Pathogenicity and Gene Expression Pattern of the Exocrine Protein LtGH61A of Grape Canker Fungus
PENG JunBo, LI XingHong, ZHANG Wei, ZHOU Ying, HUANG JinBao, YAN JiYe,Institute of Plant and Environment Protection, Beijing Academy of Agriculture and Forestry Sciences/Beijing Key Laboratory of Environmental Friendly Management of Diseases and Pests of North China Fruits, Beijing 100097
Abstract 【Objective】Grape canker disease, caused by Botryosphaeria genus fungi, occurs in a wide range of grape-producing areas in China and seriously threatens the yield and quality of grape. The objective of this study is to analyze the function of a hypothetical exocrine protein, LtGH61A, in grape canker fungus Lasiodiplodia theobromae, and to lay a foundation for in-depth analysis of the pathogenic mechanism and disease control of grape canker fungus.【Method】The signal peptide of LtGH61A protein was predicted by SignalP 4.0. The function of LtGH61A protein was predicted by the homologous comparison and functional annotation. The exocrine characteristic of LtGH61A protein was analyzed by yeast complementary experiment. The quantitative real-time PCR (qRT-PCR) was used to analyze the expression of LtGH61A in vegetative hyphae and different infection processes. The expression of LtGH61A was inhibited through RNA interference (RNAi). The effect of LtGH61A protein on the pathogenicity of L. theobromae was analyzed by in vitro inoculation test of grape shoots. The effect of LtGH61A protein on the hyphal growth rate of L. theobromae was analyzed by comparing the colony diameter.【Result】Amino acid sequence analysis predicts that the N-terminal of the LtGH61A protein contains a signal peptide with a length of 18 amino acids. The gene function annotation suggests that LtGH61A belongs to glycoside hydrolase family 61 (GH61) and can degrade cellulose as a substrate. Yeast complementary experiments showed that the signal peptide of LtGH61A protein could guide the secretion of invertase of yeast YTK12. Compared with the vegetative hyphae, the expression of LtGH61A was increased significantly at the infectious stages, and the mRNA accumulation of LtGH61A at 48 h post inoculation was 19 times of that in the vegetative hyphae. Moreover, RNAi lines were constructed for LtGH61A and two lines RNAi-LtGH61A1 and RNAi-LtGH61A2 were confirmed by qRT-PCR. The results of in vitro inoculation test of wild-type and RNAi transformants on wounded grape shoots showed that the lesion length caused by both RNAi-LtGH61A1 and RNAi-LtGH61A2 was significantly shorter than that of wild type (WT) CSS-01s, which was about 55% of WT, indicating that LtGH61A affected the pathogenicity of L. theobromae. The colony diameter comparison showed that compared with WT, the colony diameter of RNAi-LtGH61A1 and RNAi-LtGH61A2 transformants became smaller, about 85% of WT, indicating that LtGH61A affected the hyphal growth rate of L. theobromae.