 ,1, 贾哓玮,1, 张洪艳1, 贾羊羊1, 田士军1, 张科,2, 潘延云,1
,1, 贾哓玮,1, 张洪艳1, 贾羊羊1, 田士军1, 张科,2, 潘延云,1Genomic Profiling and Expression Analysis of Phosphatidylinositol- specific PLC Gene Families Among Chinese Spring Wheat
SI XuYang,1, JIA XiaoWei,1, ZHANG HongYan1, JIA YangYang1, TIAN ShiJun1, ZHANG Ke,2, PAN YanYun,1通讯作者:
责任编辑: 李莉
收稿日期:2020-03-23接受日期:2020-06-2网络出版日期:2020-12-16
| 基金资助: |
Received:2020-03-23Accepted:2020-06-2Online:2020-12-16
作者简介 About authors
司旭阳,E-mail:
贾哓玮,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (2746KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
司旭阳, 贾哓玮, 张洪艳, 贾羊羊, 田士军, 张科, 潘延云. 中国春小麦肌醇磷脂依赖的磷脂酶C基因的全基因组鉴定及表达分析[J]. 中国农业科学, 2020, 53(24): 4969-4981 doi:10.3864/j.issn.0578-1752.2020.24.001
SI XuYang, JIA XiaoWei, ZHANG HongYan, JIA YangYang, TIAN ShiJun, ZHANG Ke, PAN YanYun.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】小麦(Triticum aestivum)是世界上最重要的作物之一。随着世界人口的不断增长,粮食安全已成为一个主要问题。干旱、土壤盐分以及极端温度等渗透胁迫是影响植物生长最主要的不利环境[1,2]。植物肌醇磷脂信号系统在应答渗透胁迫,调节植物生长中发挥着重要的作用[3,4]。肌醇磷脂依赖性磷脂酶C(phosphatidylinositol-dependent phospholipase,PI-PLC,简称为PLC)是肌醇磷脂信号系统中的关键酶之一[3,4],研究小麦基因组中编码PLC的基因及其功能对于澄清小麦在胁迫中的应答反应和改良小麦在不利环境中的生长都有重要的理论和应用价值[1,3]。【前人研究进展】在哺乳动物中已鉴定出5种PLC亚型,即β、γ、δ、ε和ζ,被激活的PLC可以水解质膜上的4,5-磷酸磷脂酰肌醇(phosphatidylinositol 4,5-bisphosphate,PIP2)产生二酰甘油(diacylglycerol,DAG)和三磷酸肌醇(inositol 1,4,5-trisphosphate,IP3)双信使分子,分别激活蛋白激酶C(protein kinase C,PKC)和释放胞内钙库Ca2+,进而调控一系列靶蛋白和靶酶的活性,参与细胞生长、增殖、代谢、分泌、收缩等发育过程和生理过程的调节[2]。植物PLC的研究始于20世纪80年代,目前已经在多个物种——拟南芥[5]、大豆[6]、烟草[7,8]、水稻[9]、马铃薯[10]、玉米[11]、百合[12]和绿豆[13,14]中克隆获得PLC基因序列。每个植物物种几乎都由多个PLC基因组成基因家族,而各个成员可能参与不同的生理过程。以拟南芥为例,其基因组编码9个AtPLC家族成员(AtPLC1—AtPLC9),其中AtPLC4可能编码质膜相关蛋白[15],并负调控幼苗对盐的耐受性[16]。AtPLC2是组成型表达基因,其不仅参与对幼苗生长发育和内质网应激反应的调控[17],而且还通过生长素信号调节雌雄配子体的发育[18]。AtPLC5参与初级和次级根的生长调控,其过表达提高了植物的抗旱性[19]。AtPLC3也参与了根系的发育[20],AtPLC3和AtPLC7的过表达都可增加植物对干旱的耐受性,并在ABA调控的种子萌发和气孔运动中发挥作用[20,21]。【本研究切入点】与动物PLC不同,目前发现的植物PLC结构均与动物中最简单的PLC ζ亚型相似。植物PLC的作用模式与动物也不相同,植物细胞中缺乏受IP3调控的受体,IP3被进一步磷酸化形成六磷酸肌醇(inositol hexaphosphate,IP6),参与细胞内钙信号形成,但具体的作用机制尚不清楚;植物中目前也没有鉴定出PKC的同源类似物,DAG可用于磷脂的再生或继续被磷酸化形成磷脂酸(phosphatidic acid,PA),而PA被认为是植物特有的第二信使,调节植物的多种生理过程,但是目前也只鉴定了少数的PA结合蛋白,且蛋白与PA互作的结构域尚未确定[3,22-24]。因此,需要更广泛地分析植物PLC基因的功能,以诠释其作用机制。小麦TaPLC的研究起始较早,1987年,就在小麦根细胞的质膜中检测到PLC活性[25];1992年检测了小麦PLC酶活性的生化特征[26,27]。但目前仅有2个TaPLC基因被克隆,即TaPLC1和TaPLC2(GenBank:HM754654.1和HM754653.1),初步结果显示这两个基因在调节小麦幼苗生长中发挥着作用且表达受到盐或干旱的诱导[28]。基于小麦为异源六倍体的特征以及植物中PLC基因多是以多基因家族形式存在,小麦基因组中应该还有其他的TaPLC基因序列。【拟解决的关键问题】本研究通过全面检索小麦基因组,获得全部TaPLC基因序列,明确其结构、表达模式和进化特征,并检测TaPLC基因在各种组织中以及在干旱或盐胁迫下的表达模式,为全面探究TaPLC基因家族成员的生物学功能奠定基础。1 材料与方法
1.1 植物材料
所用小麦品种有中国春,洛旱7#(LD)为抗旱小麦品种,科农199(KD)为对照小麦品种,小偃60#(XS)为耐盐小麦品种,石麦15#(SS)为对照小麦品种。选取小麦两叶一心期的根、叶以及成熟后的茎、穗等材料,液氮速冻后,-80℃保存备用。
小麦在霍氏培养液水培至两叶一心期时,分别移至20% PEG 6000和200 mmol·L-1 NaCl的培养液中,于0、0.5、1、2、6、12、24和48 h取叶片,液氮速冻后,-80℃保存备用。
1.2 TaPLC基因的鉴定
首先利用已知的水稻和拟南芥的PLC蛋白序列在全基因组数据库(Ensembl Plants)中查询小麦中的PLC同源基因的序列信息(守域的序列被认定为小麦TaPLC基因序列。
1.3 基因的染色体定位及序列分析
利用Ensembl Plants网站查询TaPLC基因在染色体上的定位;利用在线工具ExPASy(1.4 多序列比对及进化分析
利用DNAMAN软件对TaPLC蛋白序列进行多序列比对。为了研究不同物种PLC之间的进化关系,从NCBI下载拟南芥、水稻、大豆等物种PLC氨基酸序列,用Clustal W进行多序列比对,然后用MEGA 7.0采用邻接法构建系统发育树,Bootstrap值设置为1 000。利用Evolview在线软件对进化树进行优化处理。1.5 基因的表达分析
利用在线软件WoLF PSORT(Table 1
表1
表1小麦TaPLC基因家族各成员的分子和生化特征
Table 1
| 基因名称 Gene name | 基因ID Gene ID | 染色体定位a Chromosomal location | 开放阅读框 Open reading frame (bp) | 蛋白质b Protein | ||
|---|---|---|---|---|---|---|
| 大小 Size (aa) | 分子量 MW (kD) | 等电点 pI | ||||
| TaPLC1A | TraesCS1A02G069300 | 1A:51700021—51707185 | 1758 | 585 | 66.01 | 6.07 |
| TaPLC1D | TraesCS1D02G071800 | 1D:52338417—52345805 | 1761 | 586 | 66.19 | 6.03 |
| TaPLC2A | TraesCS2A02G084000 | 2A:38534860—38538480 | 1830 | 609 | 68.5 | 5.91 |
| TaPLC2B | TraesCS2B02G098500 | 2B:58321242—58325230 | 1821 | 606 | 68.22 | 6.06 |
| TaPLC2D | TraesCS2D02G082000 | 2D:35259471—35263395 | 1824 | 607 | 68.45 | 5.88 |
| TaPLC3A | TraesCS4A02G109000 | 4A:129086595—129090166 | 1902 | 633 | 71.11 | 5.54 |
| TaPLC3B | TraesCS4B02G195200 | 4B:420197892—420201623 | 1902 | 633 | 71.00 | 5.75 |
| TaPLC3D | TraesCS4D02G195800 | 4D:340085748—340090648 | 1902 | 633 | 71.05 | 5.84 |
| TaPLC4A | TraesCS5A02G155300 | 5A:333407514—333413175 | 1773 | 590 | 65.73 | 6.05 |
| TaPLC4B | TraesCS5B02G153600 | 5B:283008744—283014326 | 1770 | 589 | 65.65 | 6.06 |
| TaPLC4D | TraesCS5D02G160300 | 5D:250061407—250066699 | 1770 | 589 | 65.65 | 6.06 |
新窗口打开|下载CSV
利用qRT-PCR的方法检测和验证部分TaPLC基因的表达。首先利用检索到的TaPLC基因序列信息,以cDNA为模板扩增基因全长,并测序,进而根据序列信息设计区别于其他旁系同源基因的特异性定量引物(电子附表2)。
Table 2
表2
表2TaPLC基因表达蛋白的亚细胞分布特征
Table 2
| 基因Gene | 亚细胞定位Subcellular localization |
|---|---|
| TaPLC1A | 线粒体:8,叶绿体:4,细胞核:1 Mito: 8, chlo: 4, nucl: 1 |
| TaPLC1D | 线粒体:8,叶绿体:4,细胞核:1 Mito: 8, chlo: 4, nucl: 1 |
| TaPLC2A | 叶绿体:7,细胞核:3,线粒体:2.5,线粒体基质:2 Chlo: 7, nucl: 3, mito: 2.5, cyto_mito: 2 |
| TaPLC2B | 叶绿体:6,线粒体:5.5,线粒体基质:3.5,细胞核:1 Chlo: 6, mito: 5.5, cyto_mito: 3.5, nucl: 1 |
| TaPLC2D | 叶绿体:7,线粒体:4.5,线粒体基质:3,细胞核:1 Chlo: 7, mito: 4.5, cyto_mito: 3, nucl: 1 |
| TaPLC3A | 细胞质:8,细胞核:4,叶绿体:1 Cyto: 8, nucl: 4, chlo: 1 |
| TaPLC3B | 细胞质:8,细胞核:3,叶绿体:1,线粒体:1 Cyto: 8, nucl: 3, chlo: 1, mito: 1 |
| TaPLC3D | 细胞质:6,细胞核:3,线粒体:2,过氧化物:2 Cyto: 6, nucl: 3, mito: 2, pero: 2 |
| TaPLC4A | 线粒体:10,叶绿体和线粒体:6.83333,线粒体基质:5.83333,叶绿体:2.5 Mito: 10, chlo_mito: 6.83333, cyto_mito: 5.83333, chlo: 2.5 |
| TaPLC4B | 线粒体:10,叶绿体和线粒体:6.83333,线粒体基质:5.83333,叶绿体:2.5 Mito: 10, chlo_mito: 6.83333, cyto_mito: 5.83333, chlo: 2.5 |
