 ,1,2, 王世银,1, 高莉1, 杨力伟1, 邓双义1, 刘晓娜1, 石国庆2, 甘尚权,2
,1,2, 王世银,1, 高莉1, 杨力伟1, 邓双义1, 刘晓娜1, 石国庆2, 甘尚权,2Investigation of miR-486 Target Genes in Skeletal Muscle of Bashbay Sheep in Different Development Periods
ZHANG Wei,1,2, WANG ShiYin,1, GAO Li1, YANG LiWei1, DENG ShuangYi1, LIU XiaoNa1, SHI GuoQing2, GAN ShangQuan,2通讯作者:
责任编辑: 林鉴非
收稿日期:2020-06-3接受日期:2020-10-30
| 基金资助: |
Received:2020-06-3Accepted:2020-10-30
作者简介 About authors
张伟,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (1826KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
张伟, 王世银, 高莉, 杨力伟, 邓双义, 刘晓娜, 石国庆, 甘尚权. 巴什拜羊不同发育阶段骨骼肌中miR-486靶基因的研究[J]. 中国农业科学, 2021, 54(14): 3134-3148 doi:10.3864/j.issn.0578-1752.2021.14.018
ZHANG Wei, WANG ShiYin, GAO Li, YANG LiWei, DENG ShuangYi, LIU XiaoNa, SHI GuoQing, GAN ShangQuan.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】巴什拜羊(Ovis aries)是我国一个优良的肉脂兼用型地方绵羊品种,以生产优质羔羊肉而著称,原产于新疆塔城地区裕民县,现主要分布在塔城地区的裕民县、额敏县和托里县等地,2014年存栏量约150万只[1],该品种羊的养殖一直是当地牧民收入的主要来源。巴什拜羊素有贵族羊之称,其优良的肉质性状在国内外绵羊品种中亦不多见[2]。但是,近年来随着人们健康意识的提高以及肉食品消费观念的转变,低脂肪羊肉的需求量在逐渐增加。巴什拜羊由于胴体脂肪含量较高[3],已无法满足市场的需求,亟需进行遗传改良[4]。本研究对巴什拜羊不同发育阶段骨骼肌中miR-486的靶基因进行深入研究,将为最终阐明巴什拜羊优良肉质性状形成的分子调控机制,进而指导其科学选育奠定基础。【前人研究进展】miRNA(microRNA)是一类小分子非编码RNA,约为22个核苷酸大小,在骨骼肌的发育过程中发挥着重要的调控作用[5,6]。miR-486首先是从人胚胎肝组织中鉴定出来,在心肌和骨骼肌中高表达,ALEXANDER 等发现miR-486作为PTEN/AKT 途径中重要的调节因子在抗肌萎缩蛋白缺陷型肌肉和杜氏肌肉营养不良症中(duchenne muscular dystrophy, DMD)发挥着重要的作用[7]。miR-486在myostatin敲除小鼠的骨骼肌中显著上调,体外过表达miR-486 可以诱导肌管肥大,抑制miR-486可以降低AKT 的活性,说明miR-486是骨骼肌中连接MSTN信号和IGF-1/Akt/ mTOR途径重要的中间调控因子[8]。目前对miR-486的研究主要集中在人类肌肉相关疾病方面,在绵羊等家畜肌肉发育中的相关研究鲜有报道。【本研究切入点】本课题组前期利用高通量测序技术对巴什拜羊不同发育阶段骨骼肌中miRNA的差异表达进行了研究,发现miR-486在出生后巴什拜羊骨骼肌中的表达量极显著上调,为胎儿期的100多倍[9]。进一步研究表明miR-486在巴什拜羊100 d胎儿骨骼肌和心肌中的表达量显著高于胃肠组织(P<0.05),而极显著高于肝、脾和肺等其他组织(P<0.01)[10],提示miR-486在巴什拜羊骨骼肌发育中可能发挥着重要作用。miRNA通过调控其靶基因而发挥其生物学功能,但miR-486在巴什拜羊骨骼肌中的靶基因尚不清楚。【拟解决的关键问题】本研究通过构建miR-486靶基因cDNA文库结合高通量测序,对巴什拜羊胎儿期和出生后骨骼肌中miR-486的靶基因进行深入研究,并对部分靶基因进行试验验证,将为阐明miR-486在巴什拜羊骨骼肌发育过程中的调控机制奠定基础。1 材料与方法
1.1 实验动物及骨骼肌样品采集
试验用巴什拜羊购自裕民县巴什拜种羊繁育中心。试验羊的饲养、前期处理及样品采集工作于2017年8月至2018年2月在新疆雪羚生物科技有限责任公司种羊场完成。选择20只体况良好,胎次在2—3胎,来自不同家系的巴什拜羊(Ovis aries)母羊,统一进行饲养管理。15 d后进行同期发情和人工授精处理。从输精后的第15天开始每天分别在早上10点和下午6点观察母羊的发情状态,剔除返情的母羊,结合B超检查最终选择10只妊娠母羊进入后续实验。分别在妊娠期的第40,50,60,80,100和120天屠宰1头妊娠母羊,采集胎儿的四肢骨骼肌组织并投入液氮速冻,然后转入-80 ℃冰箱保存备用。其余4只母羊继续饲养至羔羊出生,并于出生当天,及出生后30,60和90 d屠宰羔羊,按照上述方法采集和保存骨骼肌组织。
1.2 RNA提取及基因组DNA消化
参照TRIzol(Invitrogen, USA)说明从上述骨骼肌样品中提取总RNA,使用Bioanalyzer 2100(Agilent, Germany)评估总RNA完整性。参照Oligotex®mRNA纯化试剂盒(Promega, USA)的说明,从总RNA中分离纯化mRNA,然后使用RNase-free DNase I(Ambion Inc., USA)消化所得mRNA,以去除微量基因组DNA污染,避免其对cDNA文库质量的影响。1.3 cDNA文库的构建和高通量测序
