 ,1, 季延海1,2, 孙旺旺3, 武占会1,2, 储昭胜1, 刘明池,1,2
,1, 季延海1,2, 孙旺旺3, 武占会1,2, 储昭胜1, 刘明池,1,2Response of Chloroplast Ultrastructure and Photosynthetic Physiology of Two Tomato Varieties to Low Light Stress
MENG XianMin,1, JI YanHai1,2, SUN WangWang3, WU ZhanHui1,2, CHU ZhaoSheng1, LIU MingChi,1,2通讯作者:
责任编辑: 赵伶俐
收稿日期:2020-05-21接受日期:2020-08-18网络出版日期:2021-03-01
| 基金资助: |
Received:2020-05-21Accepted:2020-08-18Online:2021-03-01
作者简介 About authors
孟宪敏,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (4917KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
孟宪敏, 季延海, 孙旺旺, 武占会, 储昭胜, 刘明池. 两个番茄品种叶绿体超微结构及光合生理对弱光胁迫的响应[J]. 中国农业科学, 2021, 54(5): 1017-1028 doi:10.3864/j.issn.0578-1752.2021.05.013
MENG XianMin, JI YanHai, SUN WangWang, WU ZhanHui, CHU ZhaoSheng, LIU MingChi.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】番茄(Solanum lycopersicum L.)喜温喜光,是全世界设施栽培中的主要作物之一,冬春季广泛栽培于中国北方的单坡日光温室[1]。但日光温室内由于墙体、骨架或保温被等遮挡易形成弱光及低温环境,成为冬春茬产量和品质降低的阻碍[2,3]。研究认为秋冬季番茄开花结果期,对自然环境光强要求约为2 160 μmol·m-2·s-1,日光温室内植株冠层上方平均日辐射量860 μmol·m-2·s-1,光照时长也低于室外,远不能满足番茄正常生长需求[4]。然而荷兰全年光照严重不足,尤其冬至前后最短光照时长仅为5—6 h,一天光照累积量最低为150—200 J·cm-2[5],但荷兰番茄、黄瓜产量分别可达800和1 000 t·hm-2,是中国平均水平的3—4倍[6]。经过笔者对国内外番茄品种的多年研究和生产实践表明,除先进的补光技术外,品种的耐弱光差异是荷兰番茄高产的重要因素之一。因此,深入研究我国与荷兰番茄品种在弱光下的响应差异,对指导中国日光温室番茄品种选择具有重要意义。【前人研究进展】研究表明,弱光环境导致植株株高增加,可能出现徒长趋势,影响叶绿体结构发育[7,8],叶片净光合速率Pn[9,10,11,12]、叶绿素含量和PSⅡ实际量子效率(ΦPSⅡ)下降[3],光合产物合成和输出减少[13,14],而膜脂过氧化程度较高,最终导致产量和品质降低,但因品种耐受性不同导致降低幅度有显著差异[3]。研究表明,叶片气体交换参数不仅与气孔密度及大小有关,也与气孔的空间分布格局有关[15]。ZHENG等[16]发现气孔在叶片上空间分布规则时,最小邻域距离L(d)值较小,这一分析结果是通过Ripley’s K函数将叶片上可利用所有单点距离的二阶矩阵,来探究这些点在不同尺度上二维分布格局[17]。气孔在叶片上空间分布的规律在小尺度范围内呈规则分布,而大尺度范围内随机分布。【本研究切入点】前人对弱光的研究多采用白炽灯、日光灯或农艺钠灯等,此类光源能效大、成效低,也并不完全符合植物生长的最佳光谱成分,而日渐兴起的发光二极管(LED)作为日光温室的重要补光光源之一,正在发挥更大优势[18,19]。目前,针对品种耐弱光差异机理的研究较少,而且对不同基因型品种间光合生理特征及气孔、叶绿体结构的系统性研究较薄弱,也少见关于弱光下番茄叶片气孔分布的报道。【拟解决的关键问题】本试验采用LED灯作为光源进行弱光模拟试验,通过对番茄幼苗生长、叶片光合特性、气孔特征及分布、叶绿体超微结构等的研究,探明中国和荷兰品种间耐弱光性差异的机理。1 材料与方法
试验于2018年在北京市农林科学院蔬菜研究中心人工气候室(LPH-1-CT)进行。1.1 试验材料
以北京市农林科学院蔬菜研究中心选育的不耐弱光樱桃番茄品种‘京丹1号’和荷兰瑞克斯旺公司选育的较耐弱光温室品种‘佳西娜’为试材。1.2 试验设计