【Conclusion】LtGH61A affects the pathogenicity and hyphal growth of grape canker pathogen. LtGH61A protein can be secreted outside the cell. The expression level of LtGH61A during infectious stages is significantly increased, suggesting that LtGH61A can destroy the host plant tissue by exerting its own enzyme activity function, thus promoting pathogen infection. Keywords:grape canker fungus;exocrine protein;pathogenicity;expression pattern;qRT-PCR;RNA interference (RNAi)
PDF (1781KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 彭军波, 李兴红, 张玮, 周莹, 黄金宝, 燕继晔. 葡萄溃疡病菌外泌蛋白LtGH61A的致病力及基因表达模式[J]. 中国农业科学, 2019, 52(24): 4518-4526 doi:10.3864/j.issn.0578-1752.2019.24.007 PENG JunBo, LI XingHong, ZHANG Wei, ZHOU Ying, HUANG JinBao, YAN JiYe. Pathogenicity and Gene Expression Pattern of the Exocrine Protein LtGH61A of Grape Canker Fungus[J]. Scientia Acricultura Sinica, 2019, 52(24): 4518-4526 doi:10.3864/j.issn.0578-1752.2019.24.007
供试菌株及植物:可可毛色二孢野生型(WT)菌株CSS-01s由笔者实验室保存,酵母菌株YTK12由中国农业大学孙文献老师馈赠,大肠杆菌感受态细胞Trans1-T1购自北京全式金生物技术有限公司。病原菌接种试验所用葡萄品种为‘夏黑’,顺义香逸葡萄园。真菌转化子置于灭菌的1.5 mL Eppendorf管中,保存于4℃冰箱,质粒载体保存于-20℃冰箱。
供试药剂及仪器:各种限制性内切酶及T4 DNA连接酶,New England Biolabs公司;高保真Taq聚合酶,北京擎科新业生物技术有限公司;氨苄青霉素、硫酸腺嘌呤,北京索莱宝科技有限公司;葡萄糖,国药集团化学试剂有限公司;抗霉素A,Abcam公司,英国;聚乙二醇3350及棉子糖,Sigma-aldrich公司,美国;Trizon试剂,Invitrogen,美国;植物RNA快速提取试剂盒,北京艾德莱生物科技有限公司;反转录试剂盒,北京全式金生物技术有限公司;2×RealStar Green Fast Mixture with ROX II,北京康润诚业生物科技有限公司;酵母提取物、蛋白胨、不含氨基酸的酵母氮源、琼脂,Becton, Dickinson and Company,美国;YeastmakerTM Yeast Transformation System 2试剂盒、tryptophan dropout supplement,Clonthch,日本;Applied Biosystems 7500仪器,ABI公司,美国;冷冻高速离心机,HiKOKI,日本。
A:菌落形态The colony morphology;B:菌落直径比较 Statistical analysis of the colony diameter;C:在葡萄枝干上形成的病斑The lesion on grape shoot;D:病斑长度统计Statistical analysis of the lesion length。**:P<0.01 Fig. 3The growth rate and pathogenicity of LtGH61A RNAi transformants
VH:营养菌丝 Vegetative hyphae;24—72 h:接种后24、48、72 h 24, 48 and 72 h after inoculation。**:P<0.01 Fig. 4The expression level of LtGH61A in vegetative hyphae and different infection processes
ZHANGW, LI XH, GUO FF, LIUM, HUANG JB, YAN JY . Establishment and application of duplex PCR assay for grape canker pathogens Botryosphaeria dothidea and Neofusicoccum parvum. Journal of Plant Protection, 2017,44(4):636-642. (in Chinese) [本文引用: 1]
YAN JY, XIEY, YAO SW, WANG ZY, LI XH . Characterization of Botryosphaeria dothidea, the causal agent of grapevine canker in China. Australasian Plant Pathology, 2012,41(4):351-357. [本文引用: 5]