将胚胎期6个阶段骨骼肌mRNA混合为一个样品,出生后4个阶段骨骼肌mRNA混合为另一个样品,参照甘尚权等[11]建立的方法分别构建胎儿期和出生后巴什拜羊骨骼肌miR-486的靶基因cDNA文库。现以胎儿期cDNA文库为例,简要说明建库的过程。将胎儿期骨骼肌mRNA混合样品分为两份,分别用反转录引物A(5°-3°: CTCGACTGAGTTGCCGTGAGTCGG CAACTCAGTCGAGGTCATGTCCT,斜体加粗碱基为巢式引物的上游引物序列,下划线所示碱基为miR-486与其靶基因结合的种子序列)和B(5°-3°: AACGCGTCGCGTCGAGTAGTAGACGTA+oligodT20,下划线所示碱基为巢式引物的下游引物互补序列),参照PrimeScript™ Double Strand cDNA Synthesis Kit(TAKARA)试剂盒操作说明合成cDNA双链,分别称之为A库和B库。由于用于构建A库的反转录引物3°末端的10个碱基为miR-486与其靶基因结合的种子序列,只有该序列与mRNA上的序列互补结合,才能完成反转录,所以A库中的cDNA应主要为miR-486的靶基因序列;用于构建B库的反转录引物3°末端是20个oligodT,所以B库中的cDNA应是相应骨骼肌组织中所有表达功能基因序列的集合。使用酚/氯仿法抽提纯化A库和B库的cDNA双链后将二者等体积混合,95 ℃变性5 min,然后50 ℃保温5 min形成A库和B库的杂交cDNA文库。最后以杂交的cDNA为模板,使用巢式引物(上游引物5°-3°: TGAGTCGGCAACTCAGTCGAG和下游引物5°-3°: AGCGCAGCTCATCATCTGCAT)进行巢式PCR扩增。此时,只有A库中miR-486的靶基因链与其在B库中对应的互补链形成的杂交双链同时带有上下游巢式引物,所以在巢式PCR反应中可以被指数级扩增。而其他形式的杂交双链只带有上游或者下游巢式引物,在巢式PCR反应中只能被线性扩增。所以,经巢式PCR扩增后的产物,绝大部分为miR-486的靶基因序列,本研究中称之为miR-486靶基因cDNA文库。将构建的两个cDNA文库分别采用Illumina HiSeq 3000测序平台进行双向高通量测序。1.4 高通量测序数据处理
测序原始数据使用SOAPnuke过滤,除去接头序列以及未知碱基超过10%和低质量碱基(Q≤10)超过50%的低质量reads。获得的clean reads使用SOAPaligner和SOAP2以牛(Bos taurus v3.1)和绵羊(Ovis aries v3.1)基因组序列为参考进行拼接组装和注释。1.5 候选靶基因序列筛选及分析
按照靶基因cDNA文库建库原理,所得miR-486靶基因序列应该含有反转录引物A的末端10碱基序列(GTCATGTCCT)。因此以该10碱基序列为种子序列对上述分析获得的基因序列进行比对筛选,含有该序列的基因序列即为miR-486靶基因序列,由于所得靶基因尚未经过实验验证,存在一定数量的假阳性结果,所以暂时称为候选靶基因。为了深入了解所得候选靶基因的生物学功能,利用David在线软件进行基因本体论[12](gene ontology, GO)(1.6 部分候选靶基因在巴什拜羊不同发育阶段骨骼肌中表达量检测
在以上靶基因cDNA文库测序结果分析的基础上,查阅近年来骨骼肌发育相关信号通路的文献报道,从靶基因cDNA文库中选择10个与骨骼肌生长发育调控密切相关的候选靶基因,以β-actin为内参基因,以出生后第90天的表达量为对照,使用qRT-PCR对其在巴什拜羊不同发育阶段骨骼肌中的表达量进行相对定量,并参照课题组前期检测的miR-486的表达规律[10],对10个候选靶基因与巴什拜羊骨骼肌发育的关联性,及其与miR-486的靶向调控关系进行初步分析。10个候选靶基因及内参基因在Ensemble数据库(Table 1
表1
表110个候选靶基因qRT-PCR引物信息表
Table 1
| 基因 Gene | 登录号* Transcript ID | 引物名称 Primers name | 引物序列(5′-3′) Sequence of primers | 片段大小 Size (bp) |
|---|---|---|---|---|
| PTEN | ENSOART00000015407.1 | PTENqF | GTATTTGCAGTATAGAGCGTGC | 165 |
| PTENqR | GGATTTGATGACTCCTCTACTG | |||
| Foxo1 | ENSBTAT00000061127.3 | Foxo1qF | CCACAGCAATGACGACTTCGAC | 275 |
| Foxo1qR | GACTGGGTGGACACAGTCAATG | |||
| Dock3 | ENSOART00000008795.1 | Dock3qF | AATCAGCCAAGCCTTCAGCTAG | 246 |
| Dock3qR | TCTGCTCCCAGTCCATCATGTC | |||
| PAX7 | ENSOART00000011524.1 | PAX7qF | GTTCGATTAGCCGAGTGCTCAG | 274 |
| PAX7qR | GTAGATGTCTGGGTAGTGGGTC | |||
| ARHGAP5 | ENSOART00000006997.1 | ARHG5qF | GTCTATCAGAACCATGTACAGC | 189 |
| ARHG5qR | CATACACCTCTTTGCTATCAGC | |||
| OLFM4 | ENSOART00000007293.1 | OLFM4qF | GACCATCTCCAAGAAGTTCGAG | 159 |
| OLFM4qR | CTTGATCAGCTCAAAGTCCAGC | |||