采用72孔穴盘于2018年2月播种,所用基质为草炭﹕蛭石=2﹕1,于3叶一心时,挑选长势一致的健壮番茄植株随机分组,移至人工气候室开始试验。2个品种均设对照与弱光处理。以LED灯(PHILIPS)为光源,光质为红蓝光2﹕1[20],以正常光强300—350 μmol·m-2·s-1为对照,50%正常光照下光强70—80 μmol·m-2·s-1为弱光。共4个处理,各设3次重复,每个重复50株。光周期12 h,昼夜温度25℃/15℃,空气相对湿度60%—70%。苗期统一水肥管理,营养液采用北京市农林科学院蔬菜研究中心地下水改良番茄专用配方:5 mmol?L-1 KNO3、5 mmol?L-1 Ca(NO3)2·4H2O、2 mmol?L-1 MgSO4、1 mmol?L-1 KH2PO4、50 μmol?L-1 EDTA- NaFe、50 μmol?L-1 H3BO3、10 μmol?L-1 MnSO4·4H2O、0.8 μmol?L-1 ZnSO4·7H2O、0.4 μmol?L-1 CuSO4·5H2O、0.02 μmol?L-1 (NH4)6Mo7O24·4H2O)[21,22],85%磷酸调节营养液EC值(2.0±0.2)mS·cm-1,pH 6.5。为保证植株生长条件和胁迫程度的一致性,同一光强处理的架子上分别放置2个品种的番茄幼苗。处理12 h观测荧光成像,处理15 d测定生长、光合生理指标,观察叶片气孔、叶绿体超微结构。各指标均重复测定3次,取平均值。1.3 测定项目及方法
1.3.1 生长指标 每处理选取代表性植株12株,用钢尺测定株高、最大节间距;游标卡尺测定茎粗;直尺测定叶长、叶宽,并计算叶面积;烘干称重法测定植株干重,计算壮苗指数。1.3.2 酶活性 参考李合生[23]的方法测定超氧化物歧化酶(SOD)活性;参照CAKMAK等[24]的方法测定丙二醛(MDA)含量。
1.3.3 叶片气体交换参数和光合色素含量 于8:30 —11:00采用LI-6400XT光合仪(美国LI-COR)测定第3功能叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)及蒸腾速率(Tr)。采用6400-02B红蓝光源,对照光强800 μmol·m-2·s-1,弱光处理光强为100 μmol·m-2·s-1,CO2浓度(400±2)μmol·mol-1。将已测定光合参数的叶片摘下,参考ARNON[25]的95%乙醇浸提法测定并计算总叶绿素chl.(a+b)、叶绿素a、叶绿素b含量及叶绿素a/叶绿素b(chl.a/b)。
1.3.4 叶绿体超微结构 于上午8:30取第3功能叶片中部靠叶脉处附近2 mm×2 mm小块,用2.5%戊二醛固定,pH 7.2磷酸缓冲液冲洗,1%锇酸固定,乙醇-丙醇梯度脱水,Epon-812环氧树脂包埋,LKB-5型超薄切片机切片,醋酸铀-柠檬酸铅双重染色,透射电镜选取15—20个视野并拍照。
1.3.5 气孔特征及空间分布格局分析 取样方式同1.3.4,经冲洗及梯度脱水,CO2临界点干燥。把叶片粘到扫描样品台上,离子溅射仪喷涂镀膜,扫描电镜观测15—20个视野并拍照。Auto CAD 2009测量单个气孔的长度和宽度,计算周长(P)、面积(A)及形状指数(S)。S是通过计算单一气孔形状与相同面积圆之间的偏离程度来测量其形状的复杂程度[26]。
$S=P / 2 \sqrt{\pi A}$ (以圆为参照几何形状)
假定每个气孔都是叶片表面上分布的单点,气孔最中间位置为该单点的位置。取叶片远轴面中部,采用透明指甲油印迹法制作临时装片,于LEICA CTR 4000下观察,随机选5个视野,各拍3张照片。各处理选5张照片,计算气孔密度。利用Arc GIS 10.2将照片于相同的地理坐标、投影坐标系下进行数字化处理,再运用多距离空间聚类分析工具(Ripley’s K函数)对表征气孔分布状况的点进行空间分析[16,17],为了确定95%的可信任区间,采用蒙特卡洛算法模拟随机分布点1 000次,分析结果用L(d)值表示。
$ \mathrm{L}(\mathrm{d})=\sqrt{\mathrm{K}(\mathrm{d}) / \pi}-\mathrm{d}$
式中,L(d)表示最小邻域距离,K(d)为任何个体在一定空间尺度内对其他个体的期望值,d为空间尺度。
1.3.6 荧光参数及成像 处理12 h,采用Fluor Cam开放式叶绿素荧光成像系统(北京Eco Tech)测定第3功能叶片叶绿素荧光参数,以颜色的形式对参数大小进行注释,相应数据可直接从Report窗口导出。待测叶片经暗适应20 min,设定快门Shutter=1,敏感度Sensitivity=2.8,光照Act2=19,Super=20,采用protocols程序测定叶绿素荧光参数。
1.4 数据分析
用Microsoft Excel 2010及SPSS17.0软件进行数据处理与分析。2 结果
2.1 番茄叶片叶绿体结构对弱光的响应
图1显示,正常光照下,2个番茄品种叶片的叶绿体结构良好,呈长椭圆形或梭形,基粒片层垛叠紧致,基质片层结构清晰,叶绿体被膜清晰、完整,有少量嗜锇颗粒。弱光下,‘佳西娜’的叶绿体有轻微膨胀,基粒片层垛叠整齐有序,结构清晰,只有少数叶绿体内含淀粉粒;‘京丹1号’的叶绿体趋于圆形或短粗形,发生严重肿胀,叶绿体膜严重溶解,片层结构数量减少并且变得混乱,基粒片层部分解体,模糊不清,叶绿体与细胞壁脱离而游离分布在细胞内,产生淀粉粒较多并有嗜锇颗粒积累。图1
