 ,1, 骆强伟2, 李莎莎1, 王跃进,1
,1, 骆强伟2, 李莎莎1, 王跃进,1Optimization of Embryo Rescue Technique and Production of Potential Seedless Grape Germplasm with Rosy Aroma
JIA ShanShan,1, LUO QiangWei2, LI ShaSha1, WANG YueJin,1通讯作者:
责任编辑: 赵伶俐
收稿日期:2020-01-9接受日期:2020-06-5网络出版日期:2020-08-16
| 基金资助: |
Received:2020-01-9Accepted:2020-06-5Online:2020-08-16
作者简介 About authors
贾姗姗,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (2798KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
贾姗姗, 骆强伟, 李莎莎, 王跃进. 葡萄胚挽救技术优化及无核和玫瑰香味新种质创制[J]. 中国农业科学, 2020, 53(16): 3344-3355 doi:10.3864/j.issn.0578-1752.2020.16.012
JIA ShanShan, LUO QiangWei, LI ShaSha, WANG YueJin.
0 引言
【研究意义】无核作为果树育种的重要性状之一,因其在不改变果实大小的情况下可减少种子量,增加水果可食率,进而提高水果对食品加工业的适用性;在增强对消费者吸引力的同时,无核果实又是人类消费文明进程中的重要需求。葡萄果实的芳香气味不但能增加鲜食葡萄的风味和口感,其中含有的挥发性芳香物质可以通过味觉及嗅觉被人感知从而产生愉悦感,调节人体心理与生理机能平衡,与人类营养健康密切相关。目前,无核和香味形成的机理研究较少,无核和香味性状结合的果实育种更少,因此,结合无核及香味两种性状的新品种研发十分迫切。随着现代研究手段与分析技术不断完善,带有香味的无核葡萄逐渐成为国内外葡萄育种工作者的主要育种目标。目前利用常规杂交可以实现无核和香味两种优良性状的聚合从而育出无核香味新品种,而大田杂交结合胚挽救技术可在聚合两种优良性状的基础上,扩大亲本选择范围、缩短育种年限,成为育种新手段[1,2,3,4,5]。影响胚挽救效率的主要因素有亲本基因型、取样时间、培养基添加物、培养时间、培养方式等[3,6-9]。利用分子标记技术,通过检测后代DNA序列差异直接选择合适的基因型,加快筛选速度,改变了传统的田间杂交种表型鉴定法,提高了育种效率。【前人研究进展】自20世纪90年代固相微萃取技术(SPME)提出[10]后,随着质谱分析的发展,近年来固相微萃取结合气相质谱联用技术(HS-SPME-GC-MS)开始在果实香气物质测定领域广泛应用[11,12]。目前已有研究证实葡萄中芳香物质的形成存在3种途径:脂肪酸代谢途径、氨基酸代谢途径和萜烯类化合物代谢途径[13]。随着控制果实中萜烯类合成酶及脂肪氧合酶的陆续发现,其关键基因也开始被报道。其中,VvDXS是萜类合成的关键酶[14]。萜烯合酶(TPS)为控制萜烯类合成最终产物的酶,在欧洲葡萄中被大量检测到[15]。研究认为萜烯类物质是葡萄玫瑰香味的主要呈香物质,其形成的香气,是果实及其加工的食品中与营养和健康最为密切相关的物质[16,17,18]。目前大部分无核香味新品种是利用常规杂交培育的,后代聚集无核、香味等优良性状概率较低,且对父本品种要求较高,自20世纪70年代开始,国内外育出的香味品种共40余种[19],而利用胚挽救技术培育出的无核香味新品种较少。笔者课题组前期选择带有玫瑰香味的葡萄品种,通过配置无核×玫瑰香味杂交组合,并利用胚挽救育种技术,获得大批带有玫瑰香味的无核葡萄新种质,确定了胚挽救过程中不同母本的杂交组合适宜取样时间[9,20]。【本研究切入点】近年来无核香味葡萄新品种的育种进程比较缓慢。本研究从胚挽救影响的因素出发,筛选玫瑰香味性状优良亲本配置无核品种×玫瑰香味品种的杂交组合;优化胚萌发培养基,对杂种后代进行早期无核性状的辅助筛选。【拟解决的关键问题】测定香味品种果实中挥发性化合物含量以揭示葡萄果实中决定玫瑰香味的关键物质,通过对香味物质的定量分析为培育优质香味葡萄提供重要参考;同时将葡萄香味性状融入无核品种,通过分子标记初步筛选携带无核标记的杂交后代,进而获得携带香味基因的无核新种质。通过比较亲本基因型对胚挽救结果的影响和对胚萌发培养基的筛选与优化,提高无核香味葡萄胚挽救效率。1 材料与方法
试验于2018年5月至2019年5月分别在新疆维吾尔自治区瓜果研究所、旱区作物逆境生物学国家重点实验室和西北农林科技大学资源圃进行。1.1 材料
用于香气物质测定的品种为:‘玫瑰香’(V. vinifera L. cv. Muscat Hamburg)、‘阳光玫瑰’(V. labruscana Bailey×V. vinifera L. cv. Shine Muscat)、‘早玫瑰’(V. vinifera L. cv. Zaomeiguixiang)、‘巨玫瑰’(V. vinifera L.×V. labrusca L. cv. Jumeigui)、‘亚历山大’(V. vinifera L. cv. Muscat of Alexandria)、‘沙巴珍珠’(V. vinifera L. cv. Pearl of Csaba)、‘意大利’(V. vinifera L. cv. Italia)、‘粉红玫瑰’(V. vinifera L. cv. Muscat Rose)、‘无核白’(V. vinifera L. cv. Thompson Seedless),其中‘亚历山大’取于北京市林业果树科学研究院试验场,其他葡萄品种取于西北农林科技大学资源圃。用于胚挽救杂交的供试材料中,母本为5个种子败育型无核品种,具体为:‘爱神玫瑰’‘火焰无核’‘红宝石无核’‘昆香无核’‘克瑞森无核’,父本为6个玫瑰香味品种‘玫瑰香’‘阳光玫瑰’‘爱神玫瑰’‘红亚历山大’‘昆香无核’‘新郁’,以上杂交试验于新疆维吾尔自治区瓜果研究所进行,供试葡萄品种均采用水平棚架栽培,株距1 m,行距5 m,东西行向,葡萄园地理位置北纬42.91°、东经90.30°。杂交组合的配置见表1。Table 1
表1
表1杂交组合配置
Table 1
| 杂交组合(母本×父本) Cross combination (♀×♂) | 母本Female parent | 父本Male parent | ||
|---|---|---|---|---|
| 种或杂种 Species or hybrid | 性状 Characteristic | 种或杂种 Species or hybrid | 性状 Characteristic | |
| 爱神玫瑰×玫瑰香 Aishenmeigui × Muscat Hamburg | 欧亚种 V. vinifera | 无核、玫瑰香味 Stenospermic, Muscat-scented | 欧亚种 V. vinifera | 有核、玫瑰香味 Seeded, Muscat-scented |