| IGF1R | ENSOART00000010895.1 | IGF1RqF | GGAAGAGCTCGAGACTGAGTAC | 159 |
| IGF1RqR | GACAAAGTTAGAGGCGCTGCAG | |||
| PIK3R1 | ENSOART00000006390.1 | PIK3R1qF | CATGGGGATTACACTCTTACAC | 147 |
| PIK3R1qR | CTAGAGATTCATTCCGGTAGTG | |||
| FBN1 | ENSOART00000022901.1 | FBN1qF | GGATTTCACGTCACACGAGATG | 208 |
| FBN1qR | ACACAACGCCCATTCATGCAGA | |||
| PDCD4 | ENSOART00000011353.1 | PDCD4qF | ATTGCTAGAGCTGTTGGAGATG | 250 |
| PDCD4qR | TCAGCTTCAGAAATGTCTCCAG | |||
| β-actin | ENSOART00000003275.1 | β-actin qF | TGTGCGTGACATCAAGGAGAAG | 177 |
| β-actinqR | AGGAAGGAAGGCTGGAAGAG |
新窗口打开|下载CSV
1.7 miR-486与部分候选靶基因靶向调控关系的验证
由于通过构建靶基因文库获取的靶基因存在一定比例的假阳性结果,为了验证上述靶基因文库的可靠性,结合上述10个候选靶基因在巴什拜羊不同发育阶段表达量的变化规律,以及课题组前期检测到的miR-486的表达规律[10],从上述10个候选靶基因中选择PTEN、Foxo1、IGF1R和PIK3R14个表达量与miR-486的表达量呈明显负相关的候选靶基因作进一步验证。1.7.1 双荧光素酶报告载体试验 在候选靶基因序列上miR-486结合靶位点上下游约100 bp片段范围设计PCR引物,以巴什拜羊基因组为模板PCR扩增获得靶位点区域约200 bp大小的野生型片段,同时根据种子序列设计突变引物,与上述PCR引物进行重叠PCR对靶位点进行点突变,获得靶位点处4个碱基突变的突变型片段。将野生型片段和突变型片段经XhoI和HindIII双酶切后连入PGL4双荧光素酶报告载体(Promega, USA),并使用双酶切和测序确认载体构建的正确性。上述载体构建引物信息见表2。
Table 2
表2
表2荧光素酶报告载体构建引物
Table 2
| 引物 Primer | 引物序列(5′-3′) Sequence of primers | 片段大小Size (bp) | 试验用途 Experiment |
|---|---|---|---|
| PTENcF | ccctcgagggCACCTTTCTTTAGCATGCTAC | 262 | 载体构建 Vector construction |
| PTENcR | ccaagcttggGATAGCCTCCACATTTGTATG | ||
| PTENmutF | GAATCTGTATTGGGGTACcttcATGAACCTTCCACAACAT | 262 | 靶位点突变 Mutate bind site |
| PTENmutR | ATGTTGTGGAAGGTTCATgaagGTACCCCAATACAGATTC | ||
| Foxo1cF | ccctcgagggGAGAAGCAGTCCAAAGATGTC | 221 | 载体构建 Vector construction |
| Foxo1cR | ccaagcttggATGGTGTAGTGAGTTTGGCAC | ||
| Foxo1mutF | CGAAGACGCTTCCTGTACcttcTGTTTGCCCAGTGTTTGC | 221 | 靶位点突变 Mutate bind site |
| Foxo1mutR | GCAAACACTGGGCAAACAgaagGTACAGGAAGCGTCTTCG | ||
| IGF1RcF | ccctcgagggGAGAATCCCAATGGATTGATC | 237 | 载体构建 Vector construction |
| IGF1RcR | ccaagcttggGGATGAAGTTCTCATATGTCG | ||
| IGF1RmutF | GTGTCCAGACAGGAGTACcttcAGTATGGAGGAGCCAAGC | 237 | 靶位点突变 Mutate bind site |
| IGF1RmutR | GCTTGGCTCCTCCATACTgaagGTACTCCTGTCTGGACAC | ||
| PIK3R1cF | ccctcgagggCTGAAGGCTAAATTCACAGTG | 267 | 载体构建 Vector construction |
| PIK3R1cR | ccaagcttggTAACAGCCAAGTACTCTGTAC | ||
| PIK3R1mutF | GCTTTTAAAGAAATGTACcttcTGCCAGTTTGTCAAGTCG | 267 | 靶位点突变 Mutate bind site |
| PIK3R1mutR | CGACTTGACAAACTGGCAgaagGTACATTTCTTTAAAAGC |
新窗口打开|下载CSV
参照Lipofectamine 2000(Invitrogen, USA)的操作说明,将构建好的报告载体及miR-486 mimics(表3)分别转入Hela细胞,培养48 h后裂解细胞,按照Dual-Luriferase Reporter Assay System(Promega, USA)的操作说明检测萤火虫荧光素酶和海肾荧光素酶的活性。
