性别进行单因素分析。Table 3
表3
表3性别和群体对120日龄不同杂交组合鸡脂肪沉积的影响
Table 3
| 项目 Project | 性别 Gender | 群体Population | 显著性Significant | |||
|---|---|---|---|---|---|---|
| 母本矮小型 The dwarf type of female parent | 母本正常型 The normal type of female parent | 群体Population | 性别 Gender | 群体×性别 Population×Gender | ||
| 活重 Live weight (kg) | 公Male | 2.09±0.15**## | 1.78±0.24 | 0.000 | 0.000 | 0.466 |
| 母Female | 1.71±0.20**## | 1.48±0.18 | ||||
| 腹脂重 Abdominal fat weight (g) | 公Male | 27.15±23.02** | 3.50±7.17 | 0.004 | 0.001 | 0.152 |
| 母Female | 37.22±26.68 | 28.86±19.53## | ||||
| 腹脂率 Percentage of abdominal fat (%) | 公Male | 1.83±1.55** | 0.27±0.54 | 0.033 | 0.000 | 0.095 |
| 母Female | 3.02±1.97 | 2.83±1.79## | ||||
| 肌间脂肪宽 Intermuscular fat width (mm) | 公Male | 9.89±3.97** | 4.88±3.18 | 0.000 | 0.000 | 0.138 |
| 母Female | 12.11±3.89 | 9.94±3.38## | ||||
| 皮下脂肪厚 Subcutaneous fat thickness (mm) | 公Male | 5.43±1.45** | 4.16±1.09 | 0.051 | 0.002 | 0.025 |
| 母Female | 5.72±0.73 | 5.81±1.12## | ||||
新窗口打开|下载CSV
在活重指标中,总体上公鸡的活重大于母鸡,母本矮小型群体的活重大于母本正常型群体。在公鸡中,母本矮小型公鸡的活重极显著大于母本正常型公鸡(P<0.01,表3)。在母鸡中,母本矮小型母鸡的活重也大于母本正常型母鸡(P<0.01,表3)。
进一步的分析发现,在母本正常型群体中,公鸡的脂肪沉积少,母鸡的腹脂重、腹脂率、肌间脂肪宽、皮下脂肪厚均显著高于公鸡(P<0.01,表3),腹脂重、腹脂率在公母间有近10倍的差异。而在母本矮小型群体中,公母鸡间在腹脂重、腹脂率、肌间脂肪宽、皮下脂肪厚指标上均无显著差异。母本矮小型公鸡的脂肪沉积能力较母本正常型公鸡有大幅提升,母本矮小型群体公鸡(dw杂合子)的腹脂重、腹脂率、肌间脂肪宽、皮下脂肪厚均显著高于母本正常型群体公鸡(DW纯合子,P<0.01,表3);而在母鸡中,母本矮小型群体的体脂相关指标和母本正常型差异不显著(表3)。
2.5 120日龄鸡只的血脂指标的比较
采用双因素方差分析对血脂指标进行分析,发现群体因素对几个血脂指标的效应均未达到显著水平,而性别因素对TG、HDL的效应达到显著水平,群体因素和性别因素对血脂指标的互作效应均不显著(表4)。母本矮小型与母本正常型群体间在TC、TG、LDL、HDL含量上均无显著差异。母本矮小型母鸡的TG含量显著高于公鸡(P<0.05)。母本正常型公鸡的HDL极显著高于母鸡(P<0.01)。Table 4
表4
表4性别、群体对120日龄不同杂交组合鸡血清生化指标的影响
Table 4
| 项目 Project | 性别 Gender | 群体 Population | 显著性Significant | |||
|---|---|---|---|---|---|---|
| 母本矮小型 The dwarf type of female parent | 母本正常型 The normal type of female parent | 群体 Population | 性别 Gender | 群体×性别 Population×Gender | ||
| 总胆固醇 TC (mmol·L-1) | 公 Male | 2.41±0.48 | 2.72±0.61 | 0.531 | 0.249 | 0.787 |
| 母Female | 2.91±0.79 | 3.03±2.12 | ||||
| 甘油三酯 TG (mmol·L-1) | 公Male | 0.29±0.06 | 0.30±0.09 | 0.539 | 0.023 | 0.541 |
| 母Female | 2.59±2.56## | 4.22±8.77 | ||||
| 低密度脂蛋白 LDL (mmol·L-1) | 公Male | 0.75±0.25 | 0.96±0.29 | 0.943 | 0.082 | 0.139 |
| 母Female | 1.19±0.79 | 0.99±0.38 | ||||
| 高密度脂蛋白 HDL (mmol·L-1) | 公Male | 1.55±0.27 | 1.59±0.31## | 0.581 | 0.001 | 0.373 |
| 母Female | 1.27±0.38 | 1.13±0.42 | ||||
