Analysis of Key Genes About Flower Color Variation in Iris hollandica
LIN Bing,1, CHEN YiQuan2, ZHONG HuaiQin1, YE XiuXian1, FAN RongHui,11Institute of Crop Sciences, Fujian Academy of Agricultural Science, Fuzhou 350013 2Institute of Agriculture and Engineering Technology, Fujian Academy of Agricultural Science, Fuzhou 350013
Abstract 【Objective】 Flower color variation is of great significance for enriching color of ornamental plants, but it is difficult to clarify variation mechanism due to uncertainty of flower color variation. Dutch iris (Iris hollandica) is an important bulbous ornamental plant. In this study, the blue-purple wild type ‘Zhanchi’ and white mutant strain 'Yufei' of Dutch iris were investigated to explore molecular mechanism and difference of pigment accumulation, so as to provide a basis for mechanism of flower color variation.【Method】In this study, using inner tepals of ‘Zhanchi’ and ‘Yufei’ from Dutch Iris as materials, UHPLC-QTOF-MS method was used to determine types and contents of anthocyanins and flavonols from two varieties of Dutch Iris, and the different expression genes related to anthocyanin synthesis were screened by transcriptome sequencing. Using different flowers in developmental stages of two varieties as materials, the different expression genes were verified by qRT-PCR.【Result】Results of metabolomics analysis revealed that delphinidin, cyanidin and its derivatives were accumulated in blue-purple flowers, which were almost no accumulation in white flowers, while the flavonol contents were increased in white ‘Yufei’. Results of RNA-seq analysis revealed that a total of 46 485 unigenes were obtained, and 27 073 unigenes of them were functionally annotated by public databases, accounting for 41.85% of the total. And 701 differentially expressed genes were obtained, 485 genes of which were up-regulated and 216 genes were down-regulated in white ‘Yufei’. Two dihydroflavonol-4-reductase genes and one flavonol synthase gene involving in anthocyanin biosynthetic pathway had different expression, named IhDFR1, IhDFR2 and IhFLS1. Down-regulated expression of IhDFR1 and IhDFR2 as well as up-regulated expression of IhFLS1 in white ‘Yufei’ led to significant decrease of anthocyanins and accumulation of flavonols, which caused metabolic flow from anthocyanin to flavonol. The qRT-PCR results of three genes showed that expression levels of IhDFR1 and IhDFR2 increased in blue-purple flowers during flower developmental stage, but low expression in white flowers, and expression level of IhFLS1 increased in white flowers during flower developmental stage, but low expression in blue-purple flowers, which was consistent with RNA-seq results.【Conclusion】Low expression of IhDFR1 and IhDFR2 as well as high expression of IhFLS1 in white ‘Yufei’ blocked accumulation of anthocyanins, and some of metabolic flow changed from anthocyanins to flavonols, resulting in the change of flower color from blue violet to white. Keywords:Iris hollandica;anthocyanin;flavonol;transcriptome analyses;dihydroflavonol-4-reductase gene;flavonol synthase gene
