GWAS Analysis of Hypocotyl Length and Candidate Gene Mining in Cucumber Seedlings
CAI HeXu, BO KaiLiang, ZHOU Qi, MIAO Han, DONG ShaoYun, GU XingFang,, ZHANG ShengPing,Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences, Beijing 100081
Abstract 【Objective】The aim of this study was to identify SNP loci and candidate genes significantly correlated with cucumber hypocotyl length trait, which could provide a theoretical basis for revealing the genetic basis and molecular mechanism of cucumber hypocotyl length trait, and lay a foundation for marker-assisted selection breeding of cucumber hypocotyl length trait.【Method】The natural population including 95 cucumber germplasm was employed in this study, and seedlings were grown in the plastic house in Nankou Experimental Field of Chinese Academy of Agricultural Sciences in spring 2016, spring 2017, autumn 2017 and spring 2018, respectively. The hypocotyl length was measured at the two true leaves stage. Structure 2.3.4 software was used to analyze the population structure, and Haploview software was used to analyze the attenuation of linkage imbalance. Then, the whole genome association analysis of hypocotyl length was carried out based on the optimal model. The important candidate genes related to hypocotyl length were predicted according to the LD interval sequence of the associated SNP loci, and the expression pattern of candidate genes were performed by fluorescence quantitative PCR. 【Result】A total of 8 loci, including Hl1.1, Hl1.2, Hl2.1, Hl3.1, Hl3.2, Hl4.1, Hl5.1 and Hl6.1, were detected on Chr. 1, 2, 3, 4 and 5, respectively. Five of them, Hl2.1, Hl3.1, Hl3.2, Hl5.1 and Hl6.1, were detected repeatedly in two or more different environments. By analyzing the LD interval sequences of the associated SNP loci, eight candidate genes, Csa1G074930, Csa1G475980, Csa2G381650, Csa3G141820, Csa4G051570, Csa3G627150, Csa5G174640 and Csa6G362970, were predicted, which were related to cucumber hypocotyl length. Some of the candidate genes involved in regulating plant photomorphogenesis, ubiquitination, and hormone signaling pathway. And some of them were downstream genes regulating cell growth, development and cell size, thus they directly regulated hypocotyl length. Thus, the varied distribution of above genes in different cucumber materials resulted in the different hypocotyl length cucumber germplasm. The organic distribution of polygenes in different cucumber materials formed cucumber germline with different Hypocotyl length. Gene expression analysis showed that Csa1G074930, Csa1G475980, Csa2G381650, Csa4G051570 and Csa5G174640 were highly expressed in short hypocotyl materials and Csa3G141820 and Csa3G627150 were highly expressed in long hypocotyl materials.【Conclusion】Eight SNP loci linked with hypocotyl length, Hl1.1, Hl1.2, Hl2.1, Hl3.1, Hl3.2, Hl4.1, Hl5.1 and Hl6.1, were detected in this study. Eight candidate genes regulating hypocotyl length were predicted, including Csa1G074930, Csa1G475980, Csa2G381650, Csa3G141820, Csa4G051570, Csa3G627150, Csa5G174640 and Csa6G362970. Keywords:cucumber;hypocotyl length;genome-wide association study;candidate gene