新窗口打开|下载CSV
2.6 120日龄鸡只IMF含量的比较
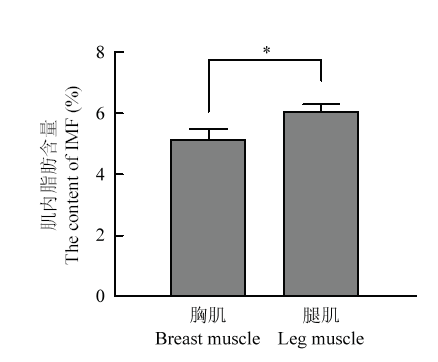
双因素方差分析(表5)显示,群体因素对胸肌IMF含量达到显著水平(P=0.017),对腿肌IMF的效应接近显著水平(P=0.076)。母本矮小型后代胸肌(P=0.017)和腿肌(P=0.076)IMF含量均高于母本正常型。其中母本矮小型母鸡与母本正常型母鸡的胸肌IMF含量的差异未达到显著水平,母本矮小型公鸡(DW杂合子)的胸肌IMF含量显著高于母本正常型公鸡(DW纯合子,P<0.01)。总体来说,腿肌的IMF含量显著高于胸肌(P<0.05,图3)。Table 5
表5
表5性别、群体对120日龄不同杂交组合鸡IMF含量的影响
Table 5
| 项目 Project | 性别 Gender | 群体 Population | 显著性 Significant | |||
|---|---|---|---|---|---|---|
| 母本矮小型 The dwarf type of female parent | 母本正常型 The normal type of female parent | 群体Population | 性别 Gender | 群体×性别 Population×Gender | ||
| 腿肌IMF The IMF of leg muscle (%) | 公Male | 6.31±1.26 | 5.04±2.48 | 0.076 | 0.499 | 0.542 |
| 母Female | 6.34±1.85 | 5.72±2.26 | ||||
| 胸肌IMF The IMF of breast muscle (%) | 公Male | 5.77±2.19* | 3.45±2.39 | 0.017 | 0.411 | 0.215 |
| 母Female | 5.51±2.27 | 4.75±2.80 | ||||
新窗口打开|下载CSV
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3120日龄杂交组合鸡胸肌、腿肌IMF含量
*表示差异显著
Fig. 3Intramuscular fat content of chicken breast muscle and leg muscle in hybrid combination at 120 days
* mean significant difference (P<0.05)
2.7 120日龄鸡只的肌纤维特性的比较
母本正常型后代肌纤维密度均呈现高于母本矮小型的趋势(P=0.089),肌纤维面积、肌纤维直径在母本矮小型和母本正常型间无显著差异。母鸡肌纤维密度显著低于公鸡(P=0.018),而其肌纤维面积(P= 0.088)、肌纤维直径(P=0.064)呈现高于公鸡的趋势(表6)。Table 6
表6
表6性别、群体对120日龄不同杂交组合鸡胸肌肌纤维特性的影响
Table 6
| 项目 Project | 性别 Gender | 群体Population | 显著性Significant | |||
|---|---|---|---|---|---|---|
| 母本矮小型 The dwarf type of female parent | 母本正常型 The normal type of female parent | 群体 Population | 性别 Gender | 群体×性别 Population×Gender | ||
| 肌纤维密度 Density of muscle fiber (根/mm2) | 公Male | 474.33±91.01 | 508.76±154.20 | 0.089 | 0.018 | 0.542 |
| 母Female | 380.53±85.11 | 452.66±97.41 | ||||
| 肌纤维面积 Area of muscle fiber (μm2) | 公Male | 1229.82±340.91 | 1190.59±414.89 | 0.509 | 0.088 | 0.778 |
| 母Female | 1438.52±363.63# | 1340.72±364.60 | ||||
| 肌纤维直径 Muscle fiber diameter (μm) | 公Male | 38.56±5.37 | 37.65±6.69 | 0.483 | 0.064 | 0.894 |
| 母Female | 41.78±5.35 | 40.44±5.37 | ||||
新窗口打开|下载CSV
3 讨论
3.1 杂合型伴性矮小基因对正常体型鸡体尺性状的影响
MARKS [15]研究发现纯合dw基因可缩短成年鸡只的跖骨长度。BLOHOWIAK等[23]研究发现纯合正常型与杂合型正常鸡的胫长差异不显著,但均大于矮小鸡。LILBURN[21]等研究发现8周龄时慢速型的纯合正常型鸡的胫长显著低于杂合型正常鸡,而快速型的纯合正常鸡的胫长略高于杂合型正常鸡。MERAT [24] 等研究发现在8、17和52周龄时,由于dw基因的导入胫长性状存在杂种优势。因此我们猜测在不同日龄间母本矮小型鸡只的胫长不仅没有短于母本正常型鸡只反而长于母本正常型鸡只。可能与dw基因的杂交优势有关,其具体的原因还值得进一步研究。3.2 杂合型伴性矮小基因对正常体型鸡脂肪沉积的影响