PDF (1211KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 林兵, 陈艺荃, 钟淮钦, 叶秀仙, 樊荣辉. 荷兰鸢尾‘玉妃’花色变异关键结构基因分析[J]. 中国农业科学, 2021, 54(12): 2644-2652 doi:10.3864/j.issn.0578-1752.2021.12.014 LIN Bing, CHEN YiQuan, ZHONG HuaiQin, YE XiuXian, FAN RongHui. Analysis of Key Genes About Flower Color Variation in Iris hollandica[J]. Scientia Acricultura Sinica, 2021, 54(12): 2644-2652 doi:10.3864/j.issn.0578-1752.2021.12.014
LINB, ZHONG HQ, HUANG ML, FAN RH, LUO YH. The study of 60Co-γ ray irradiation effects on flower color of Iris hollandica Journal of Nuclear Agricultural Sciences, 2019,33(4):633-639. (in Chinese) [本文引用: 1]
ZHOU CB, MEIX, RORHENBERGD O N, YANGZ B, ZHANGW T, WANS H, YANGH J, ZHANGL Y. Metabolome and transcriptome analysis reveals putative genes involved in anthocyanin accumulation and coloration in white and pink tea (Camellia sinensis) flower Molecules, 2020,25:190. DOI:10.3390/molecules25010190URL [本文引用: 1]
LOUQ, LIU YL, QI YY, JIAO SZ, TIAN FF, JIANGL, WANG YJ. Transcriptome sequencing and metabolite analysis reveals the role of delphinidin metabolism in flower colour in grape hyacinth Journal of Experimental Botany, 2014,65(12):3157-3164. DOI:10.1093/jxb/eru168URL [本文引用: 2]
WUQ, WUJ, Li SS, ZHANG HJ, FENG CY, YIN DD, WU RY, WANG LS. Transcriptome sequencing and metabolite analysis for revealing the blue flower formation in waterlily BMC Genomics, 2016,17:897. DOI:10.1186/s12864-016-3226-9URL [本文引用: 1]
TANAKAY, BRUGLIERAF, KALCG, SENIORM, DYSONB, NAKAMURAN, KATSUMOTOY, CHANDLERS. Flower color modification by engineering of the flavonoid biosynthetic pathway: Practical perspectives Bioscience Biotechnology and Biochemistry, 2010,74(9):1760-1769. DOI:10.1271/bbb.100358URL [本文引用: 1]
TANAKAY, SASAKIN, OHMIYAA. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids The Plant Journal, 2008,54(4):733-749. DOI:10.1111/j.1365-313X.2008.03447.xURL [本文引用: 1]
NAKATSUKAT, MISHIBAK, KUBOTAA, ABEY, YAMAMURAS, NAKAMURAN, TANAKAY, NISHIHARAM. Genetic engineering of novel flower colour by suppression of anthocyanin modification genes in gentian Journal of Plant Physiology, 2010,167(3):231-237. DOI:10.1016/j.jplph.2009.08.007URL [本文引用: 1]
WU XX, GONG QH, NI XP, ZHOUY, GAO ZH. UFGT: the key enzyme associated with the petals variegation in Japanese Apricot Frontiers in Plant Science, 2017,8:108. [本文引用: 1]
MIZUNOT, UEHARAA, MIZUTAD, YABUYAT, IWASHINAT. Contribution of anthocyanin-flavone copigmentation to grayed violet flower color of Dutch iris cultivar ‘Tiger’s Eye’ under the presence of carotenoids Scientia Horticulturae, 2015,186(21):201-206. DOI:10.1016/j.scienta.2015.01.037URL [本文引用: 1]