| 爱神玫瑰×阳光玫瑰 Aishenmeigui × Shine-Muscat | 欧亚种 V. vinifera | 无核、玫瑰香味 Stenospermic, Muscat-scented | 欧美杂种 V. labruscana ×V. vinifera | 有核、玫瑰香味 Seeded, Muscat-scented |
| 爱神玫瑰×昆香无核 Aishenmeigui × Kunxiang Seedless | 欧亚种 V. vinifera | 无核、玫瑰香味 Stenospermic, Muscat-scented | 欧亚种 V. vinifera | 无核、玫瑰香味 Stenospermic, Muscat-scented |
| 火焰无核×红亚历山大 Flame Seedless × Red Alexander | 欧亚种 V. vinifera | 无核 Stenospermic | 欧亚种 V. vinifera | 有核、玫瑰香味 Seeded, Muscat-scented |
| 火焰无核×阳光玫瑰 Flame Seedless × Shine-Muscat | 欧亚种 V. vinifera | 无核 Stenospermic | 欧美杂种 V. labruscana ×V. vinifera | 有核、玫瑰香味 Seeded, Muscat-scented |
| 火焰无核×新郁 Flame Seedless × Xinyu | 欧亚种 V. vinifera | 无核 Stenospermic | 欧亚种 V. vinifera | 有核、玫瑰香味 Seeded, Muscat-scented |
| 昆香无核×爱神玫瑰 Kunxiang Seedless × Aishenmeigui | 欧亚种 V. vinifera | 无核、玫瑰香味 Stenospermic, Muscat-scented | 欧亚种 V. vinifera | 无核、玫瑰香味 Stenospermic, Muscat-scented |
| 昆香无核×玫瑰香 Kunxiang Seedless × Muscat Hamburg | 欧亚种 V. vinifera | 无核、玫瑰香味 Stenospermic, Muscat-scented | 欧亚种 V. vinifera | 有核、玫瑰香味 Seeded, Muscat-scented |
| 克瑞森无核×玫瑰香 Crimson Seedless × Muscat Hamburg | 欧亚种 V. vinifera | 无核 Stenospermic | 欧亚种 V. vinifera | 有核、玫瑰香味 Seeded, Muscat-scented |
| 克瑞森无核×新郁 Crimson Seedless × Xinyu | 欧亚种 V. vinifera | 无核 Stenospermic | 欧亚种 V. vinifera | 有核、玫瑰香味 Seeded, Muscat-scented |
| 红宝石无核×玫瑰香 Ruby Seedless × Muscat Hamburg | 欧亚种 V. vinifera | 无核 Stenospermic | 欧亚种 V. vinifera | 有核、玫瑰香味 Seeded, Muscat-scented |
| 红宝石无核×红亚历山大 Ruby Seedless × Red Alexander | 欧亚种 V. vinifera | 无核 Stenospermic | 欧亚种 V. vinifera | 有核、玫瑰香味 Seeded, Muscat-scented |
| 红宝石无核×爱神玫瑰 Ruby Seedless × Aishenmeigui | 欧亚种 V. vinifera | 无核 Stenospermic | 欧亚种 V. vinifera | 无核、玫瑰香味 Stenospermic, Muscat-scented |
新窗口打开|下载CSV
Table 2
表2
表2玫瑰香味葡萄果实中所含化合物种类及含量
Table 2
| 化合物 Compound | 沙巴珍珠 Pearl of Csaba | 早玫瑰 Zao Meiguixiang | 玫瑰香 Muscat Hamburg | 亚历山大 Muscat of Alexandria | 巨玫瑰 Jumeigui | 金星无核 Venus Seedless | 意大利 Italia | 粉红玫瑰 Muscat Rose | 阳光玫瑰 Shine- Muscat | 无核白 Thompson Seedless | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 醛类Aldehydes | 总醛 Total aldehydes | 8.9300 | 4.4924 | 9.2314 | 4.9050 | 15.8080 | 15.7184 | 8.0026 | 10.1754 | 2.1530 | 15.5756 |
| 醇类 Alcohols | 总醇 Total alcohols | 2.0900 | 0.0842 | 0.2296 | 1.4376 | 1.4816 | 0.3200 | 0.1754 | 0.5486 | 0.5210 | 0.1138 |
| 烯类 Olefins | 总烯 Total olefins | 0.3522 | 0.0504 | 0.0612 | 0.2074 | 0.1184 | 0.1266 | 0 | 0.0872 | 0.0816 | 0 |
| 烷类 Alkenes | 总烷 Total alkenes | 0.0362 | 0.0328 | 0.0098 | 0.0112 | 1.8162 | 0.2784 | 0.0154 | 0.0314 | 0.0588 | 0.3464 |
| 酮类 Ketones | 总酮 Total ketones | 0 | 0 | 0.0848 | 0.0076 | 0 | 0.0234 | 0.0382 | 0.0122 | 0 | 0 |
| 酯类 Esters | 总酯 Total esters | 0 | 0 | 0 | 0.1202 | 0.6918 | 0.0084 | 0 | 0.0122 | 0.0094 | 0 |
| 酸类 Acids | 总酸 Total acids | 0 | 0 | 0 | 0.0328 | 0 | 0.0116 | 0.0272 | 0 | 0 | 0 |
| 萜烯类 Terpenes | 香茅醇 Citronellol | 0.2912 | 0.0062 | 0 | 0 | 0.3490 | 0 | 0 | 0 | 0 | 0 |
| 香叶醇 Geraniol | 0.4432 | 0.0152 | 0.0088 | 0.1524 | 0.6448 | 0 | 0.0246 | 0.0446 | 0 | 0 | |
| 香叶醛 Citral | 0.0608 | 0.0024 | 0.0286 | 0.0244 | 0 | 0 | 0 | 0 | 0 | ||
| 里那醇 Linalool | 0.5532 | 0.0396 | 0.1656 | 1.0790 | 0.0980 | 0 | 0 | 0.1184 | 0.4812 | ||