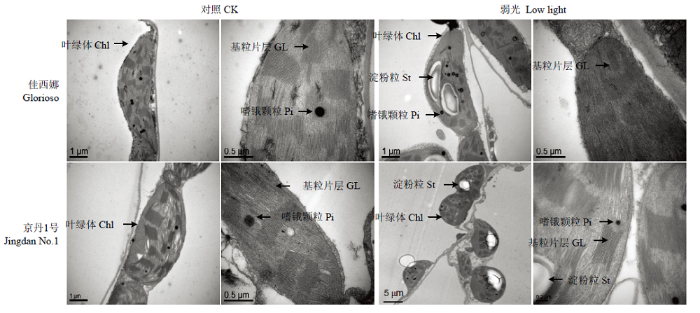 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1弱光对叶绿体超微结构的影响
Fig. 1Effects of low light on ultra-structures of leaf chloroplast
2.2 番茄叶片气孔特征及空间分布格局对弱光的响应
弱光显著降低了气孔密度及宽度(表1)。‘佳西娜’和‘京丹1号’的气孔密度分别比其对照降低47.39%、51.24%(P<0.05),气孔宽度分别比其对照降低19.77%、40.33%;弱光下,‘京丹1号’的气孔面积、周长、长度及形状指数最低(P>0.05)(图2、表1)。Table 1
表1
表1弱光对叶片气孔特征的影响
Table 1
| 处理 Treatment | 气孔特征 Stomatal characteristics | ||||||
|---|---|---|---|---|---|---|---|
| 密度Density (Number/mm2) | 面积 A (μm2) | 周长 P (μm) | 长度 Length (μm) | 宽度 Width (μm) | 形状指数 S | ||
| 佳西娜 Glorioso | 对照 CK | 306.22±92.15a | 20.04±2.05a | 31.41±6.54a | 14.72±2.98a | 1.77±0.59b | 1.21±0.68a |
| 弱光Low light | 161.10±57.27b | 17.14±1.41a | 28.54±2.99a | 12.93±1.33a | 1.42±0.35bc | 1.04±0.20a | |
| 京丹1号 Jingdan No.1 | 对照 CK | 303.15±97.72a | 25.93±5.27a | 30.51±1.35a | 13.72±1.16a | 2.43±0.61a | 1.37±0.41a |
| 弱光Low light | 147.81±52.42b | 14.03±2.18a | 24.89±2.39a | 12.53±3.30a | 1.45±0.47bc | 0.84±0.63a | |
新窗口打开|下载CSV
图2
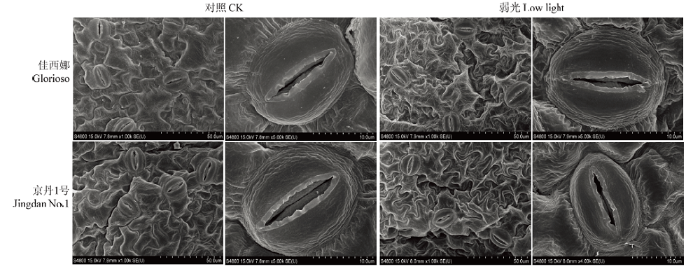 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2叶片单个气孔的扫描电子显微镜照片
Fig. 2Scanning electron photographs of individual stoma of leaves
对叶片上单个气孔的空间分布格局分析,结果如图3。气孔在小尺度范围内呈规则分布,在大尺度范围内随机分布。2个品种的叶片气孔空间分布格局与单个气孔间L(d)值对弱光响应有差异,即对照叶片的气孔由规则分布转变为随机的空间尺度约为300 μm,而弱光下空间尺度减小为250 μm。弱光下‘京丹1号’叶片单个气孔间L(d)比其对照提高5.95%,‘佳西娜’的L(d)值最小,比其对照降低34.78%,说明弱光下‘佳西娜’的气孔在叶片上呈现更规则的空间分布格局。
图3
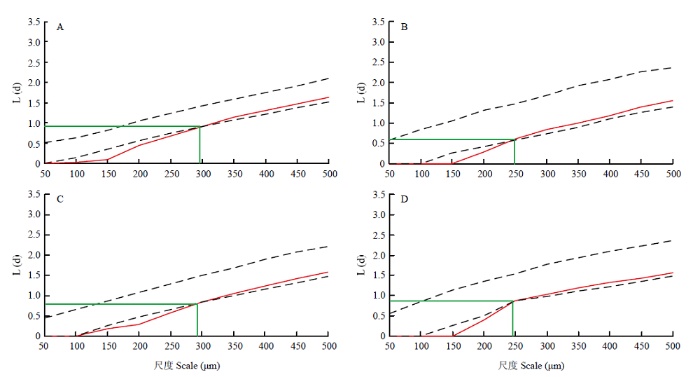 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3弱光对气孔空间分布格局的影响
A、B分别为‘佳西娜’的对照和弱光处理,C、D分别为‘京丹1号’的对照和弱光处理。图中上下两条虚线代
Fig. 3Effect of low light on spatial distribution pattern of stomata
A, B are CK and low light treatment of Glorioso; C, D are CK and low light treatment of Jingdan No.1, respectively. The upper and lower dashed lines represent the upper and lower boundaries of 95% confidence intervals, the red line represents the K(d) value, and the green line represents the corresponding spatial scale and L(d) value when stomatal distribution on leaves changed from regular to random. L(d) is the minimum neighborhood distance, when L(d) value is less than 95% confidence interval, the stomata distribution is regular in this scale, and the smaller of L(d), the more regular the spatial distribution of stomata