| TaPLC4D | 线粒体:10,叶绿体和线粒体:6.83333,线粒体基质:5.83333,叶绿体:2.5 Mito: 10, chlo_mito: 6.83333, cyto_mito: 5.83333, chlo: 2.5 |
新窗口打开|下载CSV
将所取材料研磨成粉末,提取总RNA,并反转录为cDNA,用于实时荧光定量PCR检测。以ACTIN为内参基因,qRT-PCR的反应体系为模板1/3 μL、2×AceQ qPCR SYBR Green Master Mix 10 μL、上下游引物各0.5 μL(10 μmol·L-1),ddH2O补至20 μL,每个样品进行3次技术重复。反应程序为95℃ 5 min;95℃ 10 s,60℃ 30 s,40个循环,绘制融解曲线,95℃ 15 s,60℃ 60 s,95℃ 15 s。利用2-ΔΔCT法分析数据,试验设计3个生物学重复。
2 结果
2.1 小麦TaPLC基因家族成员的鉴定
为了全面鉴定小麦TaPLC基因家族成员,利用拟南芥AtPLC基因和水稻OsPLC基因的编码蛋白序列信息对小麦基因组数据库进行局部BLAST比对,并进行关键字搜索和蛋白质域搜索。发现小麦基因组序列中有11个序列同源基因(电子附表1),分别分布在第1、2、4和5染色体(表1)。小麦基因组为AABBDD六倍体,这11个TaPLC基因中有4组直系同源基因,每组直系同源基因分别有A、B和D 3个部分同源基因,分别将其命名为TaPLC1A、TaPLC1D、TaPLC2A、TaPLC2B、TaPLC2D、TaPLC3A、TaPLC3B、TaPLC3D、TaPLC4A、TaPLC4B和TaPLC4D。未能在第1B染色体上检出同源序列,推测是由于亚基因组间的不对称进化所导致。为了进一步确定第1B染色体中TaPLC基因丢失的时间,利用TaPLC1A和TaPLC1D序列分别检索了AA(Triticum urartu)二倍体、DD(Aegilops tauschii)二倍体和AABB(Triticum dicoccoides)四倍体小麦祖先种基因组。结果显示,在祖先种的AA和DD基因组中均检索到高度相似同源基因(TRIUR3_33781-T和AET1Gv20175500.1),但在AABB基因组中只在A亚组发现同源基因(TRIDC1AG009650.1),而没有检索到B亚组上的同源基因,揭示B亚基因组中的TaPLC基因在异源六倍体形成之前就已经缺失(电子附图1),暗示了小麦亚基因组间进化的不对称性。
图1
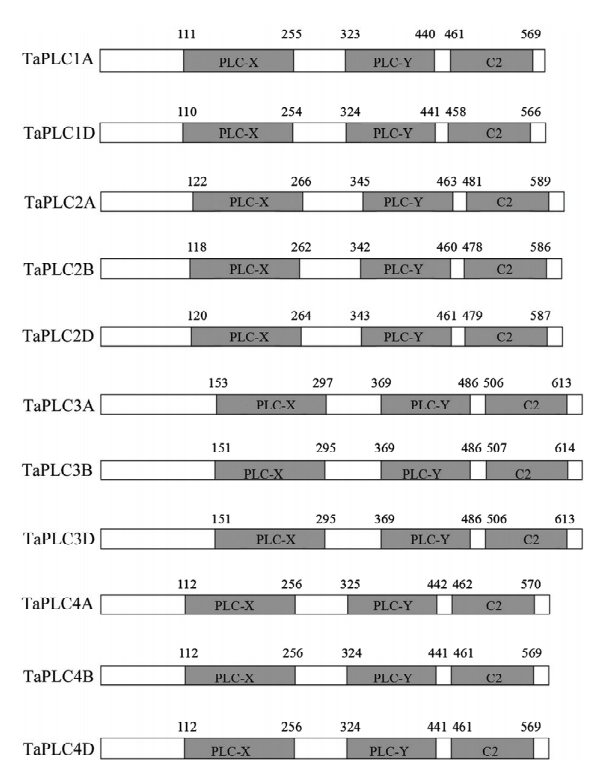 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1小麦TaPLC蛋白质的功能结构域分析
数字表示每个结构域在蛋白质序列中的位置
Fig. 1Functional domain analysis of wheat TaPLC protein
Numbers indicating positions of each domain in protein sequences
根据Pfam、SMART和CDD的结果,获得的TaPLC基因编码序列都具有X、Y和C2结构域,并用IBS绘制TaPLC蛋白结构域示意图(图1)。通过分析TaPLC基因的分子和生化特性,发现这些基因的ORF长度范围为1 770—1 902 bp,其编码589—633 aa多肽,预测分子量范围为5.54—71.11 kD。理论等电点为5.37—6.07(表1)。
2.2 小麦TaPLC基因的系统发育分析
将检索到的TaPLC蛋白序列与目前已知的植物PLC蛋白序列进行比对和分析,包括9个拟南芥AtPLC基因、4个水稻OsPLC基因和6个番茄SlPLC基因等共15种植物的56个PLC蛋白序列,7种苔藓PLC、2种酵母PLC、2种微藻和3种动物ζ亚型的PLC等共计67个蛋白序列,结果显示,小麦TaPLC基因家族与已知PLC序列有很高的相似性,其中TaPLC2组(即TaPLC2A、TaPLC2B和TaPLC2D)、TaPLC3组和TaPLC4组同源关系较近,它们与水稻、玉米、百合和高粱等单子叶植物的PLC的相似性较高,但TaPLC1组的2个蛋白(TaPLC1A和TaPLC1D)则与双子叶植物(如拟南芥、大豆和番茄等)具有较高的相似度(图2)。结合TaPLC1在B基因组中丢失的现象,说明小麦中的TaPLC基因在进化进程中既具有保守性,也出现了分歧。图2
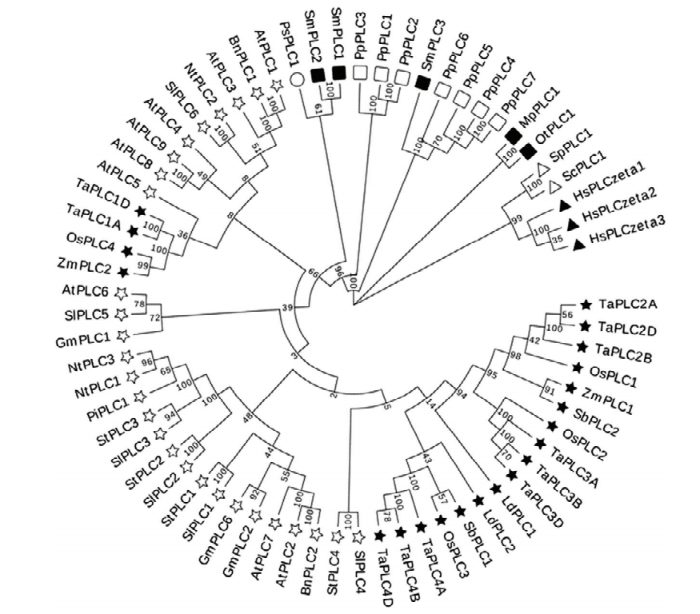 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2PLC基因家族蛋白的系统发育分析
☆:双子叶植物;★:单子叶植物;○:松柏类植物;●:石松类植物;□:苔藓植物;■:微藻;△:酵母;▲:哺乳动物。Ta:小麦;At:拟南芥;Gm:大豆;Os:水稻;Pp:小立碗藓;Ps:云杉;Nt:烟草;Sl:番茄;Ld:百合;Sb:高粱;Pi:矮牵牛;St:马铃薯;Bn:油菜;Sm:卷柏;Mp:细小微胞藻;Ot:金牛鸵球藻;Hs:智人;Sp:裂殖酵母;Sc:酿酒酵母;Zm:玉米
Fig. 2Phylogenetic analyses of PLC gene family proteins
☆: Dicots; ★: Monocots; ○: Conifer; ●: Lycophytes; □: Bryphytes; ■: Algae; △: Yeast; ▲: Mammals. Ta: Triticum aestivum; At: Arabidopsis thaliana; Gm: Glycine max; Os: Oryza sativa; Pp: Physcomitrella patens; Ps: Picea sitchensis; N: Nicotiana tabacum; Sl: Solanum licopersicum; Ld: Lilium davidii; Sb: Sorghum bicolor; Pi: Petunia integrifolia; St: Solanum tuberosum; Bn: Brassica napus; Sm: Selaginella moellendorffii; Mp: Micromonas pusilla; Ot: Ostreococcus tauri; Hs: Homo sapiens; Sp: Schizosaccharomyces pombe; Sc: Saccharomyces cerevisiae; Zm: Zea mays
2.3 小麦TaPLC基因的结构特征
为了进一步了解TaPLC基因的发育关系,分析比较TaPLC基因的DNA序列和氨基酸序列的结构,及其外显子-内含子结构。发现同一组的基因(如TaPLC2A/TaPLC2B/TaPLC2D)彼此之间不仅核酸序列和氨基酸序列相似性都达到98%以上(电子附表1),而且外显子-内含子结构也具有高度同一性(图3),表明小麦TaPLC基因每组的部分同源基因是经历了基因复制而形成的。TaPLC2组的3个基因均有8个外显子和7个内含子,其他组的基因均有9个外显子和8个内含子,与目前发现的其他植物的PLC基因有着相似的剪接方式。图3
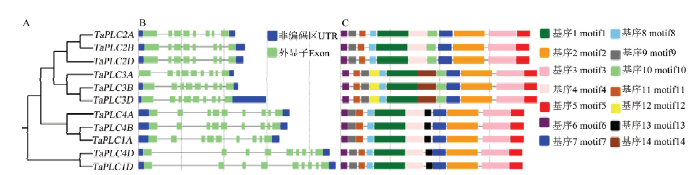 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3TaPLC基因的系统发育关系、基因结构和基序
A:TaPLC基因系统发育树;B:TaPLC基因外显子-内含子结构示意图;C:TaPLC蛋白基序示意图,基序的序列和logo图见电子附
Fig. 3Phylogenetic relationship, gene structure and motifs of the TaPLC genes
A: The phylogenetic tree of TaPLC genes; B: Schematic diagram for the exon-intron organization of TaPLC基因; C: Schematic diagram for the motifs of TaPLC genes. The motif's sequence and logo diagrams are in
利用MEME,在TaPLC基因家族中找到了14个不同的motif(图3-C)。亲缘关系较近的成员具有相似的motif组合。TaPLC基因共同具有10个motif,TaPLC1A/TaPLC1D特有motif 13,TaPLC3A/TaPLC3B/ TaPLC3D特有motif 12,TaPLC4A/TaPLC4B/TaPLC4D特有motif 13。14个motif的图示和序列在电子附表3和电子附图2中列出。
2.4 TaPLC基因启动子的顺式作用元件
为了探究能够影响TaPLC基因表达因素及TaPLC基因可能参与的调控途径,利用在线软件(PlantCARE和PLACE)分析了翻译起始位点(ATG)上游的1.5 kb区域的调控序列。结果显示,TaPLC基因都含有多种顺式作用元件,除了2个核心顺式元件T和C(分别是TATA box和CAAT box),还发现了6个植物激素响应元件,包括ABA响应元件、乙烯响应元件、2个GA响应元件及2个生长素响应元件;同时发现8个应激响应元件,即干旱响应元件、低温响应元件,热激响应元件和光响应元件等(图4和电子附表4),暗示TaPLC基因可能参与了植物对激素和逆境胁迫的响应过程。此外,一些部分同源基因之间,如TaPLC2A、TaPLC2B和TaPLC2D之间,有着类似的顺式调控元件;而不同组的基因之间却有较大的差异,如TaPLC2组的元件数量,较TaPLC4组的多(图4),暗示小麦中不同的TaPLC基因,有些因受共同的因素调控而呈现相近表达模式的基因,也有的调控和表达方式彼此差异较大,从而能够在多种的生理过程中发挥作用。图4
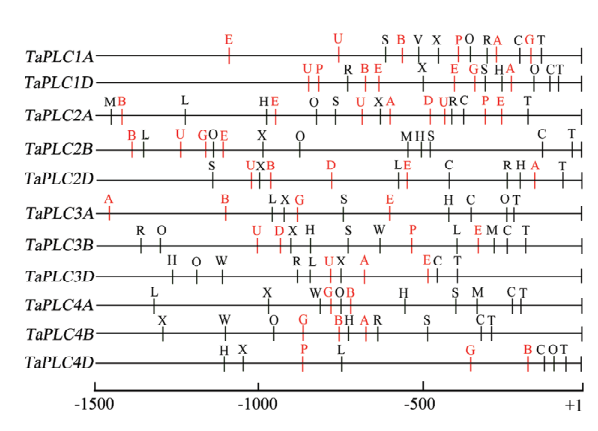 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4TaPLC基因启动子顺式元件调控分析
A:ABA响应元件;E:乙烯反应元件;G:赤霉素反应元件;P:赤霉素反应元件;B:参与茉莉酸反应的元件;D和U:生长素反应元件;W:伤害反应元件;L:低温响应元件;S:干旱反应相关的MYB结合位点;M:光反应相关的MYB结合位点;H:热激反应元件;R:光反应的顺式调控元件;O:光反应元件的一部分;X:应答病菌反应的WRKY蛋白结合位点。激素反应顺式元件为红色字母;胁迫反应顺式元件为黑色字母
Fig. 4Putative regulatory cis-elements in the TaPLC genes promoters of wheat
A: Abscisic acid (ABA)-responsive element; E: Ethylene-responsive element; G: Gibberellin-responsive element; P: Gibberellin-responsive element; B: Element involved in the MeJA-responsiveness; D and U: Auxin-responsive element; W: Wound-responsive element; L: Element involved in low-temperature responsiveness; S: MYB binding site involved in drought-inducibility; M: MYB binding site involved in light responsiveness; H: Heat shock-responsive element; R: Element involved in light responsiveness; O: Part of a light responsive element; X: WRKY protein-specific binding site, responsive to pathogen infection. The hormone response cis-elements are red letters; The stress response cis-elements are black letters
2.5 小麦TaPLC基因在细胞、组织中的表达模式