| 松油醇 Terpineol | 0.0340 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| à-松油醇 à-Terpineolα- | 0 | 0 | 0 | 0.0102 | 0 | 0 | 0.0156 | 0 | 0 | ||
| 脱氢芳樟醇 1,5,7-Octatrien-3-ol, 3,7- dimethyl | 0 | 0 | 0 | 0.0035 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 总萜烯Total terpenes | 1.3824 | 0.0634 | 0.1744 | 1.2636 | 1.1264 | 0 | 0.0246 | 0.1788 | 0.4812 | 0 | |
| 其他 Others | 0.0234 | 0.0274 | 0.0374 | 0.0454 | 0.2674 | 0.0516 | 0.0554 | 0.1038 | 0.0076 | 0.0194 | |
| 总挥发性物质含量 Total volatile matter content | 12.8142 | 4.7506 | 9.8286 | 8.0308 | 21.3098 | 16.5384 | 8.3388 | 11.1496 | 3.3126 | 16.0552 | |
新窗口打开|下载CSV
1.2 方法
1.2.1 果实香味物质测定 选择10个葡萄品种的成熟果实,利用气质联用仪(HS-SPME-GC-MS)测定葡萄果实香气物质含量,所用色谱柱为HP- INNOWAX(60 m×0.25 mm×0.25 μm)毛细管柱,内标物为10 μL 2-辛醇(0.1 g?L-1),其余具体操作参照谭伟等[21]的方法。1.2.2 胚挽救流程 选择长势良好的花序,盛花前3—4 d人工去雄。去雄3 d后,柱头分泌黏液时,进行人工授粉。授粉后约45 d时取杂交组合果穗,将果实摘下放入网袋中流水冲洗6—8 h。在无菌条件下,将果实内胚珠取出,接种于胚发育MM3固体培养基中暗培养。10周后,在解剖镜下取胚接种于胚萌发培养基中进行光照培养,培养1个月后统计萌发胚数,2个月后统计成苗数。胚挽救具体程序参照文献[7,22-23]的方法进行。光照过程中及时观察胚的生长状态,统计不同组合发育胚的个数和正常成苗数,计算胚发育率和成苗率,胚发育率(%)=发育胚数/接种胚珠数×100,胚萌发率(或成苗率)(%)=胚萌发数(或成苗数)/接种胚数×100。
1.2.3 胚萌发培养基最适生长调节剂浓度筛选 试验共设置了9种不同的胚萌发培养基,对2,4-二氯苯氧乙酸(2,4-D)、激动素(KT)、玉米素(ZT)及萘乙酸(NAA)4种生长调节剂的不同浓度配比,以WPM培养基为基础培养基,进行胚萌发培养基的优化(表3)。比较不同培养基中胚的萌发率和成苗率,从而筛选出胚萌发培养基中最适生长调节剂浓度及成分。取3次生物学重复平均值,统计方法同上。
Table 3
表3
表3无核香味葡萄胚挽救育种结果
Table 3
| 杂交组合 Cross | 果粒数 Number of berries | 胚珠数 Number of cultured ovule | 发育胚Developed embryos | 成苗Plantlets formation | ||
|---|---|---|---|---|---|---|
| 数量 Number | 占比 Rate (%) | 数量 Number | 占比 Rate (%) | |||
| 爱神玫瑰×玫瑰香 Aishenmeigui × Muscat Hamburg | 285 | 508 | 75 | 15.69 | 14 | 18.67 |
| 爱神玫瑰×阳光玫瑰 Aishenmeigui × Shine-Muscat | 1504 | 2086 | 335 | 16.06 | 79 | 23.58 |
| 爱神玫瑰×昆香无核 Aishenmeigui × Kunxiang Seedless | 620 | 750 | 71 | 9.47 | 17 | 23.49 |
| 火焰无核×红亚历山大 Flame Seedless × Red Alexander | 925 | 1208 | 133 | 11.01 | 50 | 37.59 |
| 火焰无核×阳光玫瑰 Flame Seedless × Shine-Muscat | 638 | 922 | 98 | 10.80 | 20 | 20.41 |
| 火焰无核×新郁 Flame Seedless × Xinyu | 665 | 635 | 34 | 5.48 | 8 | 23.53 |
| 昆香无核×爱神玫瑰 Kunxiang Seedless × Aishenmeigui | 2050 | 2184 | 305 | 13.97 | 70 | 22.95 |
| 昆香无核×玫瑰香 Kunxiang Seedless × Muscat Hamburg | 385 | 405 | 102 | 25.19 | 0 | 0 |
| 克瑞森无核×玫瑰香 Crimson Seedless × Muscat Hamburg | 435 | 441 | 19 | 4.31 | 1 | 5.26 |
| 克瑞森无核×新郁 Crimson Seedless × Xinyu | 635 | 540 | 16 | 5.61 | 1 | 6.25 |
| 红宝石无核×玫瑰香 Ruby Seedless × Muscat Hamburg | 1160 | 1895 | 395 | 20.84 | 186 | 47.09 |
| 红宝石无核×红亚历山大 Ruby Seedless × Red Alexander | 1308 | 2956 | 763 | 25.81 | 249 | 32.63 |
| 红宝石无核×爱神玫瑰 Ruby Seedless × Aishenmeigui | 1243 | 2486 | 1139 | 48.59 | 589 | 51.71 |
| 合计 Total | 11853 | 17016 | 3485 | 20.48 | 1284 | 36.84 |
新窗口打开|下载CSV
Table 4
表4
表4不同生长调节剂对胚挽救成苗的影响
Table 4
| 杂交组合 Cross | 处理组 Treatment | 生长调节剂成分 Exogenous hormone (mg·L-1) | 胚珠数 Number of cultured ovules | 发育胚 Developed embryos | 萌发 Germination | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2,4-D | KT | ZT | NAA | 数量Number | 占比 Rate (%) | 数量Number | 占比 Rate (%) | |||
| 昆香无核×爱神玫瑰 Kunxiang Seedless × Aishenmeigui | T1 | 0 | 0 | 0 | 0 | 135 | 22 | 16.30b | 10 | 7.41a |