1.7.2 骨骼肌卫星细胞中miR-486干扰表达试验 检索miRBase数据库,根据其收录的bta-miR-486序列(MIMAT0009329)设计其模拟物miR-486 mimics,抑制物miR-486 inhibitor及其阴性对照negative control(表3)并由上海吉玛基因合成。扩大培养巴什拜羊骨骼肌卫星细胞(通过原代培养获得),参照Lipofectamine 2000(Invitrogen, USA)的操作说明将miR-486 mimics,miR-486 mimics negative control,miR-486 inhibitor和miR-486 inhibitor negative control分别转入骨骼肌卫星细胞,以不转染任何试剂的细胞组作为空白对照。转染后的细胞继续培养至24、48和72 h时分别消化收集细胞,按照1.2所述方法提取总RNA,并按照1.6所述方法及引物检测不同处理组骨骼肌卫星细胞中4个候选靶基因mRNA的水平。
Table 3
表3
表3miR-486 mimics、miR-486 inhibitor及其阴性对照序列
Table 3
| 名称 Name | 序列(5′-3′) Sequence |
|---|---|
| miR-486 mimics | UCCUGUACUGAGCUGCCCCGAG |
| miR-486 mimics negative control | CAGUACUAGCGUGUAGUACCAA |
| miR-486 inhibitor | CUCGGGGCAGCUCAGUACAGGA |
| miR-486 inhibitor negative control | GCAUGGCUUGAGUCCGUAAGAU |
新窗口打开|下载CSV
1.8 数据分析
qRT-PCR结果采用2-△△Ct法进行计算,双荧光素酶检测结果采用萤火虫荧光素酶活性/海肾荧光素酶活性计算。试验均重复3次,结果用“平均值±标准差”表示,使用SPSS13.0进行单因素方差分析,P<0.05表示差异显著,P<0.01表示差异极显著。2 结果
2.1 靶基因cDNA文库高通量测序及分析
胎儿期和出生后miR-486靶基因cDNA文库高通量测序分别得到6.21M和5.94M原始数据,剔除接头序列以及低质量reads后分别获得5.73M和5.42M的clean reads。进一步拼接组装、剔除冗余序列后分别得到283条和276条Unigenes,平均序列长度分别为673 bp和681 bp。将胎儿期和出生后Unigenes序列进行blastn比对,发现分别有68和61个为胎儿期和出生后特异表达Unigenes,有215个为两个时期的共表达Unigenes。2.2 候选靶基因注释及功能分析
按照靶基因cDNA文库建库原理,所得miR-486候选靶基因序列应该含有反转录引物A的末端10碱基序列(GTCATGTCCT)。因此以该10碱基序列为种子序列对上述分析获得的Unigenes序列进行比对筛选,胎儿期和出生后分别获得123个和118个符合条件的Unigenes,其中96个为两个阶段共表达Unigenes,其余27个和22个分别为胎儿期和出生后骨骼肌中特异表达的Unigenes。由于miR-486与所得Unigenes的靶向调控关系尚未经过试验验证,所以暂时称之为miR-486的候选靶基因。GO分析结果表明,胎儿期和出生后分别有96个和87个候选靶基因得到注释信息,胎儿期的候选靶基因显著富集于26个细胞组分条目,32个生物学过程条目和18个分子功能条目。出生后的候选靶基因显著富集于20个细胞组分条目,36个生物学过程条目和23个分子功能条目,富集基因数前10的GO条目及富集于相应条目的候选靶基因占该阶段检测到候选靶基因的比例见图1。
图1
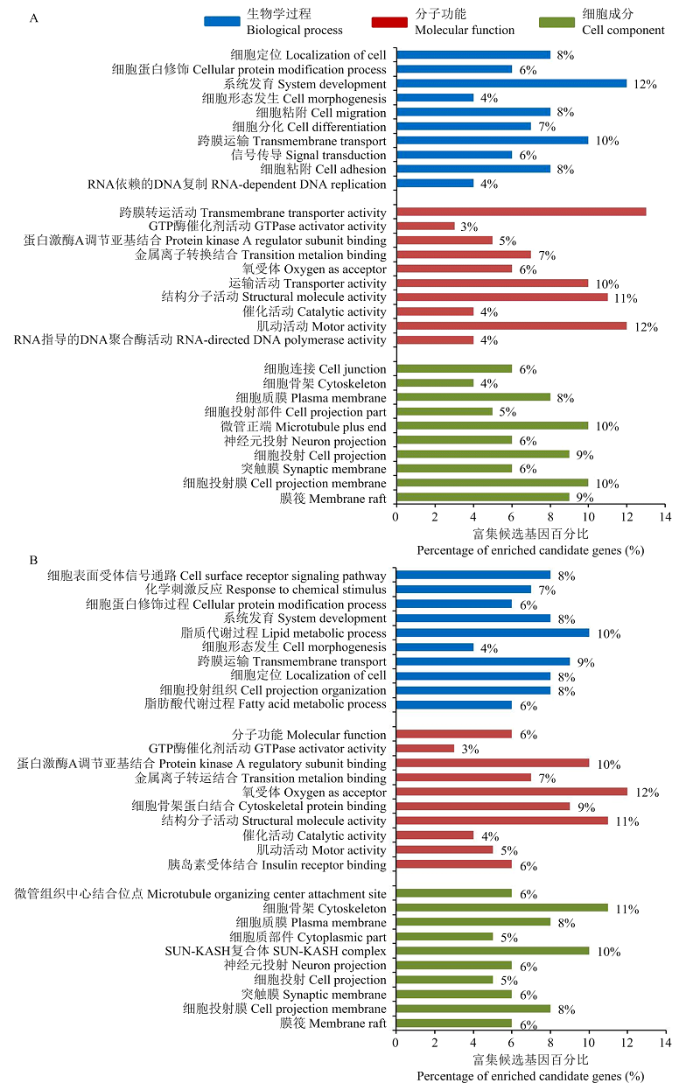 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1候选靶基因GO分析富集基因数前10的GO条目
A和B分别为胎儿期和出生后候选靶基因GO分析结果,Y轴为GO条目名称,X轴为相应条目富集的候选靶基因占注释于该类型候选靶基因的比例
Fig. 1The top 10 items in GO analysis of candidate target genes
A and B are GO analysis results of fetal and postnatal period, y axis show the GO items, X axis show the percentage of the candidate target genes enriched in one GO item account for the candidate target genes noted in one kind of GO mold
KEGG通路富集分析结果表明,胎儿期和出生后候选靶基因显著富集于多个与肌肉发育相关的信号通路,如PI3k-Akt、MAPK、Wnt和Adherens junction等,提示miR-486可能通过调控一些与肌肉发育相关的基因及信号通路,在巴什拜羊骨骼肌发育调控中发挥着重要的作用。胎儿期和出生后候选靶基因显著富集的前20个信号通路见图2。
图2
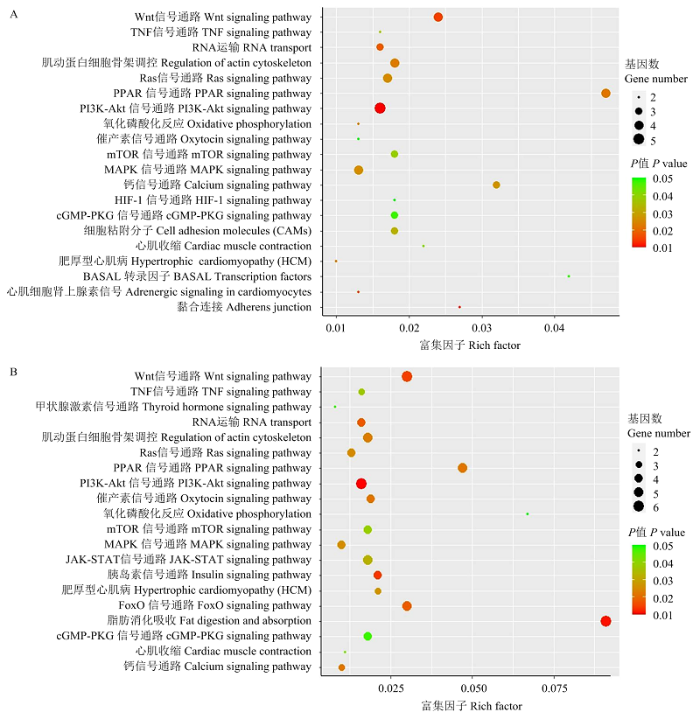 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2候选靶基因的KEGG通路分析
A和B分别为胎儿期和出生后候选靶基因参与通路分析结果,Y轴为pathway名称,X轴为富集因子,气泡大小反映了富集到某个信号通路中的候选靶基因的数量占该通路已发现参与基因的比例,颜色变化反映了某个通路富集的显著性水平
Fig. 2KEGG analysis result of candidate target genes