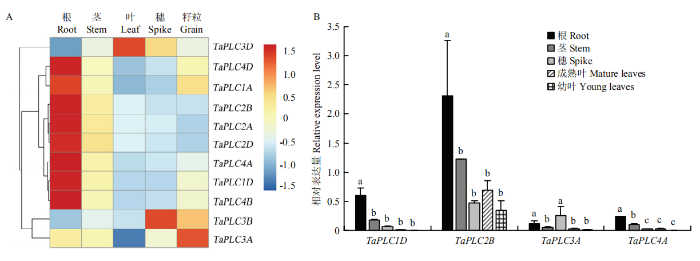
利用WoLF POSRT分析结果显示,不同的TaPLC基因具有不同的亚细胞分布特征,其中TaPLC1组和TaPLC4组分别主要在线粒体中分布,TaPLC2组主要在叶绿体中分布,而TaPLC3组主要在细胞质中分布(表2)。为了更好地揭示TaPLC基因在异源六倍体小麦中的潜在功能,在2种分析网站上检测了该家族在小麦各个组织中的表达丰度。分析结果彼此相似,数据显示,TaPLC1(A/D)、TaPLC2(A/B/D)和TaPLC4(A/B/D)均在根中高表达,而TaPLC3A、TaPLC3B和TaPLC3D则分别在穗、籽粒和叶中有较高丰度(图5-A和电子附图3)。
图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5TaPLC基因家族组织水平表达模式
A:RNA-seq数据分析;B:qRT-PCR检测结果。小写字母表示各基因在不同组织的显著性水平(P = 0.05)
Fig. 5Tissue expression analysis of TaPLC gene
A: RNA-seq data analysis results; B: qRT-PCR results. Lowercase letter indicates significance level (P=0.05) of each gene at different tissues
为了进一步验证转录组的结果,根据获得的TaPLC1D、TaPLC2B、TaPLC3A和TaPLC4A cDNA序列设计引物(电子附表2),用实时荧光定量PCR方法分析这些基因在不同组织的表达情况。结果显示,4个基因在5种组织(根、茎、穗、成熟叶和幼叶)中均有表达,但表达水平彼此不同。TaPLC2B在所检测的组织中均为表达最高的,TaPLC4A表达最低;TaPLC1D、TaPLC2B和TaPLC4A均在根中表达最高,TaPLC3A则是在穗中表达最高。qRT-PCR结果与转录组数据具有一定的相似性,如TaPLC3在穗中表达最高,其他基因均在根中表达最高(图5)。以上结果表明TaPLC基因在不同组织的表达模式具有相似性,但也存在差异,暗示该基因家族在小麦的生长发育过程中发挥着作用,同时也暗示不同成员之间可能发生了一定程度的功能分化。
2.6 非生物胁迫对TaPLC基因表达的影响
基于膜磷脂代谢的肌醇磷脂信号系统,在植物应答渗透胁迫的反应中发挥着重要的作用。检测了TaPLC基因在盐或干旱胁迫下的表达变化。结果显示,所检测TaPLC基因的转录均显著受到盐或干旱的诱导(图6)。干旱刺激0.5 h后,4个TaPLC基因的表达量均提高,上调最显著的是TaPLC1D和TaPLC2B;其中TaPLC2B和TaPLC3A在干旱6 h时表达量升至最高,而TaPLC1D和TaPLC4A,则在干旱12 h时达到高峰(图6-A),暗示TaPLC基因在参与植物对干旱胁迫响应的过程中受到了不同的调控策略。同样,该家族成员也受到盐胁迫的诱导,但对盐胁迫的敏感性也存在着一定的差异,如TaPLC1D、TaPLC1D2B和TaPLC1D3A均在胁迫处理6 h时表达量最高,而TaPLC4A则在受刺激后2 h即达到高峰(图6-B)。图6
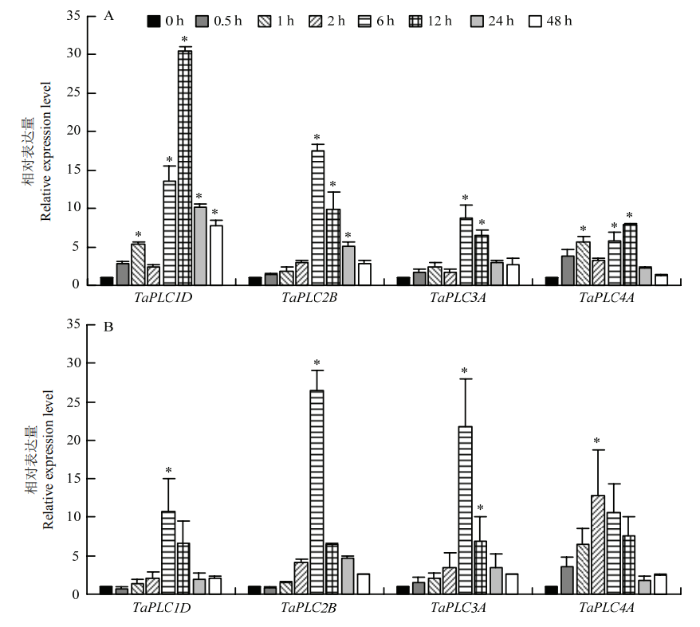 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6TaPLC基因盐和干旱条件下表达分析
A:TaPLC基因干旱条件下表达分析;B:TaPLC基因盐条件下表达分析。星号表示与对照组比较差异显著(P= 0.05)
Fig. 6Analysis of TaPLC gene expression under salt and drought condition
A: Analysis of TaPLC gene expression under drought condition; B: Analysis of TaPLC gene expression under salt condition. Asterisk indicate the significant difference (P = 0.05) compared with control
为了解TaPLC基因受盐或干旱诱导表达是否具有普适性,选择抗旱小麦品种洛旱7#(LD)[29](以科农199(KD)为对照)和耐盐品种小偃60#(XS)[30](以石麦15#(SS)为对照)检测了TaPLC基因在胁迫下的表达特征。结果显示,无论是耐旱/盐的品种,还是相对不耐旱/盐的对照品种,TaPLC基因的表达均显著受到干旱/盐胁迫的诱导;另外,干旱胁迫下,抗旱的品种LD中,TaPLC2B表达较对照KD更高;耐盐的品种中,TaPLC1D等在盐的胁迫下也比对照有更高的表达量(图7)。
图7
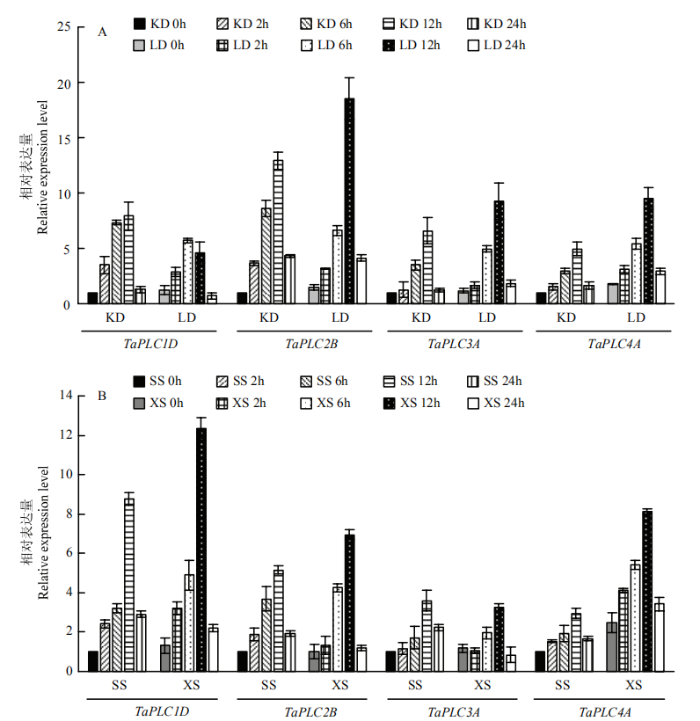 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7耐旱和耐盐品种中TaPLC基因盐和干旱条件下表达分析
A:耐旱品种中TaPLC基因在干旱条件下表达分析;B:耐盐品种中TaPLC基因在盐条件下表达分析。KD:科农199;LD:洛旱7#; SS:石麦15#;XS:小偃60#
Fig. 7Analysis of TaPLC genes expression in drought and salt tolerance varieties under drought and salt condition
A: Analysis of TaPLC genes expression in drought tolerance varieties under drought condition; B: Analysis of TaPLCs gene expression in salt tolerance varieties under salt condition. KD: Kenong 199; LD: Luohan 7#; SS: Shimai 15#; XS: Xiaoyan 60#
综上所述,TaPLC基因受到干旱和盐胁迫的诱导,且各基因对不同胁迫处理的敏感性存在差异,揭示家族成员在响应非生物胁迫过程存在着一定的功能分化。
3 讨论
3.1 小麦中的TaPLC基因家族
通过对小麦基因数据库的检索,在中国春小麦基因组中发现11个PI-PLC编码基因。生物信息学分析结果表明,该家族成员的基因结构、编码蛋白序列的结构以及生化特征等参数与目前已知的植物PLC基因相似[31],显示了TaPLC基因序列在进化上的保守性。目前发现植物中PLC基因都是以多成员的家族形式存在,如拟南芥中9个成员、水稻中4个、番茄中6个等[31,32,33]。由于小麦异源六倍体的遗传特征,11个TaPLC基因根据其序列的同源性和结构特征,可以分成4组直系同源基因,即TaPLC1—TaPLC4,每组内包括3个分别位于A、B和D亚基因组中的部分同源基因,其序列相似性达到98%—99%(电子附表1)。另外,TaPLC1在B染色体上却没有拷贝,进一步检索AA二倍体,DD二倍体和AABB四倍体小麦祖先种基因组,发现B基因组中的TaPLC1在异源六倍形成之前就已经缺失(电子附图1)。TaPLC3组内的3个基因在4A、4B和4D染色体上的位置不尽相同。这些结果说明在小麦的进化过程中染色体可能丢失部分基因或发生了基因易位(表1)。通过系统进化分析发现,不同物种的PLC基因均存在于不同的分支中,即使在植物中,单子叶和双子叶的PLC也明显地被区分在不同的模块中,显示出PLC在不同物种中存在着进化上的分歧。然而,小麦TaPLC1A和TaPLC1D,与单子叶植物水稻OsPLC4和玉米ZmPLC2序列的相似度却比双子叶植物的PLC基因序列相似性更高些,表明小麦的TaPLC1A和TaPLC1D,在进化上与其他的旁系同源基因有较大的差异(图2)。另一方面,根据外显子-内含子结构进行的聚类分析,TaPLC1组的2个基因,均有较长的第一个内含子,与其他3组TaPLC基因的相似度最低,反而与拟南芥AtPLC2和AtPLC4,以及水稻OsPLC4的结构相似[31,33]。外显子-内含子结构是基因演变的重要参数。因此这一组的基因同源关系较其他组较远,这与图3显示的进化树的结果也是一致的。
3.2 TaPLC基因的表达模式
植物PLC的研究最早始于小麦,早在1987、1992年人们分别从小麦中检测到了PLC的生物活性[25,26,27],河北省植物生理与分子病理学重点实验室前期根据文献报道的2个小麦TaPLC1和TaPLC2(GenBank:HM754654.1和HM754653.1)[34]序列进行了药物学和表达分析检测,发现高盐或干旱至少诱导了TaPLC1的转录和翻译水平的表达,并且施加PLC抑制剂U73122或edelfosine会损害幼苗生长并增强幼苗对干旱和高盐分胁迫的敏感性,表明TaPLC1参与了小麦对盐或干旱胁迫的应答反应[28]。干旱和高盐导致的渗透胁迫是影响植物的生长发育,造成世界范围内的作物减产的重要因素。固着生长的植物进化出复杂的信号传导系统,以感受环境变化,调控基因表达,从而在细胞和分子水平上对环境胁迫做出响应[1,35]。磷酸肌醇信号传导途径在植物对渗透胁迫的响应中具有重要作用[4]。作为该途径的关键酶之一,植物PLC不仅在渗透胁迫下被诱导表达,而且它们的过表达植物在胁迫环境下还表现多种抵抗或耐受胁迫的表型。拟南芥有9个AtPLC家族成员,第一个被克隆的AtPLC1s就是受干旱或盐强烈诱导的亚型[36];qRT-PCR检测结果显示,盐胁迫下AtPLC4、AtPLC5和AtPLC7 3种亚型的转录水平提高最多,达5倍以上[37,38];遗传学试验也证实,AtPLC4负调节拟南芥的耐盐性,AtPLC5和AtPLC7的超表达均提高了拟南芥的耐旱性能[18,21-23]。玉米ZmPLC1主要在根中表达,干旱胁迫可以诱导其表达上升[36],超表达ZmPLC1转基因玉米,比野生型玉米表现出更好的耐旱性能[11]。NaCl处理后,水稻叶片中的PLC的底物分子PIP2的含量会在30 min内迅速增加至4倍[40];水稻的4种OsPLC基因中,OsPLC1在盐胁迫下表达上调,OsPLC4的表达则降低[22];OsPLC1介导Ca2+信号产生,建立了整个植株的耐盐性[8];两个敲除突变体osplc4-1和osplc4-2株系的幼苗在盐胁迫下的生长和成活率都降低,过表达OsPLC4可以提高高盐和缺水条件下的成活率[41]。另外,其他物种如烟草NtPLC基因、马铃薯StPLC基因和绿豆VrPLC基因等被克隆时,也检测到其表达受到盐、干旱等刺激诱导[8,10,13-14]。甘蓝型油菜BnPLC2的过量表达,增强了植物的抗旱性并促进早花和成熟[42]。以上表明,表达受到胁迫影响的PLC基因,在植物体内超表达或缺失后,都会改变植物抵抗或耐受盐或干旱刺激的生理性状。
本研究检索小麦基因组序列,获得了11个TaPLC基因序列后,检测了它们在高盐或干旱胁迫下的表达变化,发现所检测的4个TaPLC基因在转录水平都显著受到胁迫刺激的诱导,其中TaPLC1D在干旱胁迫下表达上升最高,而TaPLC2B受盐胁迫诱导最显著(图6)。通过序列比对,核实了TaPLC2B即前述报道的TaPLC1(GenBank:HM754654.1),TaPLC1D即TaPLC2(GenBank:HM754653.1)。另外有趣的是,这些TaPLC基因在耐旱/盐的小麦品种洛旱7#(LD)/小偃60#(XS)[29,30]以及不耐的对照品种中,表达均显著受到干旱/盐胁迫的诱导(图7)。综上,植物PLC以表达量变化的方式普遍参与的盐或干旱的应答响应。植物面对高盐或干旱甚至冷、热等渗透胁迫的物理性因子刺激时,一是可以在细胞的各个位置通过各种大分子和结构独立地感知,此外植物在应答这些胁迫信号时具有一些共同的特征,其中最主要的就是Ca2+作为主要应答的通用第二信使[1,4]。而植物PLC,作为调节膜脂分子代谢的酶类,不仅依附于细胞质膜存在,还在线粒体、叶绿体等细胞器中分布(表2),并且是钙信号形成的上游调控者[3,4]。结合TaPLC基因在不同品系的小麦中受盐或干旱诱导后的表达模式的相似性,推测PLC基因作为膜肌醇磷脂信号转导途径的关键因子,其部分功能可能是参与植物对胁迫信号的感知。另外,在TaPLC基因的调控序列区,除了共有的TATA box和CAAT box外,还有ABA、乙烯、赤霉素、生长素等激素反应的顺式作用元件,光反应相关元件以及低温、热激和干旱等渗透反应的响应元件(图4)。非生物胁迫触发的信号途径与激素对植物的调节过程有很多的cross talk[1,43]。以上诸多信息对于深入研究TaPLC基因家族的每个成员参与小麦应答盐或干旱生理过程的作用机制提供了十分有用的信息,为利用这些基因对小麦进行遗传改良奠定好的基础。