| T2 | 0 | 1 | 1 | 0.5 | 150 | 33 | 22.00a | 17 | 11.33a | |
| T3 | 0 | 2 | 2 | 1 | 150 | 26 | 17.33b | 14 | 9.33a | |
| T4 | 1 | 0 | 1 | 0.5 | 150 | 7 | 4.67bc | 1 | 0.67b | |
| T5 | 1 | 1 | 2 | 0 | 132 | 7 | 5.30b | 2 | 1.52b | |
| T6 | 1 | 2 | 0 | 0.5 | 140 | 1 | 0.72c | 1 | 0.72b | |
| T7 | 2 | 0 | 2 | 0.5 | 150 | 10 | 6.67b | 2 | 1.33b | |
| T8 | 2 | 1 | 0 | 1 | 150 | 4 | 2.67c | 1 | 0.67b | |
| T9 | 2 | 2 | 1 | 0 | 150 | 6 | 4.00bc | 1 | 0.67b | |
新窗口打开|下载CSV
1.2.4 胚发育形态差异观察统计 以杂交组合‘红宝石无核’ב玫瑰香’和‘昆香无核’ב爱神玫瑰’为材料,统计不同发育阶段胚的数量,具体统计类型包括:心形胚(H)、球形胚(G)、鱼雷形胚(F)、子叶形胚(C)、畸形胚(A)5种。后期统计对比不同类型胚的成苗株数,并比较分析不同发育形态的胚接种后胚萌发率及成苗率的差异。
1.2.5 杂交后代无核标记检测 提取杂交亲本及后代基因组DNA,使用无核相关分子标记GLSP1-569、SCF27-2000及SCC8-1018标记进行检测,引物序列及反应程序参考文献[22,23]。SCC8-1018酶切具体操作如下:模板DNA经PCR扩增后,取20 μL PCR产物,加1 U Bgl Ⅱ酶,37℃下培养2—3 h,酶切产物用浓度为1.5%的琼脂糖凝胶电泳检测,两端电压为120 V,电泳结束后立即使用凝胶成像仪对其成像。
1.3 数据统计分析
香气物质经GC/MS分析,各色谱峰经计算机检索NIST 2011标准质谱图库,辅以人工分析对应化合物以定性鉴定;利用标准品法定量分析果实中提取出的香气物质,取3次生物学重复平均值作为香气物质的相对含量。胚发育率、萌发率和成苗率及其差异显著性分析利用Excel和SPSS软件进行。2 结果
2.1 果实风味物质检测与玫瑰香味品种筛选
对1个非香味品种及9个香味品种果实进行香气物质鉴定(表1)。由结果可看出,果实挥发性物质中的主要成分醛类、醇类、酯类等C6化合物含量较高。与对照相比,8个玫瑰香味葡萄品种果实香气成分中萜烯物质含量均较高,范围为0.0246—1.3824,其中含量最高的品种为‘莎巴珍珠’,而萜烯物质被认为是主导玫瑰香味的挥发性物质,基于此,本研究筛选出适于作为香味杂交父本的品种有‘莎巴珍珠’‘巨玫瑰’‘亚历山大’‘阳光玫瑰’。2.2 亲本基因型对胚挽救效果的影响
2.2.1 不同母本基因型对胚挽救效果的影响 本试验配置了13个杂交组合,共获得杂种胚珠17 016个,杂种苗1 284个。胚发育率的变化范围为4.31%—48.59%,成苗率的变化范围0—51.71%。分析母本基因型对胚挽救效率的影响,结果表明,以‘红宝石无核’‘火焰无核’为母本的杂交组合成苗率高于其他杂交组合,成苗率最高分别达到51.71%和37.59%(表3)。
2.2.2 不同父本基因型对胚挽救效果的影响 以‘爱神玫瑰’为母本,比较父本基因型影响,以‘昆香无核’作父本时成苗率最高(23.49%);以‘火焰无核’为母本与不同父本杂交,以‘红亚历山大’做父本时成苗率最高;以‘红宝石无核’为母本时,比较不同父本杂交后代成苗率,最高的为‘爱神玫瑰’。
在母本相同的情况下,以‘阳光玫瑰’‘红亚历山大’和‘爱神玫瑰’为父本的杂交组合成苗率较高,其中‘红亚历山大’成苗率最高。同时,试验中设置了‘昆香无核’和‘爱神玫瑰’正反交的杂交组合,结果表明,以‘昆香无核’为母本,‘爱神玫瑰’为父本的杂交组合胚发育率高于其反交组合,而两个组合的最终成苗率无明显差异(表3)。
2.3 生长调节剂对胚挽救结果的影响
T2处理组对胚萌发的促进效果最显著,因此,该组合中生长调节剂的最适浓度为:1.0 mg?L-1 KT+0.5 mg?L-1 NAA+1.0 mg?L-1 ZT。随着2,4-D浓度的升高,胚发育率及成苗率均下降;NAA与2,4-D同为生长素类似物,其作用机理类似,因此重复添加使培养基中生长素浓度过高则会抑制胚的生长;在一定范围内提升KT和ZT浓度对胚的发育有促进作用,二者在浓度约为1 mg?L-1时胚发育及萌发率较高。同时,T2处理的胚生根显著多于对照(图1),说明ZT对促进幼苗生根有明显的效果。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1‘昆香无核’ב爱神玫瑰’杂种胚生长发育情况
A—C:接种于对照培养基上的‘昆香无核’ב爱神玫瑰’的杂种胚生长发育情况,D—F:接种于T2处理培养基上的‘昆香无核’ב爱神玫瑰’的杂种胚生长发育情况
Fig. 1Growth and development of hybrid embryos of Kunxiang Seedless × Aishenmeigui
A-C: The growth and development of hybrid embryos of Kunxiang Seedless × Aishenmeigui inoculated on control medium, D-F: The growth and development of hybrid embryos of Kunxiang Seedless × Aishenmeigui inoculated on treatment 2 medium
2.4 不同形态杂种胚对胚挽救结果的影响
统计杂交组合‘昆香无核’ב爱神玫瑰’和‘红宝石无核’ב玫瑰香’分别暗培养7周和8周后发育形成不同形态的胚数量及所占比率,具体统计类型包括:球形胚(G)、心形胚(H)、鱼雷形胚(F)、子叶形胚(C)、畸形胚(A)。根据表5可以看出,‘昆香无核’ב爱神玫瑰’暗培养7周后获得幼胚中球形胚数量最多,而暗培养8周的‘红宝石无核’ב玫瑰香’中获得鱼雷形胚数量在总幼胚数中比率较高。对比不同类型胚的成苗株数,结果显示,在胚发育进程中,接种发育成熟的鱼雷形胚和子叶形胚可以获得较高的成苗率,其中子叶形胚的萌发率及成苗率都显著高于其他形态的杂种胚,均可达100%,而畸形胚接种后成苗率较低。Table 5
表5
表5不同形态杂种胚的萌发成苗
Table 5
| 杂交组合 Cross | 胚类型 Embryo code | 接种胚数Cultured embryos | 发育胚Developed embryos | 成苗Plantlets formation | |||
|---|---|---|---|---|---|---|---|
| 数量 Number | 占比 Rate (%) | 数量 Number | 占比 Rate (%) | 数量Number | 占比 Rate (%) | ||