YOSHIHARAN, IMAYAMAT, FUKUCHI-MIZUTANIM, OKUHARAH, TANAKAY, INOI, YABUYOT. cDNA cloning and characterization of UDP-glucose: Anthocyanidin 3-O-glucosyltransferase in Iris hollandica Plant Science, 2005,169(3):496-501. DOI:10.1016/j.plantsci.2005.04.007URL [本文引用: 1]
IMAYAMAT, YOSHIHARAN, FUKUCHIMIZUTANIM, TANAKAY, INOI, YABUYAT. Isolation and characterization of a cDNA clone of UDP-glucose: anthocyanin 5-O-glucosyltransferase in Iris hollandica Plant Science, 2004,167(6):1243-1248. DOI:10.1016/j.plantsci.2004.06.020URL [本文引用: 1]
YOSHIHARAN, FUKUCHI-MIZUTANIM, OKUHARAH, TANAKAY, YABUYAT. Molecular cloning and characterization of O-methyltransferases from the flower buds of Iris hollandica Journal of Plant Physiology, 2008,165(4):415-422. DOI:10.1016/j.jplph.2006.12.002URL [本文引用: 1]
TRAPNELLlC, WILLIAMS BA, PERTEAG, MORTAZAVIA, KWANG, BAREN MJ, SALZBERG SL, WOLD BJ, PACHTERL. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation Nature Biotechnology, 2010,28:511-515. DOI:10.1038/nbt.1621URL [本文引用: 1]
FAN RH, HUANG ML. Progress in regulation of anthocyanins Chinese Journal of Cell Biology, 2013,35(5):741-746. (in Chinese) [本文引用: 1]
ZHENGC, MA JQ, CHENJ D, MAC L, CHENW, YAOM Z, CHENL. Gene coexpression networks reveal key drivers of flavonoid variation in eleven tea cultivars (Camellia sinensis) Journal of Agricultural and Food Chemistry, 2019,67(35):9967-9978. DOI:10.1021/acs.jafc.9b04422URL [本文引用: 1]
CASTELLARIN SD, GASPERO GD. Transcriptional control of anthocyanin biosynthetic genes in extreme phenotypes for berry pigmentation of naturally occurring grapevines BMC Plant Biology, 2007,7:46. DOI:10.1186/1471-2229-7-46URL [本文引用: 1]
WANG KL, BOLITHOK, GRAFTONK, KORTSTEEA, KARUNAIRETNAMS, MCGHIE TK, ESPLEY RV, HELLENS RP, ALLAN AC. An R2R3 MYB transcription factor associated with regulation of the anthocyanin biosynthetic pathway in Rosaceae BMC Plant Biology, 2010,10:50. DOI:10.1186/1471-2229-10-50URL [本文引用: 1]
FENGC, CHENM, XU CJ, BAIL, YIN XR, LIX, ALLAN AC, FERGUSON IB, CHEN KS. Transcriptomic analysis of Chinese bayberry (Myrica rubra) fruit development and ripening using RNA-Seq BMC Genomics, 2012,13:19. DOI:10.1186/1471-2164-13-19URL [本文引用: 1]
YUANY, MA XH, SHI YM, TANG DQ. Isolation and expression analysis of six putative structural genes involved in anthocyanin biosynthesis in Tulipa fosteriana Scientia Horticulturae, 2013,153(4):93-102. DOI:10.1016/j.scienta.2013.02.008URL [本文引用: 1]
HUANGY, GOU JQ, JIA ZC, YANGL, SUN YM, XIAO XY, SONGF, LUO KM. Molecular cloning and characterization of two genes encoding dihydroflavonol-4-reductase from Populus trichocarpa PLoS ONE, 2012,7(2):e30364. DOI:10.1371/journal.pone.0030364URL [本文引用: 1]