2.3 番茄叶片SOD活性及MDA含量对弱光的响应
由图4看出,弱光下2个番茄品种叶片SOD活性均显著低于同期对照处理,2个品种正常光照处理下(CK)差异不显著,弱光处理下‘佳西娜’SOD含量显著高于‘京丹1号’。弱光下2个番茄品种MDA含量显著上升,均显著高于正常光照处理。弱光下,‘佳西娜’和‘京丹1号’MDA分别较正常光照下显著提高28.88%和60.69%。图4
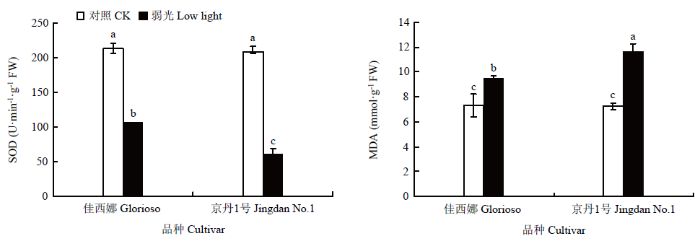 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4弱光对叶片SOD活性及MDA含量的影响
不同小写字母表示处理间差异显著(P<0.05)。下同
Fig. 4Effects of low light on SOD activity and MDA content in leaves
Different lower-case letters indicate significant differences among treatments (P<0.05). The same as below
2.4 番茄叶片叶绿素含量及气体交换参数对弱光的响应
弱光处理后,‘佳西娜’番茄叶片Chl.a、Chl.(a+b)含量和Chl.a/b均显著低于对照植株,分别降低37.81%、27.38%和43.47%;Chl.b含量高于对照植株,但与对照差异并未达显著性水平。‘京丹1号’的Chl.a、Chl.(a+b)含量均高于对照植株,但均不显著。弱光处理下,‘佳西娜’的Chl.b含量比‘京丹1号’显著增加37.50%,Chl.a/b比‘京丹1号’显著降低25.47%(表2)。Table 2
表2
表2弱光对叶片光合色素含量的影响
Table 2
| 处理 Treatment | 叶绿素a Chl.a (mg·g-1 FW) | 叶绿素b Chl.b (mg·g-1 FW) | Chl.(a+b) (mg·g-1 FW) | Chl.a/b | |
|---|---|---|---|---|---|
| 佳西娜 Glorioso | 对照 CK | 2.01±0.12a | 0.62±0.04ab | 2.63±0.15a | 3.25±0.02a |
| 弱光Low light | 1.25±0.02b | 0.66±0.09a | 1.91±0.07b | 1.99±0.33b | |
| 京丹1号 Jingdan No.1 | 对照 CK | 1.88±0.07ab | 0.63±0.03ab | 2.50±0.08a | 2.30±0.15a |
| 弱光Low light | 1.28±0.01b | 0.48±0.02b | 1.76±0.03b | 2.67±0.07a | |
新窗口打开|下载CSV
由表3可知,弱光下,2个番茄品种的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)均低于对照。弱光处理下,‘佳西娜’的Gs、Tr与对照差异不显著,而‘京丹1号’分别比其对照显著降低40.00%和27.64%。弱光下,‘佳西娜’和‘京丹1号’的Pn分别比其对照显著降低33.65%、60.91%。弱光下,‘佳西娜’的胞间CO2浓度(Ci)较其对照提高7.94%,但未达到显著性差异,而‘京丹1号’较其对照显著降低8.52%。
Table 3
表3
表3弱光对叶片光合参数的影响
Table 3
| 处理 Treatment | 净光合速率 Pn (μmol·m-2·s-1) | 气孔导度 Gs (mol·m-2·s-1) | 胞间CO2浓度 Ci (μmol·mol-1) | 蒸腾速率 Tr (mmol·m-2·s-1) | |
|---|---|---|---|---|---|
| 佳西娜 Glorioso | 对照 CK | 18.16±1.30a | 0.41±0.24a | 333.00±5.92a | 8.19±0.43a |
| 弱光Low light | 12.05±1.38b | 0.35±0.08ab | 359.44±34.45a | 7.14±1.52ab | |
| 京丹1号 Jingdan No.1 | 对照 CK | 17.09±2.17a | 0.40±0.18a | 346.35±36.94a | 7.49±1.84a |