PDF (1914KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 蔡和序, 薄凯亮, 周琪, 苗晗, 董邵云, 顾兴芳, 张圣平. 黄瓜幼苗下胚轴长度GWAS分析及候选基因挖掘[J]. 中国农业科学, 2020, 53(1): 122-132 doi:10.3864/j.issn.0578-1752.2020.01.012 CAI HeXu, BO KaiLiang, ZHOU Qi, MIAO Han, DONG ShaoYun, GU XingFang, ZHANG ShengPing. GWAS Analysis of Hypocotyl Length and Candidate Gene Mining in Cucumber Seedlings[J]. Scientia Acricultura Sinica, 2020, 53(1): 122-132 doi:10.3864/j.issn.0578-1752.2020.01.012
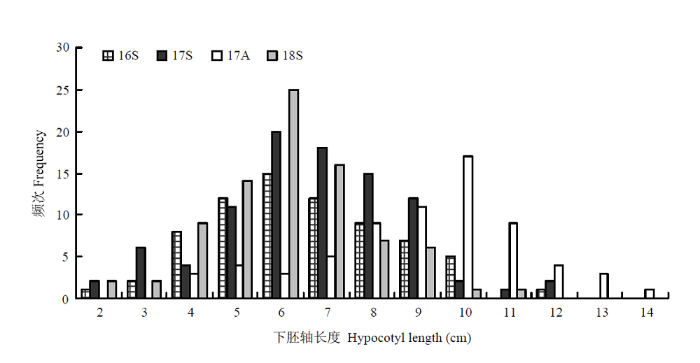
将苗期调查的表型数据和过滤后构建的高质量SNP矩阵,利用基于R的GAPITR软件包[21],通过混合线性模型(mixed linear model,MLM)进行关联分析。利用ggplot2软件绘制Quantile-Quantile散点图(QQ plot)[22],并在此基础上利用QQman绘制曼哈顿图[23],显示关联分析检测到的与目标性状显著相关的标记位点。对95份黄瓜的苗期下胚轴性状进行全基因组关联分析。
Table 3 表3 表3黄瓜下胚轴长度统计分析 Table 3Statistical analysis of hypocotyl length in cucumber
环境 Environment
均值±标准差 Mean±SD
最小值 Minimum
50%分位数 50% quantile
最大值 Max
变异系数 CV (%)
16S
6.02±1.95
1.75
5.84
11.25
32.32
17S
6.19±2.06
0.60
6.16
11.66
33.27
17A
8.53±2.24
3.28
9.02
13.18
26.28
18S
5.65±1.70
1.30
5.59
10.36
30.17
16S:2016年春季;17S:2017年春季;17A:2017年秋季;18S:2018年春季。下同 16S: Spring of 2016; 17S: Spring of 2017; 17A: Autumn of 2017; 18S: Spring of 2018. The same as below
Table 6 表6 表6qPCR材料苗期下胚轴长度 Table 6The hypocotyl length of qPCR materials at seedling stage
品种 Variety
时期1 Stage 1
时期2 Stage 2
时期3 Stage 3
时期4 Stage 4
时期5 Stage 5
CG9
5.23±0.25b
7.03±0.21b
9.77±0.25b
11.07±0.25b
12.27±0.21a
CG11
4.4±0.22c
6.27±0.21c
8.07±0.17c
9.2±0.16c
9.97±0.29b
CG19
3.17±0.21d
3.1±0.08d
3.67±0.12f
4.97±0.21f
6.6±0.08d
CG44
2.8±0.08e
3.07±0.25d
4.3±0.16e
6±0.24e
6.57±0.17d
CG54
2.1±0.08f
2.23±0.12e
3.27±0.25f
3.47±0.21g
4.17±0.12e
CG98
1.63±0.12g
3.27±0.17d
3.5±0.16f
4.1±0.29g
4.67±0.12e
CG106
5.43±0.12b
6.03±0.12c
6.87±0.12d
8.2±0.16d
8.97±0.21c
CG112
6.23±0.12a
8.13±0.12a
11.77±0.21a
12.53±0.12a
12.67±0.29a
时期1:子叶展平;时期2:第一片真叶展平;时期3:一叶一心;时期4:第二片真叶展平;时期5:两叶一心;表中数据单位均为(cm);不同小写字母表示不同品种间差异达0.05显著水平(Duncan’s 法) Stage 1: the cotyledon stage; Stage 2: the one true leaf stage; Stage 3: the one leaf and one bud stage; Stage 4: the two true leaves stage; Stage 5:the two true leaves and one bud stage; All data units in the table are (cm); Different lowercase letters indicate significant difference between different breeds at 0.05 level (Duncan's method)
CG9、CG11、CG106、CG112均为长下胚轴材料;CG19、CG44、CG54、CG98均为短下胚轴材料;每个候选基因中选取表达量最低的材料,其相对表达量设为1 Fig. 6Analysis of relative expression levels of candidate genes
CG9, CG11, CG106 and CG112 are long hypocotyl materials. CG19, CG44, CG54 and CG98, are all short hypocotyl materials. The material with the lowest expression level was selected from each candidate gene, and its relative expression level was set as 1
LINY, SCHIEFELBEINJ . Embryonic control of epidermal cell patterning in the root and hypocotyl of Arabidopsis Development, 2001,128(19):3697-3705. [本文引用: 1]