4 结论
TaPLC基因家族中有11个植物PLC直系同源序列,分为4组,每组内的3个(或2个)部分同源物为小麦六倍体演化过程中染色体复制形成的拷贝。不同的TaPLC基因有各自的组织和亚细胞水平的分布特征,并且每组的TaPLC基因成员至少有1个基因的转录受到盐或干旱的胁迫诱导。TaPLC基因的上游调控区存在激素调控的顺式元件及应激响应元件。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1007/s11427-020-1683-xURLPMID:32246404 [本文引用: 5]

Abiotic stresses and soil nutrient limitations are major environmental conditions that reduce plant growth, productivity and quality. Plants have evolved mechanisms to perceive these environmental challenges, transmit the stress signals within cells as well as between cells and tissues, and make appropriate adjustments in their growth and development in order to survive and reproduce. In recent years, significant progress has been made on many fronts of the stress signaling research, particularly in understanding the downstream signaling events that culminate at the activation of stress- and nutrient limitation-responsive genes, cellular ion homeostasis, and growth adjustment. However, the revelation of the early events of stress signaling, particularly the identification of primary stress sensors, still lags behind. In this review, we summarize recent work on the genetic and molecular mechanisms of plant abiotic stress and nutrient limitation sensing and signaling and discuss new directions for future studies.
[本文引用: 2]
[本文引用: 2]
DOI:10.1111/pce.12666URLPMID:26510494 [本文引用: 5]
Lipids are one of the major components of biological membranes including the plasma membrane, which is the interface between the cell and the environment. It has become clear that membrane lipids also serve as substrates for the generation of numerous signalling lipids such as phosphatidic acid, phosphoinositides, sphingolipids, lysophospholipids, oxylipins, N-acylethanolamines, free fatty acids and others. The enzymatic production and metabolism of these signalling molecules are tightly regulated and can rapidly be activated upon abiotic stress signals. Abiotic stress like water deficit and temperature stress triggers lipid-dependent signalling cascades, which control the expression of gene clusters and activate plant adaptation processes. Signalling lipids are able to recruit protein targets transiently to the membrane and thus affect conformation and activity of intracellular proteins and metabolites. In plants, knowledge is still scarce of lipid signalling targets and their physiological consequences. This review focuses on the generation of signalling lipids and their involvement in response to abiotic stress. We describe lipid-binding proteins in the context of changing environmental conditions and compare different approaches to determine lipid-protein interactions, crucial for deciphering the signalling cascades.
DOI:10.1111/j.1365-3040.2009.02097.xURLPMID:20429089 [本文引用: 5]
Polyphosphoinositides (PPIs) became famous for their role in inositol-1,4,5-trisphosphate (InsP3) mediated-Ca(2+) signalling in mammalian cells, generated through signal-activated phospholipase C (PLC) hydrolysis of the minor membrane lipid, phosphatidylinositol-4,5-bisphosphate. For many years, the plant field followed the same paradigm, however, slowly a completely different picture is emerging. Moreover, various novel PPI-signalling compounds have been identified meanwhile, with new functions and targets coming to light. These include lipids phosphorylated at the D3-position of inositol but also water-soluble inositolpolyphosphates (IPPs). For several of them, a relationship to water stress has been reported. This review summarizes the current status of PPIs and IPPs in plants and discusses their potential in osmotic stress signalling and drought.
DOI:10.1016/j.plaphy.2008.04.015URLPMID:18534862 [本文引用: 1]
Phosphoinositide-specific phospholipase C cleaves the substrate phosphatidylinositol 4,5-bisphosphate and generates inositol 1,4,5-trisphosphate and 1,2-diacylglycerol, both of which are second messengers in the phosphoinositide signal transduction pathways operative in animal cells. Five PI-PLC isoforms, beta, gamma, delta, epsilon and zeta, have been identified in mammals. Plant PI-PLCs are structurally close to the mammalian PI-PLC-zeta isoform. The Arabidopsis genome contains nine AtPLC genes. Expression patterns of all nine genes in different organs and in response to various environmental stimuli were studied by applying a quantitative RT-PCR approach. Multiple members of the gene family were differentially expressed in Arabidopsis organs, suggesting putative roles for this enzyme in plant development, including tissue and organ differentiation. This study also shows that a majority of the AtPLC genes are induced in response to various environmental stimuli, including cold, salt, nutrients Murashige-Skoog salts, dehydration, and the plant hormone abscisic acid. Results of this and previous studies strongly suggest that transcriptional activation of the PI-PLC gene family is important for adapting plants to stress environments. Expression patterns and phylogenetic relationships indicates that AtPLC gene members probably evolved through multiple rounds of gene duplication events, with AtPLC4 and AtPLC5 and AtPLC8 and AtPLC9 being duplicated in tandem in recent times.