YAN JY, XIEY, ZHANGW, WANGY, LIU JK, HYDE KD, SEEM RC, ZHANG GZ, WANG ZY, YAO SW, BAI XJ, DISSANAYAKE AJ, PENG YL, LI XH . Species of Botryosphaeriaceae involved in grapevine dieback in China Fungal Diversity, 2013,61(1):221-236. [本文引用: 3]
YAN JY, LI XH, KONG FF, WANG ZY, GONG LZ, HE HP . Occurrence of grapevine trunk disease caused by Botryosphaeria rhodina in China. Plant Disease, 2011,95(2):219. [本文引用: 3]
YAN JY, PENG YL, XIEY, LI XH, YAO SW, TANG ML, WANG ZY . First report of grapevine trunk disease caused by Botryosphaeria obtusa in China. Plant Disease, 2011,95(5):616. [本文引用: 2]
DISSANAYAKE AJ, ZHANGW, LIUM, CHUKEATIROTEE, YAN JY, LI XH, HYDE KD . Lasiodiplodia pseudotheobromae causes pedicel and peduncle discolouration of grapes in China. Australasian Plant Disease Notes, 2015,10:21. [本文引用: 2]
DISSANAYAKE AJ, ZHANGW, LI XH, ZHOUY, CHETHANAT, CHUKEATIROTEE, HYDE KD, YAN JY, ZHANG GZ, ZHAO WS . First report ofNeofusicoccum mangiferae associated with grapevine dieback in China. Phytopathologia Mediterranea, 2015,54(2):414-419. [本文引用: 2]
ELGOORANI MA, ELMELEIGI MA . Dieback of grapevine by Botryodiplodia theobromae Pat. in Egypt. Phytopathologia Mediterranea, 1972,11(3):210-211. [本文引用: 2]
PHILLIPS A JL . Botryosphaeria dothidea and other fungi associated with excoriose and dieback of grapevines in Portugal. Journal of Phytopathology, 1998,146(7):327-332. [本文引用: 1]
LARIGNONP, FULCHICR, CEREL, DUBOSB . Observation on black dead arm in French vineyards Phytopathologia Mediterranea, 2001,40(Suppl.):S336-S342. [本文引用: 1]
PHILLIPS A JL . Botryosphaeria species associated with diseases of grapevines in Portugal. Phytopathologia Mediterranea, 2002,41(1):3-18. [本文引用: 1]
TAYLORA, HARDY GE, WOODP, BURGESST . Identification and pathogenicity of Botiyosphaeria species associated with grapevine decline in Western Australia. Australasian Plant Pathology, 2005,34(2):187-195. [本文引用: 1]
URBEZ-TORRES JR . The status of Botryosphaeriaceae species infecting grapevines Phytopathologia Mediterranea, 2011,50(Suppl.):S5-S45. [本文引用: 1]
VAN NIEKERK JM, FOURIE PH, HALLEENF, CROUS PW . Botryosphaeria spp. as grapevine trunk pathogens. Phytopathologia Mediterranea, 2006,45(Suppl.):S43-S54. [本文引用: 1]
úRBEZ-TORRES JR, LEAVITT GM, GUERRERO JC, GUEVARAJ, GUBLER WD . Identification and pathogenicity of Lasiodiplodia theobromae and Diplodia seriata, the causal agents of bot canker disease of grapevines in Mexico. Plant Disease, 2008,92(4):519-529. [本文引用: 1]
FANK, WANGJ, FUL, ZHANG GF, WU HB, FENGC, QU JL . Baseline sensitivity and control efficacy of pyraclostrobin againstBotryosphaeria dothidea isolates in China. Plant Disease, 2019,103(7):1458-1463. [本文引用: 1]