JIN ZA, SI LT, LI DD, GAOX . Genetic analysis of hypocotyl length in cucumber seedling leaves under weak light Jiangsu agricultural science, 2009(3):158-161.(in Chinese) [本文引用: 1]
LI DD, SI LT, LUO XM, LIT . Genetic analysis of hypocotyl traits in cucumber seedling under low light stress Journal of Northwestern Agricultural and Forestry University (Natural Science Edition), 2009,37(11):113-119. (in Chinese) [本文引用: 1]
ZHANG GY, SI LT, LI DD . QTL analysis for hypocotyl traits of cucumber seedlings under low light stress Acta Horticulturae Sinica, 2011,38(2):295-302. (in Chinese) [本文引用: 1]
ZOU SC, LI DD, SUN BH, ZHANG CF, YU SQ . Response of hypocotyl of cucumber seedlings to light stress Anhui Agricultural Science Bulletin, 2015,21(19):29-30. (in Chinese) [本文引用: 1]
SONG JL, CAOK, HAO YW, SONG SW, SUW, LIU HC . Hypocotyl elongation is regulated by supplemental blue and red light in cucumber seedling Gene, 2019,707:117-125. [本文引用: 2]
XIAO SQ, WANG BH, QUM, GAO LH . Effect of supplementary light intensity on quality of winter-spring cucumber seedling in solar greenhouse Chinese Vegetables, 2018(10):40-45. (in Chinese) [本文引用: 1]
ZHANG ZM, LU JC, QI XH, XU XW, CHEN XH, XUQ . Study on genetic effects of hypocotyl length in cucumber seedlings under high temperature Molecular Plant Breeding, 2019,17(4):1326-1332. (in Chinese) [本文引用: 2]
DONG CJ, CAON, WANG LL, ZHANG HX, WANG HF, TAI LL, SHANG QM . Regulatory roles of cotyledon-generated auxin in adventitious root formation on the hypocotyls of cucumber seedling Acta Horticulturae Sinica, 2016,43(10):1929-1940. (in Chinese) [本文引用: 1]
LOPEZ-JUEZE, KOBAYASHIM, SAKURAIA, KAMIYAY, KENDRICK RE . Phytochrome, gibberellins, and hypocotyl growth (a study using the cucumber (Cucumis sativus L.) long hypocotyl mutant) Plant Physiology, 1995,107(1):131-140. [本文引用: 1]
BO KL, WANGH, PAN YP, BEHERA TK, PANDEYS, WEN CL, WANG YH, SIMON PW, LI YH, CHEN JF, WENG YQ . SHORT HYPOCOTYL 1 encodes a SMARCA3-like chromatin remodeling factor regulating elongation Plant Physiology, 2016,172:1273-1292. [本文引用: 2]
MIAOH, GU XF, ZHANG SP, ZHANG ZH, HUANG SW, WANGY, FANG ZY . Mapping QTLs for seedling-associated traits in cucumber Acta Horticulturae Sinica, 2012,39(5):879-887. (in Chinese) [本文引用: 2]
ATWELLS, HUANG YS, VILHJALMSSON BJ, WILLEMSG, HORTONM, LIY, MENGD, PLATTA, TARONE AM, HU TT, JIANGR, MULIYATI NW, ZHANGX, AMER MA, BAXTERI, BRACHIB, CHORYJ, DEANC, DEBIEUM, DE MEAUXJ, ECKER JR, FAUREN, KNISKERN JM, JONES JD, MICHAELT, NEMRIA, ROUXF, SALT DE, TANGC, TODESCOM, TRAW MB, WEIGELD, MARJORAMP, BOREVITZ JO, BERGELSONJ, NORDBORGM . Genome-wide association study of 107 phenotypes in Arabidopsis thaliana inbred lines Nature, 2010,465(7298):627-631. [本文引用: 1]
BRACHIB, MORRIS GP, BOREVITZ JO . Genome-wide association studies in plants: The missing heritability is in the field Genome Biology, 2011,12(10):232. [本文引用: 1]
WANGM, YAN JB, ZHAO JR, SONGW, ZHANG XB, XIAO YN, ZHENG YL . Genome-wide association study (GWAS) of resistance to head smut in maize Plant Science, 2012,196:125-131. [本文引用: 1]