鸡过量脂肪主要沉积在腹部和皮下,腹脂率和皮下脂肪厚是评价家禽脂肪含量高低的重要指标[25]。在正常情况下,公鸡的脂肪沉积较少,而母鸡的脂肪沉积相对较多[26,27]。因此在生产上有通过公鸡的阉割以提高其体脂沉积[28],进而提高鸡肉风味的传统。性别对屠体性状的影响主要与雄性激素有关,雄性激素会促进肌肉发育,但会降低脂肪沉积[29]。据文献报道纯合的dw基因不仅可以缩短跖骨的长度,还可以增加胴体脂肪含量[18,19]。本研究根据不同生长速度的地方优质鸡的上市日龄及鸡只体脂沉积多发生在育成期的特点[30],首先对固始鸡杂交后代不同生长时期的脂肪沉积变化进行研究发现60日龄母本矮小型公鸡比母本正常型公鸡有更多的腹部脂肪(图2-A)。此外,本研究进一步发现母本矮小型公鸡(ZDWZdw)表现了完全不同于母本正常型公鸡(ZDWZDW)的脂肪沉积变化规律,在90日龄后脂肪沉积显著增加。其90日龄和120日龄的体脂沉积能力显著高于母本正常型群体的公鸡(图2)。显示了用正常公鸡与性连锁dw母鸡杂交的后代尽管体型正常,但杂合状态的dw基因在发挥作用。进一步整合来源于固始鸡和广西瑶鸡为父本的杂交群体分析杂合型矮小基因对120日龄鸡只脂肪沉积的效应发现,母本矮小型公鸡的脂肪沉积能力显著高于母本正常型群体的公鸡(表3),进一步显示杂合状态的dw基因可显著促进生长阶段公鸡的腹部脂肪、皮下脂肪和肌间脂肪的沉积。
3.3 杂合型伴性矮小基因对正常体型鸡IMF含量的影响
IMF是改善肌肉口感和嫩度的关键因素,其含量对评价肉的感官特性具有重要作用[31,32]。YE等[19]研究报道7周龄时,矮小鸡的IMF含量是正常鸡的1.2倍。本研究证明母本矮小型公鸡的胸肌IMF含量极显著高于母本正常型公鸡,显示隐性伴性的dw基因在正常体型的杂合公鸡IMF沉积中仍在发挥着重要的作用,也进一步显示了利用dw基因可改善公鸡的脂肪沉积特性从而提高鸡肉品质的潜在应用价值。此外本研究也发现了母本矮小型群体的母鸡(ZDWW)的脂肪沉积在一定程度上高于母本正常型群体里的母鸡(ZDWW),显示母鸡尽管不含有dw基因,但母本效应仍在发挥着一定的作用。有研究表明性连锁矮小鸡的骨骼肌纤维直径小于正常鸡,肌纤维密度显著高于正常鸡[6, 31],dw基因在8—12周龄之间对肌肉纤维发育有影响,dw基因对单个肌纤维横截面积有显著影响,而对肌纤维总数没有影响[14]。本研究发现杂合状态的dw基因对肌纤维特性无明显影响。3.4 杂合型伴性矮小基因对正常体型鸡血脂指标的影响
血清脂类生化指标是反映机体脂肪代谢的重要指标。BURGHELLE-MAYEUR等[33] 研究发现dw基因没有显著改变血浆中甘油三酯、卵磷脂、总胆固醇或极密度脂蛋白的含量。我们的研究结果与之一致,母本矮小型群体公鸡与母本正常型群体公鸡的血脂指标没有显著的差异,说明杂合伴性dw基因对正常表型鸡的血脂指标没有显著影响。本研究发现120日龄母本矮小型群体的公鸡的腹部脂肪等指标显著高于母本正常型群体的公鸡,但两者的血脂指标并没有显著的差异。进而说明了母本矮小型群体公鸡的血脂指标是独立于腹部脂肪沉积。WALTON等[34]在人类中研究发现男性的TG、TC、HDL的含量独立于体脂含量和分布。也有研究在肉鸡品系中发现血浆蛋白与体脂之间无显著关系[35]。4 结论
杂合伴性矮小型基因显著改善了正常体型公鸡的脂肪沉积特性,提高了公鸡的腹部脂肪、皮下脂肪和肌间脂肪的沉积;改善了胸肌的肌内脂肪含量;而对血脂指标无显著影响;对肌纤维特性无显著影响。提示,生产上可以用性连锁矮小鸡做母本来改善商品代公鸡的脂肪沉积。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
URLPMID:29653827 [本文引用: 1]
DOI:10.4238/gmr15049326URLPMID:28002609 [本文引用: 1]

In this study, eight inbred maize lines with high or low tolerance to cold temperature were investigated using bulk segregant analysis (BSA). Genetic diversity and genetic relationships were investigated using 100 simple sequence repeat (SSR) markers linked to cold tolerance in maize. A total of 319 alleles were identified. Among these, 128 were high coldtolerant-specific alleles and 61 were poorly coldtolerant-specific alleles, while the remaining 130 were shared between the two types. The number of alleles per locus ranged from 2 to 5, with an average of 3.19. The major allele frequency varied from 0.39 to 0.53 with an average of 0.47. The average gene diversity and polymorphic information