| 弱光Low light | 6.68±2.57c | 0.24±0.08b | 316.84±13.03b | 5.42±1.36b | |
新窗口打开|下载CSV
2.5 番茄叶片叶绿素荧光参数与荧光成像对弱光的响应
光照对暗适应过程PSⅡ最大光量子效率QY_max(即Fv/Fm)影响不显著(图5-A,B)。弱光下,相比对照植株,从光适应下实际光量子效率QY_Ln到光稳态下实际光量子效率QY_Lss,‘佳西娜’的QY_Ln值降低幅度逐渐减小,而‘京丹1号’的QY_Ln一直在其对照曲线内侧,降低幅度显著高于‘佳西娜’。弱光下,‘佳西娜’的QY_Lss和暗弛豫过程中PSⅡ实际光量子效率QY_Dn基本没有变化,而‘京丹1号’的QY_Lss比其对照降低14.29%。图5
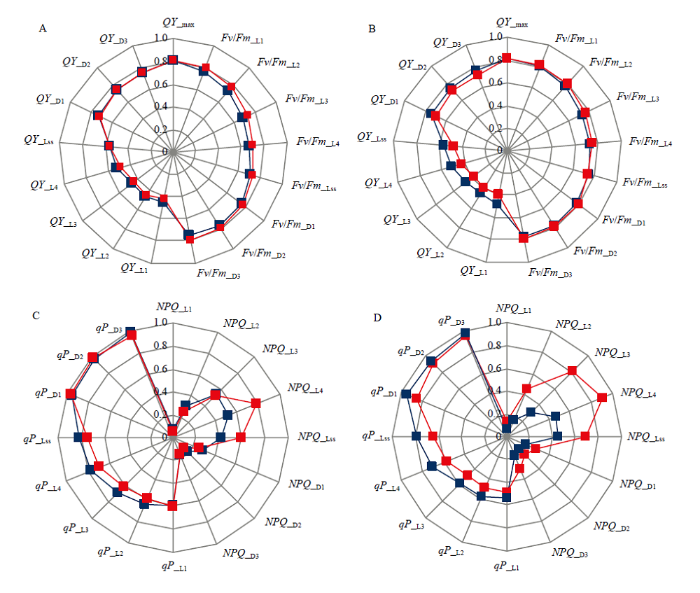 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5弱光对叶片叶绿素荧光参数的影响
蓝、红色线分别表示对照和弱光处理。A、C为‘佳西娜’,B、D为‘京丹1号’;A、B为光量子产量参数,C、D为荧光淬灭参数
Fig. 5Effects of low light on chlorophyll fluorescence parameters of leaves
The blue and red lines represent CK and low light treatment respectively. A, C are Glorioso; B, D are Jingdan No.1. A, B are photoquantum yield parameters; C, D are fluorescence quenching parameters
弱光下,2个番茄品种的NPQ升高,‘佳西娜’升高幅度小于‘京丹1号’(图5-C,D),尤其是光适应下非光化荧光淬灭NPQ_Ln至光稳态非光化荧光淬灭NPQ_Lss。弱光下,‘京丹1号’曲线均位于雷达图最外侧,上升幅度较高;‘佳西娜’和‘京丹1号’的NPQ_Lss分别比其对照提高43.90%、55.73%,而光稳态光化学淬灭系数qP_Lss分别比其对照降低9.33%、17.77%。光适应下,光化学淬灭qP_Ln到暗弛豫过程中,光化学淬灭qP_Dn均以‘京丹1号’降低幅度大。
弱光处理后,番茄叶片PSⅡ最大的光量子效率Fv/Fm、实际光化学效率QY(即ΦPSⅡ)与NPQ叶绿素荧光成像见图6。Fv/Fm成像图显示,弱光下2个品种叶片边缘有较少蓝色或绿色,但整体与对照基本一致,证明叶片PSⅡ光系统未受到严重破坏;QY图看出,弱光下‘京丹1号’叶片颜色变化较明显,范围较大,而‘佳西娜’受伤害面积较少且主要集中于叶片边缘。NPQ图中,弱光下番茄叶片表面出现少量橘黄色和浅绿色部分,‘京丹1号’面积大于‘佳西娜’,且集中于叶片表面和边缘,说明弱光下‘京丹1号’叶片受影响程度较高。
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6叶片叶绿素荧光成像图
Fig. 6Chlorophyll fluorescence imaging of leaves
2.6 番茄幼苗生长对弱光的响应
弱光下,2个品种的株高、叶面积与最大节间距显著提高,茎粗、壮苗指数和单株干重均降低(表4)。其中,相比各自对照处理,‘佳西娜’和‘京丹1号’的株高分别显著提高94.82%、105.58%(P<0.05),壮苗指数分别显著降低56.25%、72.73%(P<0.05),单株干重分别降低8.33%(P>0.05)、45.00%(P<0.05)。图7可见,弱光下‘京丹1号’变化幅度较大,其受弱光的影响更大。Table 4
表4
表4弱光对番茄生长指标的影响
Table 4
| 处理 Treatment | 株高 Plant height (cm) | 茎粗 Stem diameter (mm) | 叶面积 Leaf area (cm2) | 最大节间距 Maximum pitch spacing (cm) | 单株干重 Dry weight of single plant (g) | 壮苗指数 Healthy index | |
|---|---|---|---|---|---|---|---|
| 佳西娜 Glorioso | 对照 CK | 14.67±0.26d | 4.29±0.09a | 44.36±2.18b | 1.56±0.14d | 0.60±0.03b | 0.176±0.09a |
| 弱光Low light | 28.58±0.62b | 3.93±0.07bc | 49.39±2.16ab | 7.08±0.38b | 0.55±0.09b | 0.077±0.09b | |
| 京丹1号 Jingdan No.1 | 对照 CK | 16.50±0.99c | 4.20±0.17a | 43.99±2.25b | 2.22±0.18c | 0.80±0.09a | 0.175±0.09a |