DOI:10.1046/j.1365-313x.1995.08030381.xURLPMID:7550376 [本文引用: 1]
Phosphoinositide-specific phospholipase C (PI-PLC) is a key signal transducing enzyme which generates the second messengers inositol trisphosphate and diacylglycerol in mammalian cells. A cDNA clone (PI-PLC1) encoding a phosphoinositide-specific phospholipase C was isolated from soybean by screening a cDNA expression library using an anti-(plasma membrane) serum. Genomic DNA gel blot analysis suggested that the corresponding gene is a member of a multigene family. The deduced amino acid sequence of the soybean PI-PLC1 isozyme contains the conserved X and Y regions, found in other PI-PLCs. It is closely related to mammalian delta-type PI-PLCs, Dictyostelium discoideum PI-PLC and yeast PI-PLC1 in terms of the arrangement of the conserved region. Unlike mammalian delta-type PI-PLCs and yeast PI-PLC1, the putative Ca(2+)-binding site of the soybean PI-PLC1 is located in the region spanning the X and Y domains, and the N-terminal region is truncated. FLAG epitope-tagged PI-PLC1 fusion protein purified from transgenic tobacco plants showed phosphoinositide-specific phospholipase C activity. Heterologous expression of the soybean PI-PLC1 cDNA in a yeast PI-PLC1 deletion mutant complemented the lethality phenotype of haploid PI-PLC1 disruptants. Immunoblot analysis of the cell fractions prepared from transgenic tobacco plants over-expressing the FLAG epitope-tagged PI-PLC1 fusion protein indicated that the protein encoded by the PI-PLC1 cDNA was localized in the cytosol and plasma membrane.
DOI:10.1104/pp.114.2.747URLPMID:9235602 [本文引用: 1]
DOI:10.1007/s11105-011-0360-zURL [本文引用: 3]
The role of plant phospholipase C-mediated signaling has been implicated in various phases of plant growth and development. In this study, we report on the isolation and characterization of phospholipase C from tobacco and demonstrate that transcripts of phospholipase C are up-regulated in responses to drought and salt stress. These responses are likely by abscisic acid (ABA). Transgenic tobacco plants overexpressing the phospholipase C protein were found to tolerate higher levels of drought and also salinity stress. This tolerance could be mediated by the regulation of genes downstream to phospholipase mediated signaling. As a demonstration, when tested the transgenic plants showed higher transcript of heat shock factor NtHSF2, heat shock protein HSP70-3 and an AP2 domain transcription factor. Also the transgenic plants showed higher accumulation of sodium in older leaves compared to the young leaves. The present report is the first to demonstrate the role of phospholipase C in salinity stress tolerance.
DOI:10.1111/nph.14426URL [本文引用: 1]
DOI:10.1104/pp.116.1.239URLPMID:9449844 [本文引用: 2]
Many cellular responses to stimulation of cell-surface receptors by extracellular signals are transmitted across the plasma membrane by hydrolysis of phosphatidylinositol-4,5-bisphosphate (PIP2), which is cleaved into diacylglycerol and inositol-1,4,5-trisphosphate by phosphoinositide-specific phospholipase C (PI-PLC). We present structural, biochemical, and RNA expression data for three distinct PI-PLC isoforms, StPLC1, StPLC2, and StPLC3, which were cloned from a guard cell-enriched tissue preparation of potato (Solanum tuberosum) leaves. All three enzymes contain the catalytic X and Y domains, as well as C2-like domains also present in all PI-PLCs. Analysis of the reaction products obtained from PIP2 hydrolysis unequivocally identified these enzymes as genuine PI-PLC isoforms. Recombinant StPLCs showed an optimal PIP2-hydrolyzing activity at 10 microM Ca2+ and were inhibited by Al3+ in equimolar amounts. In contrast to PI-PLC activity in plant plasma membranes, however, recombinant enzymes could not be activated by Mg2+. All three stplc genes are expressed in various tissues of potato, including leaves, flowers, tubers, and roots, and are affected by drought stress in a gene-specific manner.
DOI:10.1007/s00425-007-0686-9URL [本文引用: 2]
Phosphatidylinositol-specific phospholipase C (PI-PLC) plays an important role in a variety of physiological processes in plants, including drought tolerance. It has been reported that the ZmPLC1 gene cloned from maize (Zea mays L.) encoded a PI-PLC and up-regulated the expression in maize roots under dehydration conditions (Zhai SM, Sui ZH, Yang AF, Zhang JR in Biotechnol Lett 27:799–804, 2005). In this paper, transgenic maize expressing ZmPLC1 transgenes in sense or antisense orientation were generated by Agrobacterium-mediated transformation and confirmed by Southern blot analysis. High-level expression of the transgene was confirmed by real-time RT-PCR and PI-PLC activity assay. The tolerance to drought stress (DS) of the homogenous transgenic maize plants was investigated at two developmental stages. The results demonstrated that, under DS conditions, the sense transgenic plants had higher relative water content, better osmotic adjustment, increased photosynthesis rates, lower percentage of ion leakage and less lipid membrane peroxidation, higher grain yield than the WT; whereas those expressing the antisense transgene exhibited inferior characters compared with the WT. It was concluded that enhanced expression of sense ZmPLC1 improved the drought tolerance of maize.
DOI:10.1093/pcp/pci181URLPMID:16085656 [本文引用: 1]
The phosphatidylinositol-specific phospholipase C (PI-PLC) activity is detected in purified Lilium pollen protoplasts. Two PI-PLC full length cDNAs, LdPLC1 and LdPLC2, were isolated from pollen of Lilium daviddi. The amino acid sequences for the two PI-PLCs deduced from the two cDNA sequences contain X, Y catalytic motifs and C2 domains. Blast analysis shows that LdPLCs have 60-65% identities to the PI-PLCs from other plant species. Both recombinant PI-PLCs proteins expressed in E. coli cells show the PIP(2)-hydrolyzing activity. The RT-PCR analysis shows that both of them are expressed in pollen grains, whereas expression level of LdPLC2 is induced in germinating pollen. The exogenous purified calmodulin (CaM) is able to stimulate the activity of the PI-PLC when it is added into the pollen protoplast medium, while anti-CaM antibody suppresses the stimulation effect caused by exogenous CaM. PI-PLC activity is enhanced by G protein agonist cholera toxin and decreased by G protein antagonist pertussis toxin. Increasing in PI-PLC activity caused by exogenous purified CaM is also inhibited by pertussis toxin. A PI-PLC inhibitor, U-73122, inhibited the stimulation of PI-PLC activity caused by cholera toxin and it also leads to the decrease of [Ca(2+)](cyt) in pollen grains. Those results suggest that the PPI-PLC signaling pathway is present in Lilium daviddi pollen, and PI-PLC activity might be regulated by a heterotrimeric G protein and extracellular CaM.
DOI:10.1016/S0014-5793(03)01388-7URL [本文引用: 2]
URLPMID:26155672 [本文引用: 2]
Phosphatidylinositol (PtdIns) is a major phospholipid in eukaryotic cells. Many studies have revealed that the phosphoinositide (PI) signaling pathway plays an important role in plant growth and development. Phospholipase C (PLC) is reported to have a crucial role in the PI pathway. This work focuses on the isolation and investigation of PLC in response to abiotic stress factors in green gram. The PLC cDNA, designated VrPLC, encoding a protein of 591 amino acids was cloned and expressed in E. coli. The predicted isoelectric point (pI) and molecular weight were 5.96 and 67.3 kDa, respectively. The tertiary structure of the PLC was also predicted and found to be mainly composed of random coils. In addition, VrPLC expression analysis was performed under environmental stress and the results showed that the expression of VrPLC was rapidly induced in an abscisic acid independent manner in response to drought and salt stress. PLC expression was found to be up-regulated by SA and down-regulated by wound in leaf tissues; however, there was no significant difference in the expression of PLC in plants subjected to high temperature and H2O2. Our results suggest that a close link/relationship between PLC expression and stress responses in green gram.
DOI:10.1016/j.pep.2006.10.007URL [本文引用: 1]
Abstract
Phosphoinositide-specific phospholipase Cs (PI-PLCs) are important enzymes in eukaryotes, which catalyze the hydrolysis of phosphatidylinositol 4,5-bisphosphate into the two second messengers inositol 1,4,5-trisphosphate and diacylglycerol. The Arabidopsis genome contains nine putative PI-PLC genes. AtPLC4, an abiotic stress induced gene, has been reported to encode an active PI-PLC isoform. However, the exact roles of putative AtPLC4 in plant remain to be elicited. The first 108 amino acid residues of the N-terminal of AtPLC4, referred to as AtPLC4 N, was expressed as a recombinant protein in Escherichia coli and used as antigen in generating antibody. Purified recombinant proteins including AtPLC1 to AtPLC5, AtPLC8, AtPLC9 and AtPLC4 N were transferred onto the same blot to test specificity of the prepared antibody. Western blot result shows that only AtPLC4 and AtPLC4 N can be recognized by the antibody. The antibody recognized a protein of approximately 68 kDa in the plasma membrane fraction and cytosolic fractions prepared from Arabidopsis thaliana plants. This corresponds very well with the calculated molecular weight of AtPLC4. The results suggest that AtPLC4 may encode a plasma membrane-associated protein.DOI:10.1111/pce.12918URLPMID:28102910 [本文引用: 1]
Previous physiological and pharmacological studies have suggested that the activity of phosphoinositide-specific phospholipase C (PI-PLC) plays an important role in regulating plant salt stress responses by altering the intracellular Ca(2+) concentration. However, the individual members of plant PLCs involved in this process need to be identified. Here, the function of AtPLC4 in the salt stress response of Arabidopsis seedlings was analysed. plc4 mutant seedlings showed hyposensitivity to salt stress compared with Col-0 wild-type seedlings, and the salt hyposensitive phenotype could be complemented by the expression of native promoter-controlled AtPLC4. Transgenic seedlings with AtPLC4 overexpression (AtPLC4 OE) exhibited a salt-hypersensitive phenotype, while transgenic seedlings with its inactive mutant expression (AtPLC4m OE) did not exhibit this phenotype. Using aequorin as a Ca(2+) indicator in plc4 mutant and AtPLC4 OE seedlings, AtPLC4 was shown to positively regulate the salt-induced Ca(2+) increase. The salt-hypersensitive phenotype of AtPLC4 OE seedlings was partially rescued by EGTA. An analysis of salt-responsive genes revealed that the transcription of RD29B, MYB15 and ZAT10 was inversely regulated in plc4 mutant and AtPLC4 OE seedlings. Our findings suggest that AtPLC4 negatively regulates the salt tolerance of Arabidopsis seedlings, and Ca(2+) may be involved in regulating this process.
DOI:10.1371/journal.pgen.1005511URLPMID:26401841 [本文引用: 1]
Phosphoinositides represent important lipid signals in the plant development and stress response. However, multiple isoforms of the phosphoinositide biosynthetic genes hamper our understanding of the pivotal enzymes in each step of the pathway as well as their roles in plant growth and development. Here, we report that phosphoinositide-specific phospholipase C2 (AtPLC2) is the primary phospholipase in phosphoinositide metabolism and is involved in seedling growth and the endoplasmic reticulum (ER) stress responses in Arabidopsis thaliana. Lipidomic profiling of multiple plc mutants showed that the plc2-1 mutant increased levels of its substrates phosphatidylinositol 4-phosphate and phosphatidylinositol 4,5-bisphosphate, suggesting that the major phosphoinositide metabolic pathway is impaired. AtPLC2 displayed a distinct tissue expression pattern and localized at the plasma membrane in different cell types, where phosphoinositide signaling occurs. The seedlings of plc2-1 mutant showed growth defect that was complemented by heterologous expression of AtPLC2, suggesting that phosphoinositide-specific phospholipase C activity borne by AtPLC2 is required for seedling growth. Moreover, the plc2-1 mutant showed hypersensitive response to ER stress as evidenced by changes in relevant phenotypes and gene expression profiles. Our results revealed the primary enzyme in phosphoinositide metabolism, its involvement in seedling growth and an emerging link between phosphoinositide and the ER stress response.