LUOP, NING GG, WANGZ, SHEN YX, JIN HN, LI PH, HUANG SS, ZHAOJ, BAO MZ. Disequilibrium of flavonol synthase and dihydroflavonol-4-reductase expression associated tightly to white vs. red color flower formation in plants Frontiers in Plant Science, 2015,6:1257 [本文引用: 1]
HAN YP, VIMOLMANGKANGS, SORIA-GUERRAR E, KORBANS S. Introduction of apple ANR genes into tobacco inhibits expression of both CHI and DFR genes in flowers, leading to loss of anthocyanin Journal of Experimental Botany, 2012,63(7):2437-2447. DOI:10.1093/jxb/err415URL [本文引用: 1]
SAITOR, FUKUTAN, OHMIYAA, ITOHY, OZEKIY, KUCHITSUK, NAKAYAMAM. Regulation of anthocyanin biosynthesis involved in the formation of marginal picotee petals in Petunia Plant Science, 2006,170(4):828-834. DOI:10.1016/j.plantsci.2005.12.003URL [本文引用: 1]
SPITZERB, ZVIM M B, OVADISM, MARHEVKAE, BAEKAIO, EDELBAUMO, MARTONI, MASCIT, ALONM, MORINS, ROGACHEVI, AHARONIA, VAINSTEINA. Reverse genetics of floral scent: Application of tobacco rattle virus-based gene silencing in Petunia Plant Physiology, 2007,145(4):1241-1250. DOI:10.1104/pp.107.105916URL [本文引用: 1]
HEMLEBENV, DRESSELA, EPPINGB, LUKACINR, MARTENSS, AUSTINM. Characterization and structural features of a chalcone synthase mutation in a white-flowering line of Matthiola incana R. Br. (Brassicaceae) Plant Molecular Biology, 2004,55(3):455-465. DOI:10.1007/s11103-004-1125-yURL [本文引用: 1]
ZHANG YZ, CHENG YW, YA HY, XU SZ, HAN JM. Transcriptome sequencing of purple petal spot region in tree peony reveals differentially expressed anthocyanin structural genes Frontiers in Plant Science, 2015,6:964. [本文引用: 1]
CLARK ST, VERWOERD WS. A systems approach to identifying correlated gene targets for the loss of colour pigmentation in plants BMC Bioinformatics, 2011,12:343. DOI:10.1186/1471-2105-12-343URL [本文引用: 1]
MA HW, ZHAO XM, YUAN YJ, ZENG AP. Decomposition of metabolic network into functional modules based on the global connectivity structure of reaction graph Bioinformatics, 2004,20(12):1870-1876. DOI:10.1093/bioinformatics/bth167URL [本文引用: 1]
BOGSJ, JAFFE FW, TAKOS AM, WALKER AR, ROBINSON SP. The grapevine transcription factor VvMYBPA1 regulates proanthocyanidin synthesis during fruit development Plant Physiology, 2007,143:1347-1361. DOI:10.1104/pp.106.093203URL [本文引用: 1]
HAN YP, VIMOLMANGKANGS, SORIA-GUERRAR E, KORBANS S. Introduction of apple ANR genes into tobacco inhibits expression of both CHI and DFR genes in flowers, leading to loss of anthocyanin Journal of Experimental Botany, 2012,63(7):2437-2447. DOI:10.1093/jxb/err415URL [本文引用: 1]
 ,1, 陈艺荃2, 钟淮钦1, 叶秀仙1, 樊荣辉
,1, 陈艺荃2, 钟淮钦1, 叶秀仙1, 樊荣辉

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT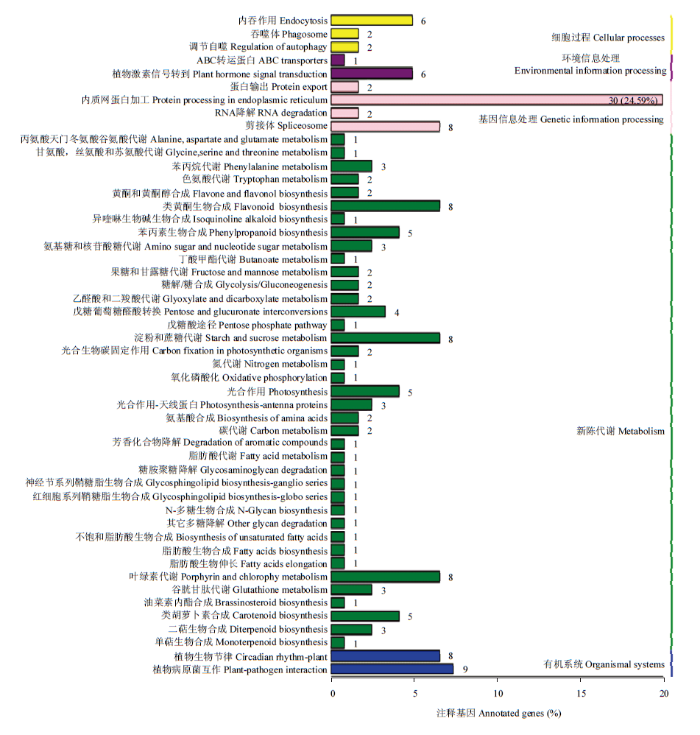 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT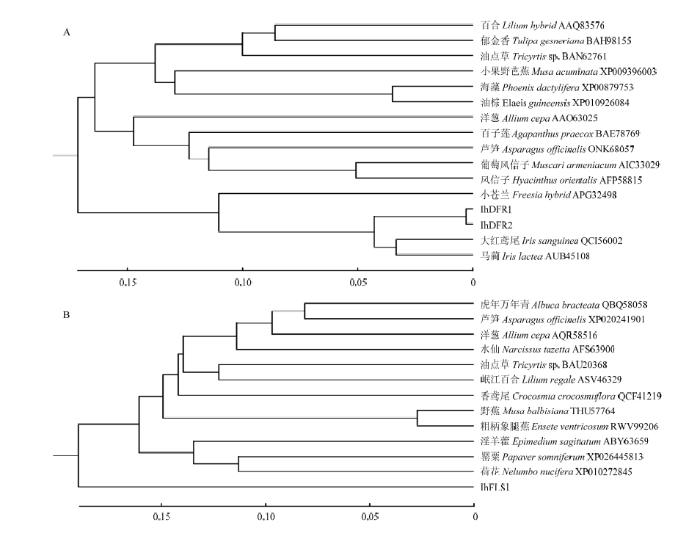 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}