| 昆香无核×爱神玫瑰 Kunxiang Seedless × Aishenmeigui | G | 55 | 47.41 | 23 | 41.82 | 12 | 21.82 |
| H | 26 | 22.41 | 22 | 84.62 | 6 | 23.08 | |
| F | 29 | 25.00 | 26 | 89.66 | 14 | 48.28 | |
| C | 2 | 1.72 | 2 | 100.00 | 2 | 100.00 | |
| A | 4 | 3.45 | 4 | 100.00 | 1 | 25.00 | |
| 红宝石无核×玫瑰香 Ruby Seedless × Muscat Hamburg | G | 77 | 39.49 | 32 | 41.56 | 17 | 22.08 |
| H | 5 | 2.56 | 2 | 40.00 | 1 | 20.00 | |
| F | 106 | 54.36 | 96 | 90.57 | 62 | 58.49 | |
| C | 4 | 2.05 | 3 | 75.00 | 3 | 75.00 | |
| A | 3 | 1.54 | 1 | 33.33 | 0 | 0 | |
新窗口打开|下载CSV
2.5 胚挽救杂交子代无核性状的早期分子标记筛选
2.5.1 不同组合亲本的无核性状分子标记筛选 利用3种无核标记对杂交组合中7个双亲进行无核性状筛选,无核探针GSLP1-569的扩增结果为:母本中‘火焰无核’‘红宝石无核’和‘克瑞森无核’扩增出了569 bp的特异性条带;父本中,‘爱神玫瑰’‘玫瑰香’未在569 bp处出现特异性条带(图2-A),可用于检测‘克瑞森无核’ב玫瑰香’及‘红宝石无核’ב玫瑰香’杂交后代无核性状。无核标记SCF27-2000的结果表明,亲本中‘玫瑰香’未扩增出2 000 bp的特异性条带(图2-B),可用于以‘爱神玫瑰’‘玫瑰香’为父本的所有组合杂交后代的无核早期鉴定。SCC8酶切后结果为‘火焰无核’和‘昆香无核’只有1 000 bp附近一条带,表现无核纯合基因型SCC8+/SCC8+,有核品种‘玫瑰香’只有750 bp附近一条带,表现出有核纯合基因型SCC8-/SCC8-(图2-C);而无核品种‘红宝石无核’及‘爱神玫瑰’同时扩增出1 000 bp及750 bp的两条带,表现为无核杂合基因型SCC8+/SCC8-,因此可用于‘红宝石无核’ב玫瑰香’‘爱神玫瑰’ב玫瑰香’‘昆香无核’ב爱神玫瑰’杂交后代无核性状的早期鉴定。图2
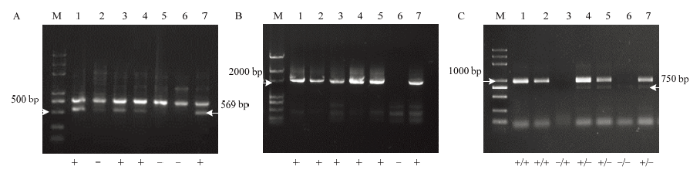 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2无核性状相关分子标记对杂交亲本的检测
A:无核探针GSLP1-569对杂交亲本的检测,B:无核分子标记SCF27-2000对杂交亲本的检测,C:无核标记SCC8酶切后对杂交亲本的检测。M:2k plus DNA Marker,1:火焰无核,2:昆香无核,3:克瑞森无核,4:红宝石无核,5:爱神玫瑰,6:玫瑰香,7:阳光玫瑰;“+”表示特异条带出现,“-”表示特异条带不出现。下同
Fig. 2Molecular marker-assisted selection of parents with grapevine seedlessness gene
A: The detection of parents with seedlessness gene using GSLP1-569 probe; B: Schematic the detection of parents with grapevine seedlessness gene using marker SCF27-2000; C: Schematic the detection of parents with grapevine seedlessness gene using marker SCC8. M: 2K plus Marker; 1: Flame Seedless; 2: Kunxiang Seedless; 3: Crimson Seedless; 4: Ruby Seedless; 5: Aishenmeigui; 6: Muscat Hamburg; 7: Shine-Muscat. “+” means the presence of specific bands, “-” means the absence of specific bands. The same as below
2.5.2 无核标记GSLP1-569、SCF27-2000及SCC8对‘红宝石无核’ב玫瑰香’杂交子代的检测 对‘红宝石无核’ב玫瑰香’41个子代的检测结果中,使用无核探针GSLP1-569扩增出569 bp特异性条带的共32个株系(图3-A),使用无核探针SCF27-2000扩增出2 000 bp特异性条带的共5个株系(图3-B),使用SCC8酶切后扩增出1 018 bp特异性条带的有23个株系,且均为纯合(图3-C),同时扩增出3种无核特异性条带的有4个株系(图3-A、3-B、3-C),编号为8-16、8-25、8-28、8-32。初步推断这4个株系为无核的概率最大,后代无核率为12.20%。
图3
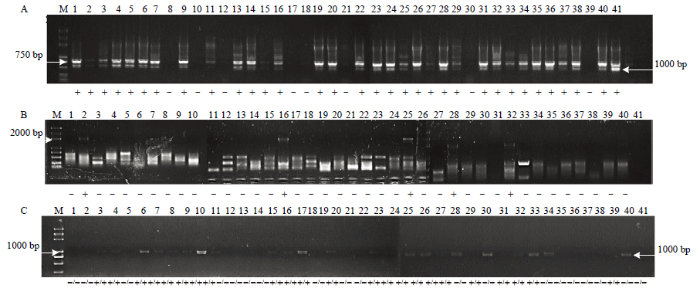 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同分子标记对‘红宝石无核’ב玫瑰香’杂交后代的检测
A:无核探针GSLP1-569的检测结果,B:无核标记SCF27-2000的检测结果,C:无核标记SCC8的检测结果;1—41:‘红宝石无核’ב玫瑰香’杂交子代
Fig. 3Molecular marker-assisted selection of hybrid progeny of Ruby Seedless × Muscat Hamburg cross with seedlessness gene
A: The probe of GSLP1-569 linked to seedlessness gene; B: The molecular marker of SCF27-2000 linked to seedlessness gene; C: The molecular marker of SCC8-1018 linked to seedlessness gene; 1-41: hybrid progeny seedlings of the cross Ruby Seedless × Muscat Hamburg
2.5.3 无核标记SCF27-2000、SCC8对‘爱神玫瑰’ב玫瑰香’杂交子代的检测 对‘爱神玫瑰’ב玫瑰香’14个杂种子代检测结果中,使用SCF27-2000标记扩增出2 000 bp特异性条带的有5个株系(图4-A),使用SCC8酶切后扩增出2 000 bp特异性条带的有9个株系(图4-B)。在两种标记检测时均扩增出了无核特异性条带的株系有3株(图4-A、4-B),编号为2-1、2-7、2-13,可认为这3个株系的无核概率更大,该组合杂交后代无核率为21.43%。
图4