PAN JL, HAOX, YAO HW, GE KK, MAL, MAW . Matrine inhibits mycelia growth of Botryosphaeria dothidea by affecting membrane permeability. Journal of Forestry Research, 2019,30(3):1105-1113. [本文引用: 1]
TROTEL-AZIZP, ABOU-MANSOURE, COURTEAUXB, RABENOELINAF, CLéMENTC, FONTAINEF, AZIZA . Bacillus subtilis PTA-271 counteracts Botryosphaeria dieback in grapevine, triggering immune responses and detoxification of fungal phytotoxins. Frontiers in Plant Science, 2019,10:25. [本文引用: 1]
COBOSR, BARREIROC, MATEOS RMCOQUE J JR . Cytoplasmic- and extracellular-proteome analysis ofDiplodia seriata: A phytopathogenic fungus involved in grapevine decline. Proteome Science, 2010,8:46. [本文引用: 1]
PAOLINELLI-ALFONSOM, VILLALOBOS-ESCOBEDO J M, ROLSHAUSENP, HERRERA-ESTRELLAA, GALINDO-SáNCHEZC, LóPEZ-HERNáNDEZ JF, HERNANDEZ-MARTINEZR . Global transcriptional analysis suggests Lasiodiplodia theobromae pathogenicity factors involved in modulation of grapevine defensive response. BMC Genomics, 2016,17:615. [本文引用: 1]
YAN JY, ZHAO WS, CHENZ, XING QK, ZHANGW, CHETHANA K WT, XUE MF, XU JP, PHILLIPS A JL, WANGY, LIU JH, LIUM, ZHOUY, JAYAWARDENA RS, MANAWASINGHE IS, HUANG JB, QIAOGH, FU CY, GUO FF, DISSANAYAKE AJ, PENG YL, HYDE KD, LI XH . Comparative genome and transcriptome analyses reveal adaptations to opportunistic infections in woody plant degrading pathogens of Botryosphaeriaceae DNA Research, 2018,25(1):87-102. [本文引用: 4]
MARTOSS, ANDOLFIA, LUQUEJ, MUGNAIL, SURICOG, EVIDENTEA . Production of phytotoxic metabolites by five species of Botryosphaeriaceae causing decline on grapevines, with special interest in the species Neofusicoccum luteum and N. parvum. European Journal of Plant Pathology, 2008,121(4):451-461. [本文引用: 1]
EVIDENTEA, PUNZOB, ANDOLFIA, CIMMINOA, MELCKD, LUQUEJ . Lipophilic phytotoxins produced by Neofusicoccum parvum, a grapevine canker agent. Phytopathologia Mediterranea, 2010,49(1):74-79. [本文引用: 1]
ANDOLFIA, MUGNAIL, LUQUEJ, SURICOG, CIMMINOA, EVIDENTEA . Phytotoxins produced by fungi associated with grapevine trunk diseases Toxins, 2011,3(12):1569-1605. [本文引用: 1]
ABOU-MANSOURE, DéBIEUX JL, RAMíREZ-SUEROM, BéNARD-GELLONM, MAGNIN-ROBERTM, SPAGNOLOA, CHONGJ, FARINES, BERTSCHC, L'HARIDONF, SERRANOM, FONTAINEF, REGOC, LARIGNONP . Phytotoxic metabolites from Neofusicoccum parvum, a pathogen of Botryosphaeria dieback of grapevine. Phytochemistry, 2015,115:207-215. [本文引用: 1]
VAAJE-KOLSTADG, HORN SJ, VAN AALTEN D MF, SYNSTADB, EIJSINK V GH . The non-catalytic chitin-binding protein CBP21 from Serratia marcescens is essential for chitin degradation. The Journal of Biological Chemistry, 2005,280(31):28492-28497. [本文引用: 1]
LANGSTON JA, SHAGHASIT, ABBATEE, XUF, VLASENKOE, SWEENEY MD . Oxidoreductive cellulose depolymerization by the enzymes cellobiose dehydrogenase and glycoside hydrolase 61 Applied and Environmental Microbiology, 2011,77(19):7007-7015. [本文引用: 3]