DOI:10.1111/tpj.2015.84.issue-3URL [本文引用: 2]
DOI:10.1093/pcp/pcy120URLPMID:30107538 [本文引用: 1]
Phospholipase C (PLC) is a well-known signaling enzyme in metazoans that hydrolyzes phosphatidylinositol 4,5-bisphosphate (PIP2) to produce inositol 1,4,5-trisphosphate and diacylglycerol as second messengers involved in mutiple processes. Plants contain PLC too, but relatively little is known about its function there. The model system Arabidopsis thaliana contains nine PLC genes. Reversed genetics have implicated several roles for PLCs in plant development and stress signaling. Here, PLC5 is functionally addressed. Promoter-beta-glucuronidase (GUS) analyses revealed expression in roots, leaves and flowers, predominantly in vascular tissue, most probably phloem companion cells, but also in guard cells, trichomes and root apical meristem. Only one plc5-1 knock-down mutant was obtained, which developed normally but grew more slowly and exhibited reduced primary root growth and decreased lateral root numbers. These phenotypes could be complemented by expressing the wild-type gene behind its own promoter. Overexpression of PLC5 (PLC5-OE) using the UBQ10 promoter resulted in reduced primary and secondary root growth, stunted root hairs, decreased stomatal aperture and improved drought tolerance. PLC5-OE lines exhibited strongly reduced phosphatidylinositol 4-monophosphate (PIP) and PIP2 levels and increased amounts of phosphatidic acid, indicating enhanced PLC activity in vivo. Reduced PIP2 levels and stunted root hair growth of PLC5-OE seedlings could be recovered by inducible overexpression of a root hair-specific PIP 5-kinase, PIP5K3. Our results show that PLC5 is involved in primary and secondary root growth and that its overexpression improves drought tolerance. Independently, we provide new evidence that PIP2 is essential for the polar tip growth of root hairs.
DOI:10.1093/pcp/pcx194URLPMID:29309666 [本文引用: 2]
Phospholipase C (PLC) is well known for its role in animal signaling, where it generates the second messengers, inositol 1,4,5-trisphosphate (IP3) and diacylglycerol (DAG), by hydrolyzing the minor phospholipid, phosphatidylinositol 4,5-bisphosphate (PIP2), upon receptor stimulation. In plants, PLC's role is still unclear, especially because the primary targets of both second messengers are lacking, i.e. the ligand-gated Ca2+ channel and protein kinase C, and because PIP2 levels are extremely low. Nonetheless, the Arabidopsis genome encodes nine PLCs. We used a reversed-genetic approach to explore PLC's function in Arabidopsis, and report here that PLC3 is required for proper root development, seed germination and stomatal opening. Two independent knock-down mutants, plc3-2 and plc3-3, were found to exhibit reduced lateral root densities by 10-20%. Mutant seeds germinated more slowly but were less sensitive to ABA to prevent germination. Guard cells of plc3 were also compromised in ABA-dependent stomatal closure. Promoter-beta-glucuronidase (GUS) analyses confirmed PLC3 expression in guard cells and germinating seeds, and revealed that the majority is expressed in vascular tissue, most probably phloem companion cells, in roots, leaves and flowers. In vivo 32Pi labeling revealed that ABA stimulated the formation of PIP2 in germinating seeds and guard cell-enriched leaf peels, which was significantly reduced in plc3 mutants. Overexpression of PLC3 had no effect on root system architecture or seed germination, but increased the plant's tolerance to drought. Our results provide genetic evidence for PLC's involvement in plant development and ABA signaling, and confirm earlier observations that overexpression increases drought tolerance. Potential molecular mechanisms for the above observations are discussed.
DOI:10.3389/fpls.2018.01721URLPMID:30542361 [本文引用: 2]
Phospholipase C (PLC) has been suggested to play important roles in plant stress and development. To increase our understanding of PLC signaling in plants, we have started to analyze knock-out (KO), knock-down (KD) and overexpression mutants of Arabidopsis thaliana, which contains nine PLCs. Earlier, we characterized PLC2, PLC3 and PLC5. Here, the role of PLC7 is functionally addressed. Promoter-GUS analyses revealed that PLC7 is specifically expressed in the phloem of roots, leaves and flowers, and is also present in trichomes and hydathodes. Two T-DNA insertion mutants were obtained, i.e., plc7-3 being a KO- and plc7-4 a KD line. In contrast to earlier characterized phloem-expressed PLC mutants, i.e., plc3 and plc5, no defects in primary- or lateral root development were found for plc7 mutants. Like plc3 mutants, they were less sensitive to ABA during stomatal closure. Double-knockout plc3 plc7 lines were lethal, but plc5 plc7 (plc5/7) double mutants were viable, and revealed several new phenotypes, not observed earlier in the single mutants. These include a defect in seed mucilage, enhanced leaf serration, and an increased tolerance to drought. Overexpression of PLC7 enhanced drought tolerance too, similar to what was earlier found for PLC3-and PLC5 overexpression. In vivo (32)Pi-labeling of seedlings and treatment with sorbitol to mimic drought stress, revealed stronger PIP2 responses in both drought-tolerant plc5/7 and PLC7-OE mutants. Together, these results show novel functions for PLC in plant stress and development. Potential molecular mechanisms are discussed.
DOI:10.1371/journal.pone.0062494URLPMID:23638098 [本文引用: 2]
BACKGROUND: Phospholipase C (PLC) is one of the major lipid hydrolysing enzymes, implicated in lipid mediated signaling. PLCs have been found to play a significant role in abiotic stress triggered signaling and developmental processes in various plant species. Genome wide identification and expression analysis have been carried out for this gene family in Arabidopsis, yet not much has been accomplished in crop plant rice. METHODOLOGY/PRINCIPAL FINDINGS: An exhaustive in-silico exploration of rice genome using various online databases and tools resulted in the identification of nine PLC encoding genes. Based on sequence, motif and phylogenetic analysis rice PLC gene family could be divided into phosphatidylinositol-specific PLCs (PI-PLCs) and phosphatidylcholine- PLCs (PC-PLC or NPC) classes with four and five members, respectively. A comparative analysis revealed that PLCs are conserved in Arabidopsis (dicots) and rice (monocot) at gene structure and protein level but they might have evolved through a separate evolutionary path. Transcript profiling using gene chip microarray and quantitative RT-PCR showed that most of the PLC members expressed significantly and differentially under abiotic stresses (salt, cold and drought) and during various developmental stages with condition/stage specific and overlapping expression. This finding suggested an important role of different rice PLC members in abiotic stress triggered signaling and plant development, which was also supported by the presence of relevant cis-regulatory elements in their promoters. Sub-cellular localization of few selected PLC members in Nicotiana benthamiana and onion epidermal cells has provided a clue about their site of action and functional behaviour. CONCLUSION/SIGNIFICANCE: The genome wide identification, structural and expression analysis and knowledge of sub-cellular localization of PLC gene family envisage the functional characterization of these genes in crop plants in near future.
DOI:10.1016/j.ceca.2015.04.003URLPMID:25933832 [本文引用: 1]
Phospholipase C (PLC), a major membrane phospholipid hydrolyzing enzyme generates signaling messengers such as diacylglycerol (DAG) and inositol 1,4,5-trisphosphate (IP3) in animals, and their phosphorylated forms such as phosphatidic acid (PA) and inositol hexakisphosphate (IP6) are thought to regulate various cellular processes in plants. Based on substrate specificity, plant PLC family is sub-divided into phosphatidylinositol-PLC (PI-PLC) and phosphatidylcholine-PLC (PC-PLC) groups. The activity of plant PLCs is regulated by various factors and the major ones include, Ca(2+) concentration, phospholipid substrate, post-translational modifications and interacting proteins. Most of the PLC members have been localized at the plasma membrane, suited for their function of membrane lipid hydrolysis. Several PLC members have been implicated in various cellular processes and signaling networks, triggered in response to a number of environmental cues and developmental events in different plant species, which makes them potential candidates for genetically engineering the crop plants for stress tolerance and enhancing the crop productivity. In this review article, we are focusing mainly on the plant PLC signaling and regulation, potential cellular and physiological role in different abiotic and biotic stresses, nutrient deficiency, growth and development.
DOI:10.1016/j.biochi.2013.07.004URLPMID:23856562 [本文引用: 1]
Phosphoinositide-specific phospholipase C (PI-PLC) cleaves, in a Ca(2+)-dependent manner, phosphatidylinositol-4,5-bisphosphate (PI-4,5-P2) into diacylglycerol (DAG) and inositol triphosphate (IP3). PI-PLCs are multidomain proteins that are structurally related to the PI-PLCzetas, the simplest animal PI-PLCs. Like these animal counterparts, they are only composed of EF-hand, X/Y and C2 domains. However, plant PI-PLCs do not have a conventional EF-hand domain since they are often truncated, while some PI-PLCs have no EF-hand domain at all. Despite this simple structure, plant PI-PLCs are involved in many essential plant processes, either associated with development or in response to environmental stresses. The action of PI-PLCs relies on the mediators they produce. In plants, IP3 does not seem to be the sole active soluble molecule. Inositol pentakisphosphate (IP5) and inositol hexakisphosphate (IP6) also transmit signals, thus highlighting the importance of coupling PI-PLC action with inositol-phosphate kinases and phosphatases. PI-PLCs also produce a lipid molecule, but plant PI-PLC pathways show a peculiarity in that the active lipid does not appear to be DAG but its phosphorylated form, phosphatidic acid (PA). Besides, PI-PLCs can also act by altering their substrate levels. Taken together, plant PI-PLCs show functional differences when compared to their animal counterparts. However, they act on similar general signalling pathways including calcium homeostasis and cell phosphoproteome. Several important questions remain unanswered. The cross-talk between the soluble and lipid mediators generated by plant PI-PLCs is not understood and how the coupling between PI-PLCs and inositol-kinases or DAG-kinases is carried out remains to be established.
DOI:10.1016/0014-5793(87)80515-xURLPMID:2822482 [本文引用: 2]
A polyphosphoinositide phospholipase C has been identified in highly purified plasma membranes from shoots and roots of wheat seedlings. The enzyme preferentially hydrolysed phosphatidylinositol 4-phosphate and phosphatidylinositol 4,5-bisphosphate and had a different phosphoinositide substrate profile from soluble phospholipase C. The enzyme activity was lower in plasma membranes isolated from light-grown shoots than from dark-grown ones, whereas no differences in activity between plasma membranes from light- and dark-grown roots were seen. Maximum activity of the membrane-bound enzyme was observed around pH 6. It was activated by micromolar concentrations of Ca2+, but not by GTP or GTP analogues. The enzyme may participate in signal transduction over the plant plasma membrane.