content among all lines were 0.63 and 0.58, respectively. A dendrogram analysis identified three main clusters and most of the high tolerant inbred lines were clearly distinguished from the poorly tolerant inbred lines. In addition to the BSA, a total of 84 SSR markers were identified as high cold tolerance-specific alleles and 52 SSRs were detected as poorly cold tolerance-specific alleles. Of these, bnlg1273, umc1124, dupssr21, mmc0251, mmc0181, and phi041 have great potential for being molecular markers for cold tolerance in maize. Our results were in agreement with results previously reported for SSR markers linked with cold tolerance in maize. The identification and characterization of high and poorly cold tolerant maize lines based on SSR markers will be useful for future maize breeding studies.
URLPMID:31115461 [本文引用: 1]
URL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
DOI:10.3390/ijms17030276URLPMID:26927061 [本文引用: 1]
The sex-linked dwarf (SLD) chicken is an ideal model system for understanding growth hormone (GH)-action and growth hormone receptor (GHR) function because of its recessive mutation in the GHR gene. Skeletal muscle mass is reduced in the SLD chicken with a smaller muscle fiber diameter. Our previous study has presented the mRNA and miRNA expression profiles of the SLD chicken and normal chicken between embryo day 14 and seven weeks of age. However, the molecular mechanism of GHR-deficient induced muscle mass loss is still unclear, and the key molecules and pathways underlying the GHR-deficient induced muscle mass loss also remain to be illustrated. Here, by functional network analysis of the differentially expressed miRNAs and mRNAs between the SLD and normal chickens, we revealed that let-7b, miR-128 and the MAPK pathway might play key roles in the GHR-deficient induced muscle mass loss, and that the reduced cell division and growth are potential cellular processes during the SLD chicken skeletal muscle development. Additionally, we also found some genes and miRNAs involved in chicken skeletal muscle development, through the MAPK, PI3K-Akt, Wnt and Insulin signaling pathways. This study provides new insights into the molecular mechanism underlying muscle mass loss in the SLD chickens, and some regulatory networks that are crucial for chicken skeletal muscle development.