A and B are KEGG analysis result of fetal and postnatal period candidate target genes, y axis show the pathway name, X axis show the rich factor, bubble size reflect the number of candidate target genes enriched to a certain pathway/all quantities in the background, color reflect the significance level of a certain pathway
2.3 miR-486及其部分候选靶基因在巴什拜羊不同发育阶段骨骼肌中的表达规律
qRT-PCR检测结果表明所选10个候选靶基因在巴什拜羊骨骼肌中均有表达(图3),但在不同发育阶段的表达量存在差异。PTEN表达量在胎儿期呈持续下降趋势,在胎儿期第40和50 天表达量最高,极显著高于其他时期(P<0.01),胎儿期第60天开始逐步下降,至出生后降至最低且保持相对稳定(图3-A)。Foxo1表达量从胎儿期至出生后呈持续下降趋势,在胎儿期第40、50和60天表达量最高且差异不显著(P>0.05),但极显著高于其他时期(P<0.01);胎儿期第80、100和120天时表达量下调,但仍极显著高于出生后各阶段(P<0.01);出生当天至出生后第30天表达量进一步下降,这2个时期差异不显著(P>0.05),但极显著高于出生后第60和90天的表达量(P<0.01),至出生后第90天降至最低(图3-B)。Dock3和PAX7在胎儿期前期高表达,胎儿期第40、50、60和80天均极显著高于其他各阶段(P<0.01),但至胎儿期第100天迅速下调,至出生后第90天一直保持较低的表达水平(图3-C和D)。IGF1R表达量在胎儿期第40、50和60天表达量最高且差异不显著(P>0.05),至胎儿期第80和100天时表达量极显著下调(P<0.01),至胎儿期第120天时又出现极显著上调(P<0.01),但至出生后即出现极显著下调(P<0.01),直至出生后第90天降至最低水平(图3-G)。PIK3R1在胎儿期第40和50天时呈高表达,至胎儿期第60和100天时出现2次极显著下调(P<0.01),而胎儿期第80和120天时出现2次极显著上调(P<0.01),至出生后一直呈现持续下调趋势(图3-H)。FBN1基因在胎儿期第40、50、60和80天表达量最高且差异不显著(P>0.05),胎儿期第100 d时表达量极显著下调(P<0.01),胎儿期第120 d时又出现极显著上调(P<0.01),至出生后再次出现极显著下调(P<0.01),并在出生当天及出生后30、60和90 d保持稳定表达(图3-I)。上述7个基因在巴什拜羊不同发育阶段骨骼肌中的表达变化存在一定差异,但是总体而言呈胎儿期高表达,出生后低表达的趋势。图3
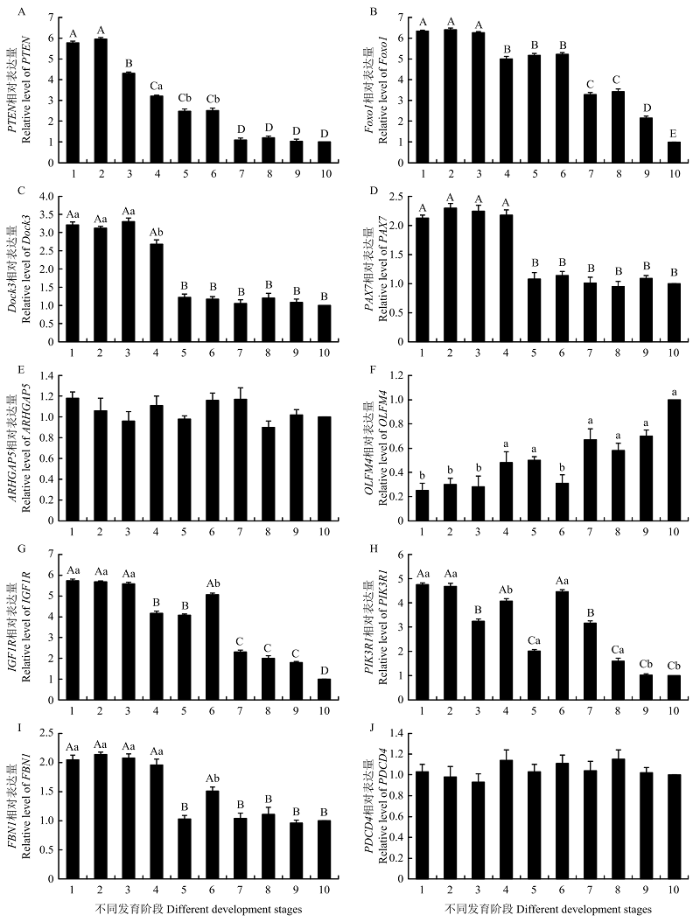 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图310个候选靶基因在巴什拜羊不同发育阶段骨骼肌中的相对表达量
A-J分别为10个候选靶基因在巴什拜羊不同发育阶段骨骼肌中相对表达量的qRT-PCR检测结果,横轴为巴什拜羊骨骼肌不同发育阶段,1-10分别指胎儿期第40、50、60、80、100和120天,以及出生当天,出生后30、60和90 d共10个不同的发育阶段,纵轴为相应基因的相对表达量,图中相同字母或无字母表示差异不显著(P>0.05),不同小写字母表示差异显著(P<0.05),不同大写字母表示差异极显著(P<0.01)。下同
Fig. 3The relative expression levels of 10 candidate target genes in skeletal muscle of Bashbay sheep in different development stages
Figure A to J are qRT-PCR results of 10 candidate target genes in skeletal muscle of Bashbay sheep in different development stages, the abscissa axis shows the different development stages of Bashbay skeletal muscle, 1 to 10 shows the 40, 50, 60, 80, 100 and 120 d of fetus periods, and the day of birth, and 30, 60 and 90 d of postnatal periods, expression levels without letters or with the same letters have no significant difference (P>0.05), with the different lowercase letters have significant differences (P<0.05), with the different capital letters have extremely significant differences (P<0.01). The same as below
已有的研究结果表明miRNA对其靶基因的调控一般为负调控,而课题组前期研究发现miR-486在巴什拜羊胎儿期总体呈低表达,出生后表达量逐渐上调[10]。由此可以推断,在上述7个候选靶基因与miR-486巴什拜羊骨骼肌中存在靶向调控关系的可能性很大。