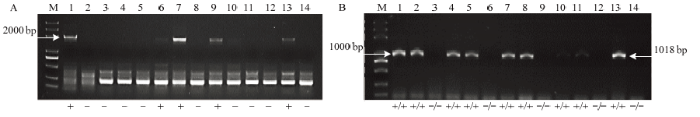 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同分子标记对‘爱神玫瑰’ב玫瑰香’杂交后代的检测
A:无核标记SCF27-2000的检测结果,B:无核标记SCC8-1018的检测结果;1—14:‘爱神玫瑰’ב玫瑰香’杂交子代
Fig. 4Molecular marker-assisted selection of hybrid progeny seedlings of the cross Aishenmeigui × Muscat Hamburg with grapevine seedlessness gene
A: The molecular marker of SCF27-2000 linked to seedlessness gene; B: The molecular marker of SCC8-1018 linked to seedlessness gene; 1-14: Hybrid progeny seedlings of the cross Aishenmeigui × Muscat Hamburg
2.5.4 无核标记SCF27-2000、SCC8酶切对‘昆香无核’ב爱神玫瑰’杂交子代的检测 对‘昆香无核’ב爱神玫瑰’46个杂种子代检测结果。其中使用SCF27-2000扩增出2 000 bp特异性条带的共20个株系(图5-A),经SCC8酶切后扩增出1 018 bp特异性条带的共31个株系(图5-B)。两种标记均可检测到无核性状的共20个株系(图5-A、5-B),编号为:1-12、1-14、1-16、1-17、1-18、1-20、1-23、1-24、1-25、1-27、1-28、1-29、1-30、1-31、1-32、1-37、1-39、1-42、1-43、1-44以上株系更可能被鉴定为无核品种,该杂交组合无核率为43.48%。
图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5不同分子标记对‘昆香无核’ב爱神玫瑰’杂交后代的检测
A:无核标记SCF27-2000的检测结果,B:无核标记SCC8酶切的检测结果,泳道1:‘昆香无核’,泳道2:‘爱神玫瑰’,泳道3-48:‘昆香无核’ב爱神玫瑰’杂交子代
Fig. 5Molecular marker-assisted selection of hybrid progeny seedlings of the cross ‘Kunxiang Seedless’× ‘Aishenmeigui’ with grapevine seedlessness gene
A: The molecular marker of SCF27-2000 linked to seedlessness gene; B: The molecular marker of SCC8-1018 linked to seedlessness gene; 1: Kunxiang Seedless; 2: Aishenmeigui; 3-48: Hybrid progeny seedlings of the cross Kunxiang Seedless × Aishenmeigui
3 讨论
已有研究认为欧亚种葡萄特有的玫瑰香味与其含有的萜类物质及其含量密切相关,玫瑰香味的主要呈香物质为萜类物质中的香叶醇、里那醇等,其中里那醇对玫瑰香味的贡献最大[11,24-26]。本研究中里那醇在8个玫瑰香味品种中含量范围为0.0396—1.0790,含量最高的品种为‘亚历山大’,含量最低的品种为‘早玫瑰’,这与感官感觉到的玫瑰香味浓淡相符[18]。本试验筛选出部分香味性状优良的玫瑰香味品种,用于创制无核香味新品种。下一步研究将以目前已测定的香味物质为参考依据,按照已报道的35个香味基因序列为基础,设计香味的基因标记,对本研究获得杂种后代的香味基因进行分子标记辅助选择,提高基因标记选择的效率。本研究发现影响胚挽救效果的因素有杂交组合亲本基因型、培养基成分、培养时间,而亲本基因型对胚挽救成苗的影响主要体现在亲本杂交亲和力差异上[27]。本研究中杂交组合‘红宝石无核’ב爱神玫瑰’所获成苗率最高,可能是由于亲本之间亲和性强,有利于合子胚的形成和萌发。母本胚珠的发育程度是胚挽救的关键[7,9,28-30],母本的种子败育类型不同,其幼胚的形成能力有较大的差异[21,27,31-32]。本研究结果显示,胚败育发生过早或可挽救性差的品种如‘克瑞森无核’‘昆香无核’等不宜用作母本材料,这可能与其胚珠发育不良有关。而成苗率较高的品种如‘红宝石无核’‘火焰无核’‘爱神玫瑰’更适宜作为无核香味葡萄胚挽救的母本材料。
利用欧亚种葡萄独特的玫瑰香味,通过胚挽救技术实现香味和无核性状的聚合,培育无核香味新品种是一个有效策略。本研究中父本材料‘玫瑰香’‘阳光玫瑰’‘爱神玫瑰’‘红亚历山大’‘昆香无核’是具有玫瑰香味的品种[9,20,30],这5个父本与同一母本杂交,以‘红亚历山大’‘爱神玫瑰’和‘阳光玫瑰’作为父本的胚挽救效率更高。说明父本基因型对胚挽救效率有一定影响,香味品种中,适合作为父本选育香味种质的为‘红亚历山大’‘爱神玫瑰’‘阳光玫瑰’。
胚萌发培养基的成分对幼胚能否萌发并存活有直接影响,因此,该阶段培养基成分是胚挽救获得成苗的关键影响因素之一。激素配比及浓度影响着成苗方式、植株状态及成苗率。邓向阳等[33]认为在培养基中添加2,4-D、NAA、ZT和KT等生长调节剂并以适当浓度配比,可分别促进诱导离体胚生根及不定芽发生,且在含有1 mg?L-1 2,4-D的培养基中幼胚的萌发率最高可达66.7%[33]。本研究中,无核葡萄胚萌发成苗率在一定范围内的增大可能是NAA、KT及ZT浓度的上升造成的,胚萌发成苗率呈下降趋势可能与2,4-D浓度的增大有关;在以WPM为基础培养基,添加1.0 mg?L-1 KT+0.5 mg?L-1 NAA+1.0 mg?L-1的生长调节剂时,对胚萌发有明显的促进作用。但对不同杂交组合,最适其生长的胚发育培养基和成苗途径,仍需进一步确定[4]。
胚珠离体培养时间同样对胚挽救效果有重要影响[9]。培养时间导致了胚发育形态的不同,不同形态胚接种于胚萌发培养基上,其萌发率及成苗率直接受到影响。本研究结果证明暗培养8周时可取得较好的胚挽救结果,即大部分胚发育至鱼雷形,而接种鱼雷形和子叶形胚可保证胚发育率和成苗率较高。
分子标记辅助选择是利用遗传标记在幼苗期对目标性状进行选择,可缩短育种年限,提高育种效率[34]。目前用于葡萄无核性状鉴定的分子辅助标记有GLSP1-569、SCF27-2000、VMCF7f2-198、SCC8-1018和p3-AGL11-196[35,36,37]。本研究选用GLSP1-569、SCF27-2000和SCC8-10183共3个无核分子标记对杂交组合的亲本和后代进行检测,以‘红宝石无核’为母本的杂交组合后代无核率更高,可为杂交亲本的选择提供依据。下一步可对已获得的杂交后代进行田间性状观察与鉴定,选育同时具有无核和香味的新材料,同时也对无核性状和香味性状的遗传规律进行研究,为无核香味育种提供参考。
4 结论