DOI:10.1210/mend.7.11.8114754URLPMID:8114754 [本文引用: 1]
Sex-linked dwarfism in chickens is a form of GH resistance that resembles the Laron syndrome in humans. The dwarfism found in chickens is due to a mutant gene (dw) carried on the sex chromosome. The homozygous dwarf (dwdw) chicken is characterized by reductions in stature and plasma insulin-like growth factor-I (IGF-I) levels. Despite the absence of hepatic GH-binding activity, Southern blot analysis shows that there is no gross structural change in the gene for the GH receptor (GHR) in this strain of dwdw chicken. GH-dependent IGF-I production can be restored in cultured dwdw hepatocytes after transfection and transient expression of a chicken GHR (cGHR) cDNA, indicating that other factors that participate in GH-mediated IGF-I synthesis are intact. Northern blot analysis of liver, muscle, fat, and pituitary RNA from normal (DwDw) chickens shows a major transcript of 4.3 kilobases (kb) and three minor transcripts (0.8, 1.7, and 3.2 kb), which correspond to the cGHR. In contrast, the 0.8-kb transcript is the major cGHR transcript expressed in these tissues from dwdw chickens. Northern blot analysis with domain-specific probes shows that the 0.8-kb transcript in DwDw and dwdw liver contains only a small portion of the extracellular domain of the cGHR. A cDNA clone encoding this transcript has been isolated from a liver library prepared from a normal chicken.(ABSTRACT TRUNCATED AT 250 WORDS)
DOI:10.1080/15476286.2019.1572440URLPMID:30764709 [本文引用: 1]
Natural antisense transcripts (NATs) are widely present in mammalian genomes and act as pivotal regulator molecules of gene expression. However, studies on NATs in the chicken are relatively rare. We identi fi ed a novel antisense transcript in the chicken, designated GHR-AS-EST, transcribed from the growth hormone receptor (GHR) locus, which encodes a well-known regulatory molecule of muscle development and fat deposition. GHR-AS-EST is predominantly expressed in the chicken liver and muscle tissues. GHR-AS-EST sequence conservation among vertebrates is weak. GHR-AS-EST forms an RNA-RNA duplex with GHBP to increase its stability, and regulates the expression of GHR sense transcripts at both the mRNA and protein levels. Further, GHR-AS-EST promotes cell proliferation by stimulating the expression of signaling factors in the JAK2/STAT pathway, and contributes to fat deposition via downregulating the expression of signaling factors in the JAK2/SOCS pathway in LMH hepatocellular carcinoma cells. We expect that the discovery of a NAT for a regulatory gene associated with cell proliferation and lipolysis will further our understanding of the molecular regulation of both muscle development and fat deposition.