ARHGAP5和PDCD4在胎儿期和出生后巴什拜羊骨骼肌中的表达量未见显著变化(图3-E和3-J),推测这2个基因的表达可能是巴什拜羊骨骼肌生长发育所必需,故其表达量保持相对稳定;而OLFM4胎儿期呈低表达,但表达量逐步上调,至出生后第90 天达到较高的表达水平(图3-F)。结合miRNA与其靶基因靶向调控的规律,推测这3个候选靶基因与miR-486存在靶向调控关系的可能性不大。
2.4 荧光素酶报告载体验证
为了进一步验证miR-486与候选靶基因的靶向调控关系,从上述10个候选靶基因中选择PTEN、Foxo1、IGF1R和PIK3R1 4个候选靶基因进行双荧光素酶报告载体实验验证。将4个候选靶基因野生型(WT)和突变型(Mut)靶位点序列插入到双荧光素酶报告载体海肾荧光素酶和萤火虫荧光素酶之间(图4-A),荧光素酶活性检测结果表明miR-486与插入野生型靶位点序列报告载体共转染细胞中萤火虫荧光素酶的活性均极显著低于对照组(P<0.01),而miR-486与插入突变型靶位点序列报告载体共转染细胞中萤火虫荧光素酶的活性与对照组无显著差异(P>0.05,图4-B),说明miR-486可与候选靶基因靶位点序列高效结合,从而抑制其后萤火虫荧光素酶的表达,由此也初步证明miR-486与PTEN、Foxo1、IGF1R和PIK3R1 4个基因存在靶向调控关系。图4
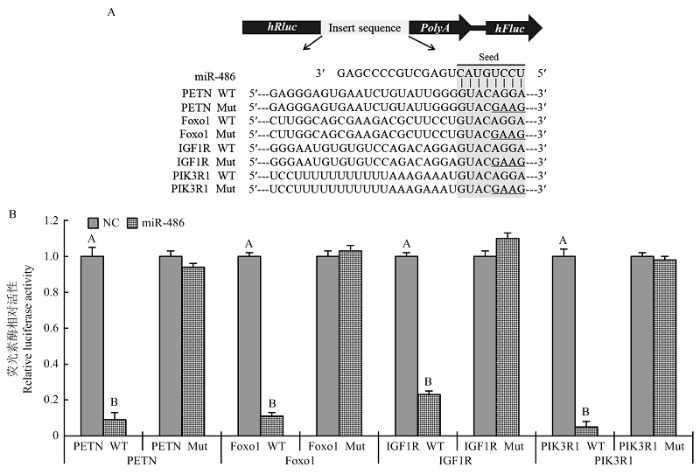 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4候选靶基因荧光素酶报告载体试验
A为候选靶基因片段插入双荧光素酶报告载体位置及4个候选靶基因野生型(WT)和突变型(Mut)靶位点序列,序列中带下划线的碱基为通过重叠PCR获得的突变碱基;B为插入野生型和突变型靶位点片段荧光素酶报告载体的荧光素酶相对活性检测结果,NC指miR-486的阴性对照(Negative control)
Fig. 4Double luciferase reporter assay experiments
A: the insert site of candidate target gene fragment in vector and the sequence of wild (WT) and mutant (Mut) type, the base with underline are mutated bases achieved by overlap extension PCR. B: the relative luciferase activity determined 48 h after transfection. Data are expressed as means ± SD of three independent experiments, NC is negative control of miR-486
2.5 骨骼肌卫星细胞中靶向调控关系验证
巴什拜羊骨骼肌卫星细胞中的验证结果表明,对于PTEN和IGF1R,当转入miR-486 mimics造成细胞中miR-486的表达水平呈上调状态时,继续培养至24、48和72 h的细胞中均检测到PTEN和IGF1R的mRNA水平随着培养时间的延长呈显著下调趋势(P<0.05),空白对照组、miR-486 inhibitor组和两个阴性对照组细胞中PTEN和IGF1R的mRNA水平未见显著变化(P>0.05)(图5-A和C);PIK3R1 的mRNA水平在培养至24 h细胞中未见显著变化(P>0.05),48 和72 h的mRNA水平变化与PTEN和IGF1R相同(图5-D);Foxo1 mRNA水平在miR-486 inhibitor组24、48和72 h的细胞中均呈显著上调趋势(P<0.05),在miR-486 mimics组48和72 h的细胞中均呈显著下调趋势(P<0.05),其余处理组之间无显著差异(P>0.05)(图5-B)。图5
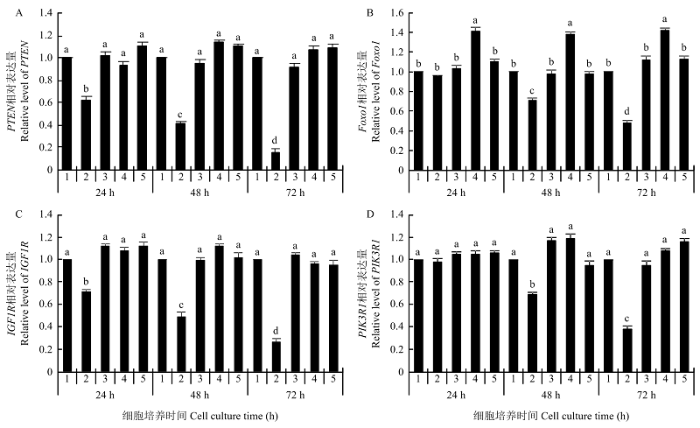 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5转染细胞中4个候选靶基因mRNA水平变化
A、B、C和D分别为候选靶基因PTEN、Foxo1、IGF1R和PIK3R1在转染细胞中24、48和72 h的相对表达量
Fig. 5The relative expression levels of 4 candidate target genes in transfected cells
A, B, C and D are relative expression levels of 4 candidate target genes in transfected cells during 24, 48 and 72 h
综合荧光素酶报告载体实验和骨骼肌卫星细胞中靶向调控关系验证实验结果,可以确认在巴什拜羊骨骼肌中miR-486可以靶向调控PTEN、Foxo1、IGF1R和PIK3R1 4个基因。
3 讨论
动物的骨骼肌是由单核的成肌细胞相互融合而成的多核肌纤维构成的[14]。成肌细胞来源于中胚层,迁移至躯干和四肢等骨骼肌发育的位置后,成肌细胞先增殖达到一定的数量,然后相互融合为肌管,并最终发育成肌纤维[15,16],这一过程一直持续至动物出生前才结束。在不同的动物中,肌管的形成可以明显的区分为两至三个时期。就大部分动物而言,第一波和第二波肌管分别来自于胚胎和胎儿成肌细胞,并分别发育成初级和次级肌纤维[15]。但在一些大型动物中,例如牛、绵羊、猪和人,在妊娠后期还会有第三波肌管形成[17,18,19,20],这可能与这些动物具有发达的骨骼肌有一定的关系。利用光学显微镜定量研究绵羊骨骼肌的超微结构,发现胎儿期的40—80 d和100—120 d为骨骼肌生长发育的关键时期[17]。李雪娇等对中国美利奴羊骨骼肌发育进行了系统研究,发现胎儿期第75天前后肌纤维的形成基本完成,至第105天时肌纤维的数量基本恒定,此后骨骼肌的发育主要表现为肌纤维直径和长度的增加[21,22]。