不同的亲本基因型对胚挽救成苗的影响不同,以‘红宝石无核’‘火焰无核’作母本和以‘红亚历山大’‘爱神玫瑰’‘阳光玫瑰’作父本的杂交组合胚挽救成苗率较高。WPM培养基+KT+ZT可以促进胚挽救的幼胚生根。在101个杂种后代中进行无核基因标记检测,筛选出27个杂种后代同时携带不同无核分子标记的特异条带。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
URLPMID:31824815 [本文引用: 1]
[本文引用: 1]
DOI:10.1007/s11240-014-0656-4URL [本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 5]
[本文引用: 5]
[本文引用: 1]
URLPMID:21807213 [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:30603096 [本文引用: 1]
DOI:10.1007/s00425-018-2986-7URLPMID:30136197 [本文引用: 1]

MAIN CONCLUSION: Five Vitis vinifera sesquiterpene synthases were characterized, two was previously uncharacterized, one being a caryophyllene/cubebene synthase and the other a cadinene synthase. Residue differences with other Vitis sesquiterpene synthases are described. The biochemical composition of grape berries at harvest can have a profound effect on the varietal character of the wine produced. Sesquiterpenes are an important class of volatile compounds produced in grapes that contribute to the flavor and aroma of wine, making the elucidation of their biosynthetic origin an important field of research. Five cDNAs corresponding to sesquiterpene synthase genes (TPSs) were isolated from Shiraz berries and expressed in planta in Nicotiana benthamiana followed by chemical characterization by GC-MS. Three of the TPS cDNAs were isolated from immature berries and two were isolated from ripe Shiraz berries. Two of the investigated enzymes, TPS26 and TPS27, have been previously investigated by expression in E. coli, and the in planta products generally correspond to these previous studies. The enzyme TPS07 differed by eight amino acids (none of which are in the active site) from germacrene B and D synthase isolated from Gewurztraminer grapes and characterized in vitro. Here in planta characterization of VvShirazTPS07 yielded ylangene, germacrene D and several minor products. Two of the enzymes isolated from immature berries were previously uncharacterized enzymes. VvShirazTPS-Y1 produced cadinene as a major product and at least 17 minor sesquiterpenoid skeletons. The second, VvShirazTPS-Y2, was characterized as a caryophyllene/cubebene synthase, a combination of products not previously reported from a single enzyme. Using in silico methods, we identified residues that could play key roles regarding differences in product formation of these enzymes. The first ring closure that is either a 1,10- or 1,11-ring closure is likely controlled by three neighboring amino acids in helices G1, H2, and J. As for many other investigated TPS enzymes, we also observe that only a few residues can account for radical changes in product formation.
[本文引用: 1]
[本文引用: 1]
URLPMID:31704071 [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 2]
[D].
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 2]
综合分析国内外相关文献及作者以往的研究经验,阐述了无核葡萄胚挽救技术的研究现状,讨论了影响无核葡萄胚挽救成功的主要因素,包括亲本选择、取样时期、花期喷施生长调节剂、培养基、外源激素种类与浓度等,并对胚挽救技术在无核葡萄育种上的发展进行了展望。
URL [本文引用: 2]
综合分析国内外相关文献及作者以往的研究经验,阐述了无核葡萄胚挽救技术的研究现状,讨论了影响无核葡萄胚挽救成功的主要因素,包括亲本选择、取样时期、花期喷施生长调节剂、培养基、外源激素种类与浓度等,并对胚挽救技术在无核葡萄育种上的发展进行了展望。
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[D].
[D].
[本文引用: 2]
[D].