KARKEHABADIS, HANSSONH, KIMS, PIENSK, MITCHINSONC, SANDGRENM . The first structure of a glycoside hydrolase family 61 member, Cel61B from Hypocrea jecorina, at 1.6 ? resolution. Journal of Molecular Biology, 2008,383(1):144-154. [本文引用: 1]
SALINASA, VEGAM, LIENQUEO ME, GARCIAA, CARMONAR, SALAZARO . Cloning of novel cellulases from cellulolytic fungi: Heterologous expression of a family 5 glycoside hydrolase from Trametes versicolor in Pichia pastoris. Enzyme and Microbial Technology, 2011,49(6/7):485-491. [本文引用: 1]
KOSEKIT, MESEY, FUSHINOBUS, MASAKIK, FUJIIT, ITOK, SHIONOY, MURAYAMAT, IEFUJIH . Biochemical characterization of a glycoside hydrolase family 61 endoglucanase from Aspergillus kawachii. Applied Microbiology and Biotechnology, 2008,77(6):1279-1285. [本文引用: 2]
RAJESHWARIR, JHAG, SONTI RV . Role of an in planta-expressed xylanase of Xanthomonas oryzae pv.oryzae in promoting virulence on rice. Molecular Plant-Microbe Interactions, 2005,18(8):830-837. [本文引用: 1]
JACOBS KA, COLLINS-RACIE LA, COLBERTM, DUCKETTM, GOLDEN-FLEETM, KELLEHERK, KRIZR, LAVALLIE ER, MERBERGD, SPAULDINGV, STOVERJ, WILLIAMSON MJ, MCCOY JM . A genetic selection for isolating cDNAs encoding secreted proteins Gene, 1997,198(1/2):289-296. [本文引用: 1]
FANGA, HAN YQ, ZHANGN, ZHANGM, LIU LJ, LIS, LUF, SUN WX . Identification and characterization of plant cell death- inducing secreted proteins from Ustilaginoidea virens. Molecular Plant-Microbe Interactions, 2016,29(5):405-416. [本文引用: 2]
KONG XJ, SHIJ, KONG FF, WANG ZY, ZHANGH . Identification of the signal peptide of candidate effector protein RXLR5 from Plasmopara viticola. Plant Protection, 2016,42(1):40-44. (in Chinese) [本文引用: 1]
GUB, KALE SD, WANG QH, WANG DH, PAN QN, CAOH, MENG YL, KANG ZS, TYLER BM, SHAN WX . Rust secreted protein Ps87 is conserved in diverse fungal pathogens and contains a RXLR-like motif sufficient for translocation into plant cells PLoS ONE, 2011,6(11):e27217. [本文引用: 1]
DOUD, KALE SD, WANGX, JIANG RH, BRUCE NA, ARREDONDO FD, ZHANGX, TYLER BM . RXLR-mediated entry of Phytophthora sojae effector Avr1b into soybean cells does not require pathogen-encoded machinery. The Plant Cell, 2008,20(7):1930-1947. [本文引用: 1]
 ,北京市农林科学院植物保护环境保护研究所/北方果树病虫害绿色防控北京市重点实验室,北京100097
,北京市农林科学院植物保护环境保护研究所/北方果树病虫害绿色防控北京市重点实验室,北京100097
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT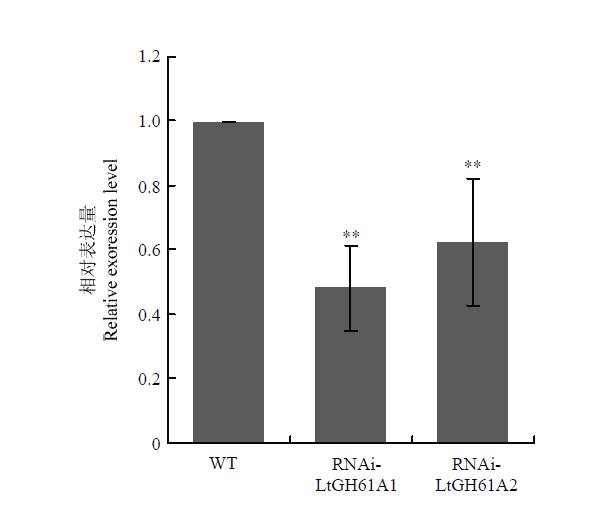 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT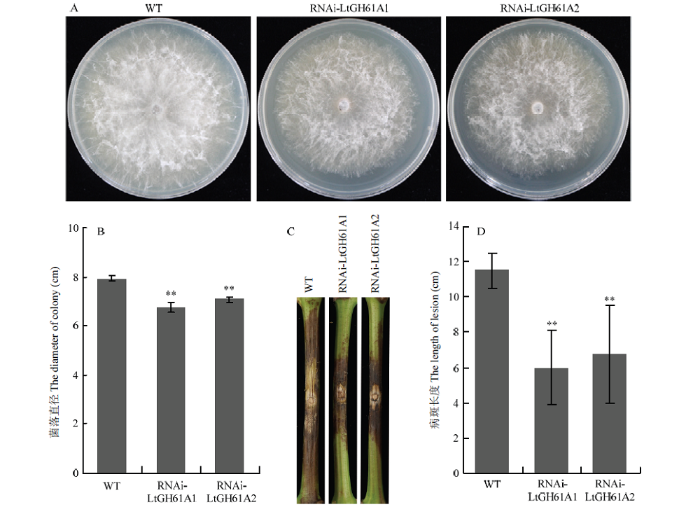 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT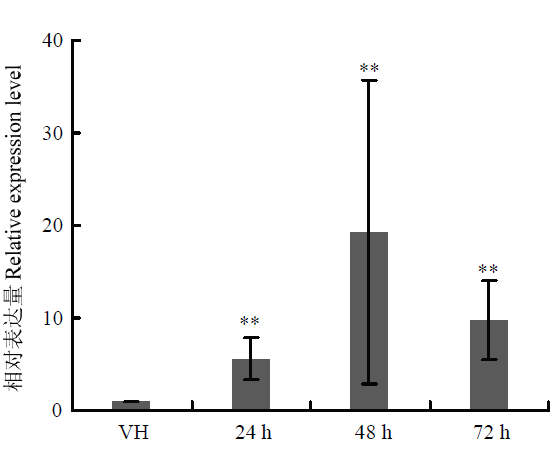 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}