石田培等利用全转录组测序对中国美利奴羊胎儿期骨骼肌中circRNA的表达情况进行研究,发现胎儿期第85、105和135天这3个阶段显著富集了大量与肌肉发育相关的基因和通路[23]。提示这些阶段骨骼肌中的基因表达模式发生了变化,以支持骨骼肌细胞不同生长发育状态的转换和维持。鉴于此,本研究进一步细化采样阶段,对巴什拜羊胎儿期第40、50、60、80、100和120天,以及出生当天,出生后第30,60和90天骨骼肌中miR-486的靶基因进行深入挖掘,发现有96个候选靶基因在胎儿期和出生后均有表达,有27个和22个候选靶基因分别在胎儿期和出生后特异表达。对其中10个候选靶基因mRNA水平的定量研究结果表明,即使是共表达的候选靶基因,在巴什拜羊不同发育阶段骨骼肌中的表达量亦有很大的变化,PTEN、Foxo1和Dock3等7个候选靶基因在胎儿期巴什拜羊骨骼肌中呈高表达,出生后其表达量显著下调,而OLFM4在胎儿期低表达,出生后表达量出现显著上调,仅有ARHGAP5和PDCD4两个候选靶基因的表达量未见显著变化。结合前人研究结果,推测这些候选靶基因可能在维持不同发育阶段骨骼肌细胞增殖和分化过程中发挥着重要的作用。miR-486由Ankyrin-1(Ank-1)的第40个内含子编码[24],Ank-1第40—42外显子编码的一个小分子Ank1蛋白仅在肌肉中表达,可将肌小节连接在肌质网上[25]。到目前为止未发现miR-486尚有其他家族成员,且仅在哺乳动物中表达,其序列高度保守,在鸟类、鱼类、两栖类和更低等的后生动物中均未检测到miR-486的表达[24]。miR-486在巴什拜羊的骨骼肌和心肌中高表达[10],且其表达量在出生后巴什拜羊骨骼肌中极显著上调[9],推测miR-486在巴什拜羊骨骼肌发育过程中可能发挥着重要的作用,本研究中发现的miR-486部分候选靶基因已证明与骨骼肌发育调控和机能维持密切相关,初步支持这一推测的可靠性。PTEN(phosphatase and tensin homolog)是一个重要的肿瘤抑制基因,可以通过抑制PI3K的活性以及降低Akt的磷酸化水平影响下游mTOR信号通路,最终影响细胞的生长和存活[26]。在成年人的骨骼肌中,骨骼肌卫星细胞处于静止、激活和分化的动态平衡状态,但是当PTEN被敲除,大量的骨骼肌卫星细胞将不经过增殖即进入激活、提前成熟分化状态,进而造成骨骼 肌中卫星细胞的消耗而最终影响骨骼肌损伤的修复和再生[27]。Foxo1可以促进小鼠肌细胞融合,同时参与骨骼肌纤维分化的调控,在富含快肌纤维的骨骼肌组织中的表达量显著高于富含慢肌纤维的骨骼肌组织[28,29]。在小鼠肌细胞中,miR-486抑制其靶基因PTEN和Foxo1的表达进而激活PI3K/Akt信号通路参与肌肉的生长的调控和内稳态的维持[24]。在杜氏肌应用不良症(duchenne muscular dystrophy, DMD)患者的肌肉组织中miR-486的表达量显著下调,同时在DMD动物模型骨骼肌中过表达miR-486,可降低DOCK3和PTEN的水平,进而提高AKT的磷酸化水平并使肌营养不良症状得到明显缓解[30]。慢性肾病(chronic kidney disease, CKD)患者由于E3连接酶的激活,泛素蛋白酶体系统被活化,进而引起肌肉中蛋白质的分解,但是若上调肌肉中miR-486的水平可显著抑制E3连接酶的表达,使患者的肌肉量明显增多,进一步研究表明这一过程是miR-486通过抑制Foxo1的转录,同时增加Foxo1磷酸化水平,降低PTEN磷酸化水平,最终使Akt的磷酸化水平显著升高进而激活相关信号通路而实现[31,32]。本研究已证实,在巴什拜羊骨骼肌中miR-486可以靶向调控PTEN、Foxo1、IGF1R和PIK3R1 4个基因的表达,但其在巴什拜羊骨骼肌发育过程中的生物学功能尚有待后续试验阐明。
KEGG分析结果表明,miR-486通过调控其候选靶基因的表达同时参与多个信号通路的调控。例如IGF1R同时参与mTOR signaling pathway、PI3K-Akt signaling pathway、FoxO signaling pathway、MAPK signaling pathway、Ras signaling pathway 和Adherens junction等信号通路的调控,PIK3R1同时参与Phosphatidylinositol signaling system、PI3K-Akt signaling pathway、FoxO signaling pathway、mTOR signaling pathway和Insulin signaling pathway等信号通路的调控,很多信号通路已证明与骨骼肌生长发育的调控密切相关[33,34,35,36,37],在小鼠中的研究结果表明,其骨骼肌中miR-486的表达受myostatin的调控,通过miR-486转录水平的调控可将myostatin信号通路和IGF-1/Akt/ mTOR信号通路有效整合[8]。在人类骨骼肌中,miR-486可通过调控PI3K-Akt、ubiquitin proteasome、FoxO和mTOR信号通路,使骨骼肌组织很好地适应有氧运动训练[38,39],提示miR-486可能在沟通和协调巴什拜羊骨骼肌内相关信号通路中发挥着重要作用。