| 弱光Low light | 33.92±0.66a | 3.72±0.08c | 53.55±2.49a | 10.33±0.56a | 0.44±0.02c | 0.048±0.02b | |
新窗口打开|下载CSV
图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7弱光下番茄生长状况
Fig. 7Growth status of tomato under low light
A:佳西娜 Glorioso;B:京丹1号 Jingdan No. 1
3 讨论
3.1 弱光影响番茄叶片叶绿素含量与叶绿体超微结构
REN等[27]发现弱光使玉米叶片叶绿体结构遭到破坏,叶绿素合成减少。而叶绿素b可以利用弱光下占比较高的蓝紫光,提高叶片捕光能力,以维持光系统稳定[28];其含量减少会导致激发能在光系统与电子传递间的分配不平衡,导致PSⅠ、PSⅡ间不稳定,一般直接表现为对逆境的耐受度降低[29]。有研究表明,弱光下,不结球白菜通过增加Chl.b含量以提高光合作用[2];MENG等[30]对弱光下番茄的研究表明叶片通过降低Chl.a/b,在一定程度上减轻对PSⅠ、PSⅡ的抑制。本试验中,弱光下2个番茄品种叶片总叶绿素含量均降低,但‘佳西娜’降低幅度小于‘京丹1号’,且‘佳西娜’的Chl.b含量较其对照提高6.48%。‘佳西娜’Chl.a/b比值降低,与ALA促进Chl.a向Chl.b转化有关[31],这一现象是‘佳西娜’适应逆境的表现[32]。说明弱光下,荷兰品种‘佳西娜’捕光能力强于‘京丹1号’,这是2个番茄品种弱光耐受性差异的原因之一。研究表明弱光会导致叶绿体结构发生不同程度的变化[10,29]。秦玉芝等[14]认为随着马铃薯对弱光环境的适应,其光合产物运输逐渐协调,叶绿体内淀粉粒积累减少。也有研究表明,相对于中度弱光胁迫,严重弱光会导致花生叶片叶绿体结构发生较大变化,淀粉粒数增多[33]。本试验中,弱光下的‘佳西娜’叶绿体基粒片层增多,垛叠整齐有序,结构清晰,少数叶绿体内含淀粉粒;而‘京丹1号’的基粒片层部分解体、模糊不清,大量淀粉粒积累。推测‘京丹1号’因Pn下降较多影响其无机磷供应,蔗糖在细胞质内外运输受阻,无机磷释放也随之减少,磷酸丙糖将主要用于叶绿体内储存淀粉。大量淀粉粒积累会对类囊体造成一定的机械损伤,并吸附部分碳同化相关酶,降低其酶活性[34]。意味着弱光下荷兰品种‘佳西娜’的光能利用率与环境适应性高于‘京丹1号’。
3.2 弱光影响番茄叶片气孔空间分布与光合气体交换参数
气孔是植株与外界环境进行H2O和CO2交换的重要介质,当植株遭受非生物胁迫后,叶片会通过调整气孔密度、开度或形状来缓解伤害[17,35]。本试验发现,弱光下2个番茄品种叶片的气孔密度及宽度均显著减小,‘佳西娜’比‘京丹1号’下降幅度少,可见‘佳西娜’自我调节能力较强。另外,弱光使2个番茄品种叶片气孔规则分布空间的尺度减小,‘佳西娜’的单个气孔间最小邻域距离L(d)略有降低,但‘京丹1号’的L(d)较其对照提高,说明弱光下‘佳西娜’的气孔空间分布比‘京丹1号’更规则;且Pn、Gs降低的同时Ci升高,CO2气体更易通过气孔扩散进入组织内部,影响光合作用的最大反应速率[36,37]。这是荷兰品种‘佳西娜’比‘京丹1号’耐弱光能力强的一个主要原因。3.3 弱光影响番茄叶片叶绿素荧光参数与荧光成像
QY_max(即Fv/Fm)反映了暗适应下PSⅡ反应中心光能转化效率,可作为判断植物对逆境耐受程度的指标[38]。QY(即ΦPSⅡ)为PSⅡ反应中心电荷分离实际量子效率,NPQ反映PSⅡ吸收的光能不用于光化学反应,而以热能耗散掉的部分[39]。本研究发现光照对QY_max影响不大,与荧光显示结果一致,说明叶片PSⅡ没有受到严重破坏。弱光下2个番茄品种的QY_Ln、qP_Ls、qP_Ln与qP_Dn下降,NPQ_Lss、NPQ_Ln和NPQ_Dn升高,表明弱光下植株通过热耗散消耗光合色素吸收的光能增多[40],光合中心有所损伤[41],可能会对RuBP再生产生抑制,但‘佳西娜’变化幅度显著低于‘京丹1号’。因此,弱光下相比‘京丹1号’,‘佳西娜’的天线色素吸收的激发能用于光化学传递的份额较多,热耗散比例少,反应中心开放比例和实际光化学速率较高,与其Chl.b含量高于‘京丹1号’的结果一致。通过叶绿素荧光成像图,发现‘佳西娜’叶片光合系统遭受损害主要位于叶片边缘,‘京丹1号’则于叶片表面大面积受影响,因此‘京丹1号’光合活性降低更多。这是荷兰品种‘佳西娜’对弱光抵御能力与适应能力较强的重要原因之一。3.4 弱光影响番茄幼苗生长状况与SOD活性、MDA含量