[本文引用: 2]
DOI:10.3864/j.issn.0578-1752.2018.21.010URL [本文引用: 1]
【Objective】 The objective of this study is to obtain new cold-resistant seedless grape germplasm and improve the breeding efficiency through embryo rescue in vitro. 【Method】 The different genotypes, basal culture media (MM3 and ER), amino acids (2.5 mmol·L -1cysteine, asparagine, glutamine and serine were added in MM3 medium) were used to investigate the effect on the embryo development rates and seedling rates from ovules of 7 cross combinations. From the parents, the molecular markers GSLP1-569 and SCC8-1018 for seedlessness and the marker S241-717 for cold-resistance were used to identify the strains with specific bands, and then the three markers were used to screen the hybrids by recognizing the specific bands. 【Result】 Of the 7 combinations, 1 168 ovules cultured in MM3 were obtained 331 embryos and 97 hybrid strains. Among these crosses, ‘Ruby Seedless’ × ‘Beichun’ got the highest embryo development rate and seedling rate, which were 46.00% and 17.33%, respectively. Compared with female parents, for ‘Shuangyou’ as male parent and ‘Blush Seedless’ ‘Thompson Seedless’ ‘Kunxiang Seedless’ as female parents, ‘Kunxiang Seedless’ × ‘Shuangyou’ had the highest embryo development rate and seedling rate, which were 45.39% and 16.31%, respectively. For ‘Beibinghong’ as male parent and ‘Blush Seedless’ ‘Beauty Seedless’ ‘Thompson Seedless’ as female parents, ‘Blush Seedless’ × ‘Beibinghong’ had the highest embryo development rate and seedling rate, which were 33.52% and 6.59%, respectively. Different male parents also influenced seedlings formation. The embryo development rates of ‘Blush Seedless’ × ‘Beibinghong’ and ‘Blush Seedless’ × ‘Shuangyou’ were 33.52% and 39.00%, and the seedling rates of them were 6.59% and 9.50%, respectively. The embryo development rates of ‘Thompson Seedless’ × ‘Beibinghong’ and ‘Thompson Seedless’ × ‘Shuangyou’ were 7.64% and 19.26%, and the seedling rates of them were 1.91% and 8.15%, respectively. The rate of embryo development and seedling of ovule cultured in MM3 medium was higher than that of ER medium. For ‘Kunxiang Seedless’ × ‘Shuangyou’, the ovules cultured in MM3 medium with 2.5 mmol·L -1 glutamine showed the highest embryo development rate, which was 59.69%. The seedling rate of ovules cultured in basal media with asparagine and glutamine was 21.43% and 20.93%, respectively. The effect of promoting seedlings development was significant. For ‘Ruby Seedless’ × ‘Beichun’, the ovules in media with 2.5 mmol·L -1 asparagine showed the highest embryo development rate, which was 55.71%. The seedling rate of ovules cultured in basal media with asparagine and glutamine was 21.43% and 22.22%, respectively. The effect of promoting seedlings development was significant. By using the molecular markers GSLP1-569 and SCC8-1018 for seedlessness and the marker S241-717 for cold-resistance, 83 hybrid strains from 6 cross combinations were detected, and there were 49 strains with specific bands of the molecular markers linked to the seedlessness, 55 strains with specific bands of the markers S241-717 for cold-resistance. Importantly, 36 strains with both seedlessness and cold-resistance were identified in this study. 【Conclusion】 ‘Kunxiang Seedless’ and ‘Blush Seedless’ are suitable as the female parents for embryo rescue in vitro of seedless grapes. The embryo rescue efficiency of ‘Shuangyou’ as the male parent is higher than that of ‘Beibinghong’. MM3 is suitable for embryo rescue in vitro, and the addition of amide acids to MM3 culture medium is helpful to the seedling formation. Among the 83 hybrid strains detected by molecular markers, 36 strains may have the seedless and cold-resistant characteristics.
DOI:10.3864/j.issn.0578-1752.2018.21.010URL [本文引用: 1]
【Objective】 The objective of this study is to obtain new cold-resistant seedless grape germplasm and improve the breeding efficiency through embryo rescue in vitro. 【Method】 The different genotypes, basal culture media (MM3 and ER), amino acids (2.5 mmol·L -1cysteine, asparagine, glutamine and serine were added in MM3 medium) were used to investigate the effect on the embryo development rates and seedling rates from ovules of 7 cross combinations. From the parents, the molecular markers GSLP1-569 and SCC8-1018 for seedlessness and the marker S241-717 for cold-resistance were used to identify the strains with specific bands, and then the three markers were used to screen the hybrids by recognizing the specific bands. 【Result】 Of the 7 combinations, 1 168 ovules cultured in MM3 were obtained 331 embryos and 97 hybrid strains. Among these crosses, ‘Ruby Seedless’ × ‘Beichun’ got the highest embryo development rate and seedling rate, which were 46.00% and 17.33%, respectively. Compared with female parents, for ‘Shuangyou’ as male parent and ‘Blush Seedless’ ‘Thompson Seedless’ ‘Kunxiang Seedless’ as female parents, ‘Kunxiang Seedless’ × ‘Shuangyou’ had the highest embryo development rate and seedling rate, which were 45.39% and 16.31%, respectively. For ‘Beibinghong’ as male parent and ‘Blush Seedless’ ‘Beauty Seedless’ ‘Thompson Seedless’ as female parents, ‘Blush Seedless’ × ‘Beibinghong’ had the highest embryo development rate and seedling rate, which were 33.52% and 6.59%, respectively. Different male parents also influenced seedlings formation. The embryo development rates of ‘Blush Seedless’ × ‘Beibinghong’ and ‘Blush Seedless’ × ‘Shuangyou’ were 33.52% and 39.00%, and the seedling rates of them were 6.59% and 9.50%, respectively. The embryo development rates of ‘Thompson Seedless’ × ‘Beibinghong’ and ‘Thompson Seedless’ × ‘Shuangyou’ were 7.64% and 19.26%, and the seedling rates of them were 1.91% and 8.15%, respectively. The rate of embryo development and seedling of ovule cultured in MM3 medium was higher than that of ER medium. For ‘Kunxiang Seedless’ × ‘Shuangyou’, the ovules cultured in MM3 medium with 2.5 mmol·L -1 glutamine showed the highest embryo development rate, which was 59.69%. The seedling rate of ovules cultured in basal media with asparagine and glutamine was 21.43% and 20.93%, respectively. The effect of promoting seedlings development was significant. For ‘Ruby Seedless’ × ‘Beichun’, the ovules in media with 2.5 mmol·L -1 asparagine showed the highest embryo development rate, which was 55.71%. The seedling rate of ovules cultured in basal media with asparagine and glutamine was 21.43% and 22.22%, respectively. The effect of promoting seedlings development was significant. By using the molecular markers GSLP1-569 and SCC8-1018 for seedlessness and the marker S241-717 for cold-resistance, 83 hybrid strains from 6 cross combinations were detected, and there were 49 strains with specific bands of the molecular markers linked to the seedlessness, 55 strains with specific bands of the markers S241-717 for cold-resistance. Importantly, 36 strains with both seedlessness and cold-resistance were identified in this study. 【Conclusion】 ‘Kunxiang Seedless’ and ‘Blush Seedless’ are suitable as the female parents for embryo rescue in vitro of seedless grapes. The embryo rescue efficiency of ‘Shuangyou’ as the male parent is higher than that of ‘Beibinghong’. MM3 is suitable for embryo rescue in vitro, and the addition of amide acids to MM3 culture medium is helpful to the seedling formation. Among the 83 hybrid strains detected by molecular markers, 36 strains may have the seedless and cold-resistant characteristics.
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s001220050976URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}