DOI:10.1093/jas/skaa397URLPMID:33313796 [本文引用: 1]
The objective was to evaluate wool (Dorset and Rambouillet) and hair (Dorper, Katahdin, and White Dorper) breeds for their ability to complement Romanov germplasm in two distinct production systems by estimating direct sire and grandsire effects on lamb growth, survival, and ewe productivity traits. Rams of the five breeds (n = 75) were mated to Romanov ewes (n = 459) over a 3-yr period to produce five types of crossbred lambs (n = 2,739). Sire breed (P > 0.06) did not impact body weight or survival traits of the first-generation crossbred (F1) lambs. The productivity of retained crossbred ewes (n = 830) mated to Suffolk and Texel terminal sires was evaluated at 1, 2, and 3 yr of age in each production system. In the intensive production system, labor and harvested feed were provided for sheep that lambed in March in barns, and ewes were limited to rearing two lambs with additional lambs reared artificially. Ewes in the extensive production system lambed in May on pasture and were responsible for rearing all lambs born with no labor or supplemental feed provided before weaning. A total of 1,962 litters and 4,171 lambs from 2,229 exposures to two terminal sire breeds (Suffolk and Texel) were produced in the experiment. Crossbred ewes in the intensive production system were mated in October, resulting in larger litter sizes than crossbred ewes mated in December for the extensive production system. However, single- and twin-born lamb mortality was similar between the two systems that differed greatly in labor, feed, and facilities. Lambs produced in the intensive system received concentrate feed from an early age and were heavier at 24 wk of age than lambs produced in the extensive system. These outcomes resulted in greater 24-wk litter weight in the intensive than in the extensive system (P < 0.0001). Unexpectedly, the relative performance of crossbred types did not differ importantly between production systems. White Dorper x Romanov crossbred ewes had numerical advantages in productivity in each system; however, differences between ewe types were not significant. In the extensive system, without labor and shelter at lambing or supplemental feed until weaning, 3-yr-old crossbred ewes of all types averaged 1.78 lambs marketed per ewe lambing, and 40% of the ewes that gave birth to triplets weaned their entire litters. These results document that prolific sheep and extensive systems can be successfully combined if appropriate crossbred types are used.
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
URL [本文引用: 2]
URL [本文引用: 1]
URL [本文引用: 1]
DOI:10.1080/00071668408454863URLPMID:6733556 [本文引用: 1]
Comparisons of age, food intake, body weight and body composition at the onset of sexual maturity were made between dwarf and normal chickens from lines selected for high or low juvenile body weight. At the same age birds which had started to lay were significantly heavier and contained significantly more abdominal and carcass fat than those which were not laying. Differences between normal and dwarf pullets within lines varied with genetic background, indicating differences between lines in meeting a body weight or a body composition requirement, or both, for the onset of sexual maturity.
URLPMID:7465494 [本文引用: 2]
[本文引用: 4]
DOI:10.3382/ps.0511701URL [本文引用: 1]
[本文引用: 2]
DOI:10.1093/jn/114.1.132URLPMID:6693975 [本文引用: 1]
The growth and lipogenic responses of broiler progeny from normal (DwDw male, Dw-female) and dwarf (Dwdw male, Dw-* female) dams to high and low dietary protein were studied. Body weight gains for the DwDw compared to Dwdw male progeny were greater from week 1 to 3, but less from week 7 to 8 and were similar for Dw-* and Dw- females. No significant effects of progeny genotype and no diet X genotype interactions were observed for feed conversion, carcass nitrogen or fat, abdominal fat, growth hormone, triiodothyronine (T3) or thyroxine (T4). Hepatic malic enzyme was elevated in DwDw male progeny compared to Dwdw males, regardless of diet, and was greater in Dw- females than in Dw-* only in response to low dietary protein. Citrate cleavage enzyme was greater in Dw-* than in Dw- females. Low protein resulted in elevated lipogenic enzyme activities, which were more pronounced in progeny of normal dams. A positive relationship observed between malic enzyme activity and T3 levels for all progeny not carrying the dw gene suggests that although circulating levels of thyroid hormones did not differ between genotypes, the cellular response to these hormones was affected by the presence of the dw gene.