DOI:10.1016/0005-2760(92)90107-7URLPMID:1310875 [本文引用: 2]
The effect of various detergents on polyphosphoinositide-specific phospholipase C activity in highly purified wheat root plasma membrane vesicles was examined. The plasma membrane-bound enzyme was solubilized in octylglucoside and purified 25-fold by hydroxylapatite and ion-exchange chromatography. The purified enzyme catalyzed the hydrolysis of phosphatidylinositol 4-phosphate (PIP) and phosphatidylinositol 4,5-bisphosphate (PIP2) with specific activities of 5 and 10 mumol/min per mg protein, respectively. Phosphatidylinositol (PI) was not a substrate. Optimum activity was between pH 6-7 (PIP) and pH 6-6.5 (PIP2). The enzyme was dependent on micromolar concentrations of Ca2+ for activity, and millimolar Mg2+ further increased the activity. Other divalent cations (4 mM Ca2+, Mn2+ and Co2+) inhibited (PIP2 as substrate) or enhanced (PIP as substrate) phospholipase C activity.
DOI:10.1104/pp.100.3.1296URLPMID:16653120 [本文引用: 2]
Polyphosphoinositide-specific phospholipase C activity was present in plasma membranes isolated from different tissues of several higher plants. Phospholipase C activities against added phosphatidylinositol 4-phosphate (PIP) and phosphatidylinositol 4,5-bisphosphate (PIP(2)) were further characterized in plasma membrane fractions isolated from shoots and roots of dark-grown wheat (Triticum aestivum L. cv Drabant) seedlings. In right-side-out (70-80% apoplastic side out) plasma membrane vesicles, the activities were increased 3 to 5 times upon addition of 0.01 to 0.025% (w/v) sodium deoxycholate, whereas in fractions enriched in inside-out (70-80% cytoplasmic side out) vesicles, the activities were only slightly increased by detergent. Furthermore, the activities of inside-out vesicles in the absence of detergent were very close to those of right-side-out vesicles in the presence of optimal detergent concentration. This verifies the general assumption that polyphosphoinositide phospholipase C activity is located at the cytoplasmic surface of the plasma membrane. PIP and PIP(2) phospholipase C was dependent on Ca(2+) with maximum activity at 10 to 100 mum free Ca(2+) and half-maximal activation at 0.1 to 1 mum free Ca(2+). In the presence of 10 mum Ca(2+), 1 to 2 mm MgCl(2) or MgSO(4) further stimulated the enzyme activity. The other divalent chloride salts tested (1.5 mm Ba(2+), Co(2+), Cu(2+), Mn(2+), Ni(2+), and Zn(2+)) inhibited the enzyme activity. The stimulatory effect by Mg(2+) was observed also when 35 mm NaCl was included. Thus, the PIP and PIP(2) phospholipase C exhibited maximum in vitro activity at physiologically relevant ion concentrations. The plant plasma membrane also possessed a phospholipase C activity against phosphatidylinositol that was 40 times lower than that observed with PIP or PIP(2) as substrate. The phosphatidylinositol phospholipase C activity was dependent on Ca(2+), with maximum activity at 1 mm CaCl(2), and could not be further stimulated by Mg(2+).
DOI:10.1371/journal.pone.0105061URLPMID:25121594 [本文引用: 2]
Plant phosphoinositide-specific phospholipases C (PI-PLCs) function in several essential plant processes associated with either development or environmental stress. In this report, we examined the expression patterns of TaPLC1 under drought and high salinity stress at the transcriptional and post-transcriptional levels. TaPLC1 mRNA was expressed in all wheat organs examined. U73122 and edelfosine, the PLC inhibitor, impaired seedling growth and enhanced seedling sensitivity to drought and high salinity stress. Though TaPLC1 expression in wheat was lowest at the seedling stage, it was strongly induced under conditions of stress. When 6-day-old wheat seedlings were treated with 200 mM NaCl or 20% (w/v) PEG 6000 for 6 or 12 h, respectively, the TaPLC1 transcript level increased by 16-fold compared to the control. Western blotting showed that the TaPLC protein concentration was also maintained at a high level from 24 to 48 h during stress treatment. Together, our results indicate the possible biological functions of TaPLC1 in regulating seedling growth and the response to drought and salinity stress.
[本文引用: 2]
[本文引用: 2]
URL [本文引用: 2]
Using drought resistant and high-yielding crop varieties is critical for high and stable crop productivity under rainfed farming condition. In this study, an experiment was conducted at Nanpi Eco-Agricultural Experimental Station of Chinese Academy of Sciences (116°40′E, 38°00′N) in 2014–2015 to investigate the performance of ‘Xiaoyan 60’ wheat cultivar under the rainfed condition. The objective of the study was to test the adaptability of ‘Xiaoyan 60’ wheat cultivar to late sowing and yield loss compensation by increased seeding rate. Treatments included two factors, sowing date and seeding density. Six sowing dates were set, which were from the 15th of October to the 14th of November with six days interval, and denoted as T1, T2, T3, T4, T5 and T6, respectively. Then there were two treatments of seeding densities — constant seeding rate (B1) and increased seeding rates with delayed sowing date (B2). In B1, sowing density was 300 kg·hm-2 for all sowing dates, whereas in B2, it started at 300 kg·hm-2 and progressively increased at 7.5 kg·hm-2·day1 with delayed sowing date. Thus the sowing densities at sowing dates in B2 treatments were 300 kg·hm-2 for T1, 345 kg·hm-2 for T2, 390 kg·hm-2 for T3, 435 kg·hm-2 for T4, 480 kg·hm-2 for T5 and 525 kg·hm-2 for T6. The population properties, growth, yield and water use characteristics were investigated under different treatments. The results showed that the plant population traits, yield and water use efficiency varied with sowing date and seeding density. The growth period shortened with delayed sowing date, the duration for all the growth stages was also shortened. In contrast, seeding rate had no effect on growth period, but positively influenced the number of seedlings, number of spikes and dry biomass amount. Also plant height was not affected by seeding density. Delayed sowing gradually decreased the rate of seedling emergence, number of spikes per plant, plant height and dry biomass. Similarly, grain yield decreased with delayed sowing. However, the grain yield reached 6 600 kg·hm-2 level through increase seeding density to compensate for delayed sowing. There was no significant difference among the first four sowing dates (from T1 to T4) for grain-yield-based water use efficiency, which was above 29 kg·hm-2·mm-1. Because ‘Xiaoyan 60’ wheat cultivar was strongly adaptable to late sowing, it was recommended for cultivation under the rainfed farming conditions. Yield loss due to delayed sowing was compensated for by increasing seeding rate, which ensured optimum plant population. The correlation between seeding density (y) and delayed days (x) of sowing could be decribed by the regression equation y = 0.368 2x2 + 1.193 9x+316.7 (R2 = 0.98).
URL [本文引用: 2]
Using drought resistant and high-yielding crop varieties is critical for high and stable crop productivity under rainfed farming condition. In this study, an experiment was conducted at Nanpi Eco-Agricultural Experimental Station of Chinese Academy of Sciences (116°40′E, 38°00′N) in 2014–2015 to investigate the performance of ‘Xiaoyan 60’ wheat cultivar under the rainfed condition. The objective of the study was to test the adaptability of ‘Xiaoyan 60’ wheat cultivar to late sowing and yield loss compensation by increased seeding rate. Treatments included two factors, sowing date and seeding density. Six sowing dates were set, which were from the 15th of October to the 14th of November with six days interval, and denoted as T1, T2, T3, T4, T5 and T6, respectively. Then there were two treatments of seeding densities — constant seeding rate (B1) and increased seeding rates with delayed sowing date (B2). In B1, sowing density was 300 kg·hm-2 for all sowing dates, whereas in B2, it started at 300 kg·hm-2 and progressively increased at 7.5 kg·hm-2·day1 with delayed sowing date. Thus the sowing densities at sowing dates in B2 treatments were 300 kg·hm-2 for T1, 345 kg·hm-2 for T2, 390 kg·hm-2 for T3, 435 kg·hm-2 for T4, 480 kg·hm-2 for T5 and 525 kg·hm-2 for T6. The population properties, growth, yield and water use characteristics were investigated under different treatments. The results showed that the plant population traits, yield and water use efficiency varied with sowing date and seeding density. The growth period shortened with delayed sowing date, the duration for all the growth stages was also shortened. In contrast, seeding rate had no effect on growth period, but positively influenced the number of seedlings, number of spikes and dry biomass amount. Also plant height was not affected by seeding density. Delayed sowing gradually decreased the rate of seedling emergence, number of spikes per plant, plant height and dry biomass. Similarly, grain yield decreased with delayed sowing. However, the grain yield reached 6 600 kg·hm-2 level through increase seeding density to compensate for delayed sowing. There was no significant difference among the first four sowing dates (from T1 to T4) for grain-yield-based water use efficiency, which was above 29 kg·hm-2·mm-1. Because ‘Xiaoyan 60’ wheat cultivar was strongly adaptable to late sowing, it was recommended for cultivation under the rainfed farming conditions. Yield loss due to delayed sowing was compensated for by increasing seeding rate, which ensured optimum plant population. The correlation between seeding density (y) and delayed days (x) of sowing could be decribed by the regression equation y = 0.368 2x2 + 1.193 9x+316.7 (R2 = 0.98).
DOI:10.1016/j.plaphy.2008.04.015URL [本文引用: 3]
Abstract
Phosphoinositide-specific phospholipase C cleaves the substrate phosphatidylinositol 4,5-bisphosphate and generates inositol 1,4,5-trisphosphate and 1,2-diacylglycerol, both of which are second messengers in the phosphoinositide signal transduction pathways operative in animal cells. Five PI-PLC isoforms, β, γ, δ, ? and ζ, have been identified in mammals. Plant PI-PLCs are structurally close to the mammalian PI-PLC-ζ isoform. The Arabidopsis genome contains nine AtPLC genes. Expression patterns of all nine genes in different organs and in response to various environmental stimuli were studied by applying a quantitative RT–PCR approach. Multiple members of the gene family were differentially expressed in Arabidopsis organs, suggesting putative roles for this enzyme in plant development, including tissue and organ differentiation. This study also shows that a majority of the AtPLC genes are induced in response to various environmental stimuli, including cold, salt, nutrients Murashige–Skoog salts, dehydration, and the plant hormone abscisic acid. Results of this and previous studies strongly suggest that transcriptional activation of the PI-PLC gene family is important for adapting plants to stress environments. Expression patterns and phylogenetic relationships indicates that AtPLC gene members probably evolved through multiple rounds of gene duplication events, with AtPLC4 and AtPLC5 and AtPLC8 and AtPLC9 being duplicated in tandem in recent times.DOI:10.1371/journal.pone.0062494URLPMID:23638098 [本文引用: 1]
BACKGROUND: Phospholipase C (PLC) is one of the major lipid hydrolysing enzymes, implicated in lipid mediated signaling. PLCs have been found to play a significant role in abiotic stress triggered signaling and developmental processes in various plant species. Genome wide identification and expression analysis have been carried out for this gene family in Arabidopsis, yet not much has been accomplished in crop plant rice. METHODOLOGY/PRINCIPAL FINDINGS: An exhaustive in-silico exploration of rice genome using various online databases and tools resulted in the identification of nine PLC encoding genes. Based on sequence, motif and phylogenetic analysis rice PLC gene family could be divided into phosphatidylinositol-specific PLCs (PI-PLCs) and phosphatidylcholine- PLCs (PC-PLC or NPC) classes with four and five members, respectively. A comparative analysis revealed that PLCs are conserved in Arabidopsis (dicots) and rice (monocot) at gene structure and protein level but they might have evolved through a separate evolutionary path. Transcript profiling using gene chip microarray and quantitative RT-PCR showed that most of the PLC members expressed significantly and differentially under abiotic stresses (salt, cold and drought) and during various developmental stages with condition/stage specific and overlapping expression. This finding suggested an important role of different rice PLC members in abiotic stress triggered signaling and plant development, which was also supported by the presence of relevant cis-regulatory elements in their promoters. Sub-cellular localization of few selected PLC members in Nicotiana benthamiana and onion epidermal cells has provided a clue about their site of action and functional behaviour. CONCLUSION/SIGNIFICANCE: The genome wide identification, structural and expression analysis and knowledge of sub-cellular localization of PLC gene family envisage the functional characterization of these genes in crop plants in near future.