在细胞质中miRNA首先与Argonaute蛋白组装成为miRNA诱导的沉默复合体(miRISC),然后miRNA通过其种子序列与靶基因mRNA互补结合引导miRISC结合到其靶基因mRNA的靶位点上,进而调控其靶基因的翻译[40,41],但在动物中miRNA是通过抑制其靶基因mRNA的翻译还是诱导其降解实现其对靶基因的调控尚存在争议,有研究结果表明在动物细胞中miRNA主要是抑制其靶基因mRNA的翻译,并不会诱导mRNA的降解[42],但也有相关研究检测到了miRNA诱导的mRNA降解[43]。或许DJURANOVIC等的研究可以更好地解释这一问题,他们利用果蝇S2细胞建立了一个可控的表达系统,并以此研究了miRNA对其靶基因调控的动力学特征,发现miRNA不论跟天然的或者人工合成的3′ UTR区靶位点结合,首先是抑制其翻译,进而引起mRNA的脱腺苷化和降解,所以miRNA可能是先抑制其靶基因mRNA的翻译,然后再诱导其降解来实现其对靶基因的调控,若在抑制阶段检测细胞中mRNA的水平,则会得出miRNA不会诱导mRNA降解的结论[44,45]。本研究中上调巴什拜羊骨骼肌卫星细胞中miR-486的水平,至24 h其靶基因Foxo1和PIK3R1的mRNA水平未见显著降低(P>0.05),但是至48和72 h时Foxo1和PIK3R1的mRNA水平显著低于对照组(P<0.05),与此不同的是PTEN和IGF1R的mRNA水平至24 h时即可检测到显著降低(P<0.05)。同为miR-486的靶基因,其调控方式却存在不同,其机理尚有待进一步研究。通过靶位点分析发现,miR-486在PTEN的mRNA上有3个靶位点,其中一个位于CDS区,在IGF1R的mRNA上有2个靶位点,而在Foxo1和PIK3R1的mRNA上仅检测到1个靶位点,这种调控方式的差异是否与靶位点的位置和数量有关尚有待后续实验验证。另外,当下调巴什拜羊骨骼肌卫星细胞中miR-486时,仅检测到Foxo1的mRNA水平较对照组显著升高,其他3个靶基因mRNA水平未见显著变化(P>0.05),其机理亦有待进一步阐明。
4 结论
本研究采用构建cDNA文库结合高通量测序的方法首次对巴什拜羊不同发育阶段骨骼肌中miR-486的靶基因进行了深入研究,在胎儿期和出生后骨骼肌中分别检测到123个和118个miR-486的候选靶基因,功能分析结果表明候选靶基因显著富集于PI3k-Akt、Wnt和MAPK等已证明与肌肉发育调控密切相关的信号通路。其中PTEN、Foxo1、Dock3、PAX7、IGF1R、PIK3I1和FBN1 7个候选靶基因在胎儿期巴什拜羊骨骼肌中呈高表达,出生后其表达量显著下调,而候选靶基因OLFM4呈相反的表达趋势,提示这些基因在巴什拜羊胎儿期和出生后骨骼肌发育状态转换中可能发挥着重要作用。选择PTEN、Foxo1、IGF1R和PIK3R1 4个候选靶基因进行实验验证,确认miR-486与所选4个候选靶基因存在靶向调控关系,说明本研究获得的miR-486的靶基因数据可靠性高。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1186/1471-2164-14-1URL [本文引用: 1]
DOI:10.1038/ng1810URL [本文引用: 1]
DOI:https://www.chinaagrisci.com/article/2021/0578-1752/10.1186/2044-5040-1-27URL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 5]
[本文引用: 5]
[P].
[本文引用: 1]
[P].
[本文引用: 1]
DOI:10.1093/nar/gkh021URL [本文引用: 1]
DOI:10.1093/nar/gkm882URL [本文引用: 1]
DOI:10.1051/rnd:2002035URL [本文引用: 1]
URL [本文引用: 2]
DOI:10.1242/dev.068353URL [本文引用: 1]
[本文引用: 2]
DOI:10.1016/0022-510X(87)90181-XURL [本文引用: 1]
DOI:10.1002/aja.1002030104URL [本文引用: 1]
DOI:10.2527/1992.7061806xURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1073/pnas.1000300107URL [本文引用: 3]
DOI:10.1080/09687860500244262URL [本文引用: 1]
DOI:10.1038/nrm3330URL [本文引用: 1]
DOI:10.1038/ncomms14328URL [本文引用: 1]
DOI:10.1371/journal.pone.0025655URL [本文引用: 1]
DOI:10.18632/oncotarget.v8i6URL [本文引用: 1]
DOI:10.1172/JCI73579URL [本文引用: 1]
DOI:10.1038/ki.2012.84URL [本文引用: 1]
DOI:10.1038/nrneph.2014.112URL [本文引用: 1]
DOI:10.1371/journal.pone.0196232URL [本文引用: 1]
DOI:10.1016/S1471-4914(03)00138-2URL [本文引用: 1]
DOI:10.1016/j.phrs.2015.05.010URL [本文引用: 1]
[本文引用: 1]
DOI:10.1016/S1097-2765(04)00211-4URL [本文引用: 1]
DOI:10.14814/phy2.13931URL [本文引用: 1]
DOI:10.3892/mmr.2015.4748URL [本文引用: 1]
DOI:10.1038/s41580-018-0045-7URL [本文引用: 1]
DOI:10.14348/molcells.2016.0013URL [本文引用: 1]
DOI:10.1093/nar/gkl743URL [本文引用: 1]
DOI:10.1038/nature09267URL [本文引用: 1]
DOI:10.1126/science.1215691URL [本文引用: 1]
DOI:10.1038/nrg3765URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}