植物处于逆境下,体内ROS增多,植物膜透性增大,膜脂肪酸中的不饱和键形成MDA[11];SOD可将ROS转化成H2O2,通过酶促系统减轻膜伤害,以保护植株[42]。本试验中,弱光下‘佳西娜’叶片的SOD活性降低较少,MDA含量低,膜脂过氧化程度较轻,而‘京丹1号’叶片过氧化程度严重。说明‘佳西娜’以提高SOD活性,来减轻叶绿体膜过氧化程度,提高逆境适应能力,这使荷兰品种‘佳西娜’对弱光耐受性强于‘京丹1号’。程亚娇等[43]认为弱光下,植株通过抑制茎横向生长,增加株高并将更多的光合同化产物转移至茎杆,以保证其营养供应,导致植株徒长。本试验中,弱光下2个番茄品种株也出现徒长现象,‘京丹1号’光合产出能力也低于‘佳西娜’。4 结论
弱光下,‘佳西娜’植株变化幅度显著小于‘京丹1号’,其适应弱光环境的能力较强。主要表现在‘佳西娜’叶片Chl.b含量、气孔空间规则分布程度、QY_Lss、qP_Lss与SOD活性高于‘京丹1号’,而NPQ_Lss、叶片受伤害面积及程度低于‘京丹1号’。因此,‘佳西娜’叶片的捕光能力较高,用于光化学传递的份额多,提高了实际光化学效率,保证了光合产物的输出平衡;且‘佳西娜’的叶绿体基粒片层垛叠整齐,光能转化效率较高,植株干重显著高于‘京丹1号’。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/j.plaphy.2016.06.021URLPMID:27362298 [本文引用: 1]

In the present study, we aimed to investigate the effects of exogenous 24-epibrassinolide (EBR) on growth, photosynthetic characteristics, chlorophyll fluorescence imaging, and nitrogen metabolism of tomato leaves under low temperature and weak light conditions. The results showed that foliar application of EBR significantly alleviated the inhibition of plant growth, and increased the fresh and dry weights of tomato plants under a combined low temperature and weak light stress. Moreover, EBR also increased the net photosynthetic rate (Pn), light saturation point (LSP), maximal quantum yield of PSII photochemistry (Fv/Fm), actual photochemical efficiency of PSII (capital EF, CyrillicPSII), and photochemical quenching coefficient (qP), but decreased the intercellular CO2 concentration (Ci), light compensation point (LCP) and apparent quantum efficiency (AQE) under low temperature and weak light conditions. In addition, application of EBR to tomato leaves significantly enhanced the activities of nitrate reductase (NR), glutamate dehydrogenase (GDH), glutamine synthetase (GS), and glutamate synthase (GOGAT), but decreased the ammonium content and nitrite reductase (NiR) activity. We observed that EBR remarkably increased the contents of aspartic acid, threonine, serine, glycine, and phenylalanine, while decreasing the accumulation of cysteine, methionine, arginine, and proline under a combined low temperature and light stress. These results suggest that EBR could alleviate the combined stress-induced harmful effects on photosynthesis and nitrogen metabolism, thus leading to improved plant growth.