DOI:10.1111/j.1365-313X.2010.04136.xURL [本文引用: 2]
DOI:10.1007/s11103-011-9801-1URL [本文引用: 1]
DOI:10.1111/j.1467-7652.2010.00547.xURL [本文引用: 1]
P>Transcription factors have been shown to control the activity of multiple stress response genes in a coordinated manner and therefore represent attractive targets for application in molecular plant breeding. We investigated the possibility of modulating the transcriptional regulation of drought and cold responses in the agriculturally important species, wheat and barley, with a view to increase drought and frost tolerance. Transgenic wheat and barley plants were generated showing constitutive (double 35S) and drought-inducible (maize Rab17) expression of the TaDREB2 and TaDREB3 transcription factors isolated from wheat grain. Transgenic populations with constitutive over-expression showed slower growth, delayed flowering and lower grain yields relative to the nontransgenic controls. However, both the TaDREB2 and TaDREB3 transgenic plants showed improved survival under severe drought conditions relative to nontransgenic controls. There were two components to the drought tolerance: real (activation of drought-stress-inducible genes) and 'seeming' (consumption of less water as a result of smaller size and/or slower growth of transgenics compared to controls). The undesired changes in plant development associated with the 'seeming' component of tolerance could be alleviated by using a drought-inducible promoter. In addition to drought tolerance, both TaDREB2 and TaDREB3 transgenic plants with constitutive over-expression of the transgene showed a significant improvement in frost tolerance. The increased expression of TaDREB2 and TaDREB3 lead to elevated expression in the transgenics of 10 other CBF/DREB genes and a large number of stress responsive LEA/COR/DHN genes known to be responsible for the protection of cell from damage and desiccation under stress.
DOI:10.1073/pnas.92.9.3903URLPMID:7732004 [本文引用: 2]
A cDNA corresponding to a putative phosphatidylinositol-specific phospholipase C (PI-PLC) in the higher plant Arabidopsis thaliana was cloned by use of the polymerase chain reaction. The cDNA, designated cAtPLC1, encodes a putative polypeptide of 561 aa with a calculated molecular mass of 64 kDa. The putative product includes so-called X and Y domains found in all PI-PLCs identified to date. In mammalian cells, there are three types of PI-PLC, PLC-beta, -gamma, and -delta. The overall structure of the putative AtPLC1 protein is most similar to that of PLC-delta, although the AtPLC1 protein is much smaller than PLCs from other organisms. The recombinant AtPLC1 protein synthesized in Escherichia coli was able to hydrolyze phosphatidylinositol 4,5-bisphosphate and this activity was completely dependent on Ca2+, as observed also for mammalian PI-PLCs. These results suggest that the AtPLC1 gene encodes a genuine PI-PLC of a higher plant. Northern blot analysis showed that the AtPLC1 gene is expressed at very low levels in the plant under normal conditions but is induced to a significant extent under various environmental stresses, such as dehydration, salinity, and low temperature. These observations suggest that AtPLC1 might be involved in the signal-transduction pathways of environmental stresses and that an increase in the level of AtPLC1 might amplify the signal, in a manner that contributes to the adaptation of the plant to these stresses.
DOI:10.1038/sj.cr.7290200URLPMID:15040888 [本文引用: 1]
The phosphatidylinositol (PI) metabolic pathway is considered critical in plant responses to many environmental factors, and previous studies have indicated the involvement of multiple PI-related gene families during cellular responses. Through a detailed analysis of the Arabidopsis thaliana genome, 82 polypeptides were identified as being involved in PI signaling. These could be grouped into different families including PI synthases (PIS), PI-phosphate kinases (PIPK), phospholipases (PL), inositol polyphosphate phosphatases (IPPase), inositol polyphosphate kinases (IPK), PI transfer proteins and putative inositol polyphosphate receptors. The presence of more than 10 isoforms of PIPK, PLC, PLD and IPPase suggested that these genes might be differentially expressed during plant cellular responses or growth and development. Accordingly, DNA chip technology was employed to study the expression patterns of various isoforms. In total, 79 mRNA clones were amplified and used for DNA chip generation. Expression profile analysis was performed using samples that represented multiple tissues or cellular responses. Tested samples included normal leaf, stem and flower tissues, and leaves from plants treated with various hormones (auxin, cytokinin, gibberellin, abscisic acid and brassinosteroid) or environmental factors (temperature, calcium, sodium, drought, salicylic acid and jasmonic acid). Results showed that many PI pathway-related genes were differentially expressed under these experimental conditions. In particular, the different isoforms of each family were specifically expressed in many cases, suggesting their involvement in tissue specificity and cellular responses to environmental conditions. This work provides a starting point for functional studies of the relevant PI-related proteins and may help shed light onto the role of PI pathways in development and cellular responses.
DOI:10.1016/j.plaphy.2008.04.015URL [本文引用: 1]
Abstract
Phosphoinositide-specific phospholipase C cleaves the substrate phosphatidylinositol 4,5-bisphosphate and generates inositol 1,4,5-trisphosphate and 1,2-diacylglycerol, both of which are second messengers in the phosphoinositide signal transduction pathways operative in animal cells. Five PI-PLC isoforms, β, γ, δ, ? and ζ, have been identified in mammals. Plant PI-PLCs are structurally close to the mammalian PI-PLC-ζ isoform. The Arabidopsis genome contains nine AtPLC genes. Expression patterns of all nine genes in different organs and in response to various environmental stimuli were studied by applying a quantitative RT–PCR approach. Multiple members of the gene family were differentially expressed in Arabidopsis organs, suggesting putative roles for this enzyme in plant development, including tissue and organ differentiation. This study also shows that a majority of the AtPLC genes are induced in response to various environmental stimuli, including cold, salt, nutrients Murashige–Skoog salts, dehydration, and the plant hormone abscisic acid. Results of this and previous studies strongly suggest that transcriptional activation of the PI-PLC gene family is important for adapting plants to stress environments. Expression patterns and phylogenetic relationships indicates that AtPLC gene members probably evolved through multiple rounds of gene duplication events, with AtPLC4 and AtPLC5 and AtPLC8 and AtPLC9 being duplicated in tandem in recent times.DOI:10.1007/s10529-005-5802-yURL
A cDNA encoding a phosphoinositide-specific phospholipase C (PI-PLC) has been isolated from Zea mays by screening a cDNA library. The cDNA, designated ZmPLC, encodes a polypeptide of 586 amino acids, containing the catalytic X, Y and C2 domains found in all PI-PLCs from plants. Northern blot analysis showed that the expression of the ZmPLC gene in roots is up-regulated under conditions of high salt, dehydration, cold or low osmotic stress conditions. Recombinant ZmPLC protein was expressed in Esch-erichia coli, purified and used to produce polyclonal antibody, this polyclonal antibody is important for further studies to assess the ultimate function of the ZmPLC gene in plants.
DOI:10.1093/pcp/pcp051URLPMID:19369274 [本文引用: 1]
Salinity is one of the major environmental factors limiting growth and productivity of rice plants. In this study, the effect of salt stress on phospholipid signaling responses in rice leaves was investigated. Leaf cuts were radiolabeled with 32P-orthophosphate and the lipids extracted and analyzed by thin-layer chromatography, autoradiography and phosphoimaging. Phospholipids were identified by co-migration of known standards. Results showed that 32P(i) was rapidly incorporated into the minor lipids, phosphatidylinositol bisphosphate (PIP2) and phosphatidic acid (PA) and, interestingly, also into the structural lipids phosphatidylethanolamine (PE) and phosphatidylglycerol (PG), which normally label relatively slowly, like phosphatidylcholine (PC) and phosphatidylinositol (PI). Only very small amounts of PIP2 were found. However, in response to salt stress (NaCl), PIP2 levels rapidly (<30 min) increased up to 4-fold, in a time- and dose-dependent manner. PA and its phosphorylated product, diacylglycerolpyrophosphate (DGPP), also increased upon NaCl stress, while cardiolipin (CL) levels decreased. All other phospholipid levels remained unchanged. PA signaling can be generated via the combined action of phospholipase C (PLC) and diacylglycerol kinase (DGK) or directly via phospholipase D (PLD). The latter can be measured in vivo, using a transphosphatidylation assay. Interestingly, these measurements revealed that salt stress inhibited PLD activity, indicating that the salt stress-induced PA response was not due to PLD activity. Comparison of the 32P-lipid responses in salt-tolerant and salt-sensitive cultivars revealed no significant differences. Together these results show that salt stress rapidly activates several lipid responses in rice leaves but that these responses do not explain the difference in salt tolerance between sensitive and tolerant cultivars.
[D].
[本文引用: 1]
[D].
[本文引用: 1]
DOI:10.1111/j.1365-3040.2009.02027.xURLPMID:19671099 [本文引用: 1]
Phosphatidylinositol-specific phospholipase C (PtdIns-PLC2) plays a central role in the phosphatidylinositol-specific signal transduction pathway. It catalyses the hydrolysis of membrane-bound phosphatidylinositol 4,5-bisphosphate to produce two second messengers, sn-1,2-diacylglycerol and inositol 1,4,5-trisphosphate. The former is a membrane activator of protein kinase C in mammalian systems, and the latter is a Ca(2+) modulator which induces distinctive oscillating bursts of cytosolic Ca(2+), resulting in regulation of gene expression and activation of proteins. Sustained over-expression of BnPtdIns-PLC2 in transgenic Brassica napus lines brought about an early shift from vegetative to reproductive phases, and shorter maturation periods, accompanied by notable alterations in hormonal distribution patterns in various tissues. The photosynthetic rate increased, while stomata were partly closed. Numerous gene expression changes that included induction of stress-related genes such as glutathione S-transferase, hormone-regulated and regulatory genes, in addition to a number of kinases, calcium-regulated factors and transcription factors, were observed. Other changes included increased phytic acid levels and phytohormone organization patterns. These results suggest the importance of PtdIns-PLC2 as an elicitor of a battery of events that systematically control hormone regulation, and plant growth and development in what may be a preprogrammed mode.
DOI:10.11983/CBB17143URL [本文引用: 1]
植物的生长发育容易受到外界环境变化的影响。非生物胁迫发生时, 表观遗传机制对胁迫应答基因的表达调控发挥了十分重要的作用。近年来, 调控植物非生物胁迫应答的表观遗传机制研究取得了一系列重要进展, 为进一步深入解析植物响应非生物胁迫的分子机制奠定了基础。该文对DNA甲基化修饰、组蛋白修饰、染色质重塑和非编码RNA等主要表观遗传调控方式在植物响应非生物胁迫中的作用进行了简要综述。
DOI:10.11983/CBB17143URL [本文引用: 1]
植物的生长发育容易受到外界环境变化的影响。非生物胁迫发生时, 表观遗传机制对胁迫应答基因的表达调控发挥了十分重要的作用。近年来, 调控植物非生物胁迫应答的表观遗传机制研究取得了一系列重要进展, 为进一步深入解析植物响应非生物胁迫的分子机制奠定了基础。该文对DNA甲基化修饰、组蛋白修饰、染色质重塑和非编码RNA等主要表观遗传调控方式在植物响应非生物胁迫中的作用进行了简要综述。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}