DOI:10.1016/0304-3762(80)90069-3URL [本文引用: 1]
DOI:10.3382/ps.0730001URLPMID:8165154 [本文引用: 1]
The effect of genotype at the sex-linked dwarf locus on heterosis in crosses between a White Leghorn and a brown egg line for body weight, egg production, and related traits was studied. Heterozygous Dw/dw males were used to produce normal and dwarf pullets in each of the pure lines and their reciprocal crosses (eight genotype-line combinations). There were 54 pullets per combination. Line differences were significant for shank length, body weights at 8, 17, and 52 wk, age at first egg, egg number, clutch length, rate of lay, and egg weight. Heterosis was observed for all of these traits. Body weight as a covariate was not important in analyses of egg number, clutch length, and egg weight. The egg production reduction associated with the dw gene in pure lines was smaller in F1 hens. This discovery may be adequate to warrant use of dwarf crossbred hens for egg production.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1051/animres:2002035URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 2]
DOI:10.1080/00071669308417603URLPMID:8358635 [本文引用: 1]
1. The number and size of muscle fibres and the DNA concentration in m. pectoralis superficialis of normal (Dw+/-) and dwarf (dw/-) White Plymouth Rock females were investigated at the ages of 9 and 52 weeks. 2. Live weight and total meat was reduced by 37.4 and 42.8% in dw/- as compared with Dw+/- females at the age of 9 weeks. The suppressive effect of the dwarf allele on the weight of m. pectoralis superficialis was 40.6% (67.8 v. 40.3 g in Dw+/- and dw/- genotypes). This reduction in weight was accompanied by a similar decrease of DNA content (28.8 v. 17.1 mg, i.e. 40.6%), a decrease in number of muscle fibres (829 x 10(3) v. 654 x 10(3), i.e. 21.2%) and fibre diameter (30.80 v. 28.95 microns, i.e. 6.0%). In mature hens, the differences between genotypes Dw+/- and dw/- were slightly smaller. 3. The different relationships between the muscle growth traits studied are discussed.
DOI:10.1093/jn/119.10.1361URLPMID:2685198 [本文引用: 1]
The levels and fatty acid composition of lipids were determined in very low density lipoproteins (VLDL, d less than 1.006), yolk and abdominal adipose tissue of normal (Dw) and sex-linked dwarf (dw) White Leghorn laying hens. Effects of adding 4% tallow to the diet were also examined. In 40-wk-old hens, neither plasma lipids (triglycerides, phospholipids and cholesterol), VLDL levels, nor the chemical composition of VLDL was altered by the dw gene or dietary fat. Dwarfism reduced egg and yolk weights. Though the yolk lipid content was similar in normal and dwarf hens, yolk from dwarfs had slightly more phospholipids and less triglycerides than yolk from normal hens. Higher linoleic acid [18:2(n-6)] and lower oleic acid [18:1(n-9)] levels were observed in triglycerides of VLDL, yolk and adipose tissue from dwarf hens. In addition, the dietary fatty acid pattern had a greater influence on the fatty acid composition of the yolk lipid major precursors (VLDL triglycerides) in dwarf laying hens than in normal hens. These results suggest that the dwarfing gene might reduce the hepatic de novo fatty acid synthesis and/or dwarf hens might incorporate more dietary lipids into yolk than do normal hens.
DOI:10.1016/0021-9150(95)05590-sURLPMID:8579629 [本文引用: 1]
Insulin resistance and hyperinsulinaemia may be important in a cluster of coronary heart disease-related metabolic disturbances known as the insulin resistance syndrome. Whether the relationships between insulin and other features of the syndrome are independent of the amount and distribution of body fat is uncertain. We have investigated these relationships in 103 healthy males, using dual-energy X-ray absorptiometry to measure body fat directly. Volunteers underwent an intravenous glucose tolerance test (IVGTT), from which insulin sensitivity, secretion and elimination were determined by mathematical modelling analysis. Independently of adiposity and body fat distribution, serum triglyceride concentration was correlated with fasting C-peptide concentration and second-phase intravenous glucose tolerance test insulin concentration (r = 0.42, P < 0.001; r = 0.28, P < 0.05). High density lipoprotein subfraction 2 (HDL2) cholesterol was correlated with fasting C-peptide, first-phase IVGTT insulin concentration, and the hepatic insulin throughout index (r = -0.15 -0.20, -0. 20 respectively, all P < 0.05). The association of HDL2 cholesterol with the hepatic throughput index was additionally independent of serum triglyceride concentration (r = -0.18, P < 0.05). Our results suggest that relative hyperinsulinaemia leads to elevated triglyceride concentration, independently of body fat mass and distribution. Furthermore, the independent association of HDL2 cholesterol with hepatic insulin throughput confirms that hepatic insulin processing may may directly influence lipoprotein metabolism.
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}