DOI:10.1371/journal.pone.0179305URLPMID:28609452 [本文引用: 2]
Anthocyanins are secondary metabolites that contribute to red, blue, and purple colors in plants and are affected by light, but the effects of low light on the physiological responses of purple pak-choi plant leaves are still unclear. In this study, purple pak-choi seedlings were exposed to low light by shading with white gauze and black shading in a phytotron. The responses in terms of photosynthetic properties, carbohydrate metabolism, antioxidant enzyme activity, anthocyanin biosynthetic enzyme activity, and the relative chlorophyll and anthocyanin content of leaves were measured. The results showed that chlorophyll b, intracellular CO2 content, stomatal conductance and antioxidant activities of guaiacol peroxidase, catalase and superoxide dismutase transiently increased in the shade treatments at 5 d. The malondialdehyde content also increased under low light stress, which damages plant cells. With the extension of shading time (at 15 d), the relative chlorophyll a, anthocyanin and soluble protein contents, net photosynthetic rate, transpiration rate, stomata conductance, antioxidant enzyme activities, and activities of four anthocyanin biosynthetic enzymes decreased significantly. Thus, at the early stage of low light treatment, the chlorophyll b content increased to improve photosynthesis. When the low light treatment was extended, antioxidant enzyme activity and the activity of anthocyanin biosynthesis enzymes were inhibited, causing the purple pak-choi seedlings to fade from purple to green. This study provides valuable information for further deciphering genetic mechanisms and improving agronomic traits in purple pak-choi under optimal light requirements.
[本文引用: 3]
[本文引用: 3]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2015.17.017URL [本文引用: 1]
近年来中国设施园艺产业发展迅速,在农产品供应、农民增收和农村经济繁荣方面发挥了重要作用,在作物栽培管理技术、环境控制技术和温室工程技术及配套装备等方面也取得较迅速的发展。文章概述了国内外设施园艺的发展概况与趋势,从设施装备、科技含量、连作障碍等方面分析了目前中国设施园艺存在的主要问题。根据实际情况对中国设施园艺的持续发展提出了相关的重点研究方向,要加强温室结构装备和环境精确控制技术、无土栽培技术、病虫害可持续控制技术、连作障碍克服技术、设施园艺专业品种培育技术等的研究。并从加强政策宏观调控,加大科技创新投入力度,加强设施装备标准化发展,完善园艺产品供求预警及构建质量安全控制长效机制等方面提出了产业化发展措施与建议。
DOI:10.3864/j.issn.0578-1752.2015.17.017URL [本文引用: 1]
近年来中国设施园艺产业发展迅速,在农产品供应、农民增收和农村经济繁荣方面发挥了重要作用,在作物栽培管理技术、环境控制技术和温室工程技术及配套装备等方面也取得较迅速的发展。文章概述了国内外设施园艺的发展概况与趋势,从设施装备、科技含量、连作障碍等方面分析了目前中国设施园艺存在的主要问题。根据实际情况对中国设施园艺的持续发展提出了相关的重点研究方向,要加强温室结构装备和环境精确控制技术、无土栽培技术、病虫害可持续控制技术、连作障碍克服技术、设施园艺专业品种培育技术等的研究。并从加强政策宏观调控,加大科技创新投入力度,加强设施装备标准化发展,完善园艺产品供求预警及构建质量安全控制长效机制等方面提出了产业化发展措施与建议。
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URLPMID:15647315 [本文引用: 1]
URL [本文引用: 2]
URL [本文引用: 2]
[本文引用: 2]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 2]
URL [本文引用: 2]
URLPMID:10648202 [本文引用: 1]
URLPMID:24101997 [本文引用: 2]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
The chloroplast structural alteration and the photosynthetic apparatus activity of cherry tomato seedlings were investigated under dysprosium lamp [white light control (C)] and six light-emitting diode (LED) light treatments designated as red (R), blue (B), orange (O), green (G), red and blue (RB), and red, blue, and green (RBG) with the same photosynthetic photon flux density (PPFD) (approximate to 320 mu mol.m(-2).s(-1)) for 30 days. Compared with C treatment, net photosynthesis of cherry tomato leaves was increased significantly under the light treatments of B, RB, and RBG and reduced under R, O, and G. Chloroplasts of the leaves under the RB treatment were rich. in grana and starch granules. Moreover, chloroplasts in leaves under RB seemed to be a distinct boundary between granathylakoid and stromathylakoid. Granathylakoid under treatment B developed normally, but the chloroplasts had few starch granules. Chloroplasts under RBG were similar to those under C. Chloroplasts under R and G were relatively rich in starch granules. However, the distinction between granathylakoid and stromathylakoid under R and G was obscure. Chloroplasts under 0 were dysplastic. Palisade tissue cells in leaves under RB were especially well-developed and spongy tissue cells under the same treatment were localized in an orderly fashion. However, palisade and spongy tissue cells in leaves under R, O, and G were dysplastic. Stomatal numbers per mm(2) were significantly increased under B, RB, and RBG. The current results suggested blue light seemed to be an essential factor for the growth of cherry tomato plants.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:16668779 [本文引用: 1]
DOI:10.1104/pp.24.1.1URLPMID:16654194 [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URLPMID:27583803 [本文引用: 1]
URLPMID:12758040 [本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:16228557 [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1034/j.1399-3054.2001.1130304.xURL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}