HUANG XH, WEI XH, SANGT, ZHAO QA, FENGQ, ZHAOY, LI CL, ZHU CR, LU TT, ZHANG ZW, LIM, FAN DL, GUO YL, WANG AH, WANGL, DENG LW, LI WJ, LU YQ, WENG QJ, LIU KY, HUANGT, ZHOU TY, JING YF, LIW, LINZ, BUCKLER ES, QIANQ, ZHANG QF, LI JY, HANB . Genome-wide association studies of 14 agronomic traits in rice landraces Nature Genetics, 2010,42(11):961-967. [本文引用: 1]
QI JJ, LIUX, SHEND, MIAOH, XIE BY, LI XX, ZENGP, WANG SH, SHANGY, GU XF, DU YC, LIY, LINT, YUAN JH, YANG XY, CHEN JF, CHEN HM, XIONG XY, HUANGK, FEI ZJ, MAO LY, TIANL, STADLERT, RENNER SS, KAMOUNS, LUCAS WJ, ZHANG ZH, HUANG SW . A genomic variation map provides insights into the genetic basis of cucumber domestication and diversity Nature Genetics, 2013,45(12):1510-1515. [本文引用: 4]
YANG JA, LEE SH, GODDARD ME, VISSCHER PM . A tool for genome-wide complex trait analysis The American Journal of Human Genetics, 2011,88(1):76-82. [本文引用: 1]
BARRETT JC, FRYB, MALLERJ, DALY MJ . Haploview: Analysis and visualization of LD and haplotype maps Bioinformatics, 2005,21(2):263-265. [本文引用: 1]
ZHANG ZW, ERSOZE, LAI CQ, TODHUNTER RJ, TIWARI HK, GORE MA, BRADBURY PJ, YU JM, ARNETT DK, ORDOVAS JM, BUCKLER ES . Mixed linear model approach adapted for genome-wide association studies Nature Genetics, 2010,42(4):355-360. [本文引用: 1]
BRADBURY PJ, ZHANG ZW, KROON DE, CASSTEVENS TM, RAMDOSSY, BUCKLER ES . TASSEL: Software for association mapping of complex traits in diverse samples Bioinformatics, 2007,23(19):2633-2635. [本文引用: 1]
ZHANG ZW, ERSOZE, LAI CQ, TODHUNTER RJ, TIWARI HK, GORE MA, BRADBURY PJ, YU JM, ARNETT DK, ORDOVAS JM, BUCKLER ES . Mixed linear model approach adapted for genome-wide association studies Nature Genetics, 2010,42(4):355-360. [本文引用: 1]
ZHAO QX, YUANS, WANGX, ZHANG YL, ZHUH, LU CM . Restoration of mature etiolated cucumber hypocotyl cell wall susceptibility to expansin by pretreatment with fungal pectinases and egta in vitro Plant Physiology, 2008,147(4):1874-1885. [本文引用: 1]
KOEDAS, SATOK, SAITOH, NAGANO AJ, YASUGIM, KUDOHH, TANAKAY . Mutation in the putative ketoacyl-ACP reductase CaKR1 induces loss of pungency in Capsicum Theoretical and Applied Genetics, 2019,132(1):65-80. [本文引用: 1]
ZHAO LM, PENGT, CHEN CY, JI RJ, GU DC, LI TT, ZHANG DD, TU YS, WU KQ, LIU XC . HY5 interacts with the histone deacetylase HDA15 to repress hypocotyl cell elongation in photomorphogenesis Plant Physiology, 2019,180(3):1450-1466. [本文引用: 1]
RAIJMAKERSR, NOORDMAN YE, VAN VENROOIJ WJ, PRUIJN G JM . Protein-protein interactions of hCsl4p with other human exosome subunits Journal of Molecular Biology, 2002,315(4):809-818. [本文引用: 1]
BREMBUT, WINGEP, SEEMM, BONES AM . NAPP and PIRP encode subunits of a putative wave regulatory protein complex involved in plant cell morphogenesis The Plant Cell, 2004,16(9):2335-2349. [本文引用: 1]
 ,, 张圣平
,, 张圣平
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT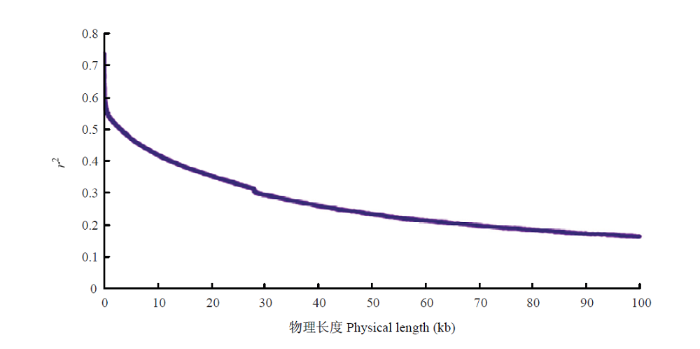 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}