 ,, 郭宁, 张海剑河北省农林科学院植物保护研究所/农业部华北北部作物有害生物综合治理重点实验室/河北省农业有害生物综合防治工程技术研究中心, 河北保定 071000
,, 郭宁, 张海剑河北省农林科学院植物保护研究所/农业部华北北部作物有害生物综合治理重点实验室/河北省农业有害生物综合防治工程技术研究中心, 河北保定 071000Genetic Diversity Analysis of Curvularia lunata in Summer Maize in Huang-Huai-Hai Region
CHANG JiaYing, LIU ShuSen, MA HongXia, SHI Jie,, GUO Ning, ZHANG HaiJianPlant Protection Institute of Hebei Academy of Agricultural and Forestry Sciences/Key Laboratory of IPM on Crops in Northern Region of North China, Ministry of Agriculture/IPM Centre of Hebei Province, Baoding 071000, Hebei通讯作者:
收稿日期:2018-10-27接受日期:2018-12-23网络出版日期:2019-03-01
| 基金资助: |
Received:2018-10-27Accepted:2018-12-23Online:2019-03-01
作者简介 About authors
常佳迎,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (1676KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
常佳迎, 刘树森, 马红霞, 石洁, 郭宁, 张海剑. 黄淮海地区夏玉米弯孢叶斑病菌遗传多样性分析[J]. 中国农业科学, 2019, 52(5): 822-836 doi:10.3864/j.issn.0578-1752.2019.05.005
CHANG JiaYing, LIU ShuSen, MA HongXia, SHI Jie, GUO Ning, ZHANG HaiJian.
0 引言
【研究意义】玉米弯孢叶斑病(Curvularia leaf spot of maize)是主要由新月弯孢(Curvularia lunata )引起的叶部病害,20世纪80年代以来,在辽宁、吉林、北京、天津、河北、山东、河南、安徽、江苏、云南等玉米种植区相继发生,已成为继大斑病、小斑病之后的又一重要叶部病害,对玉米生产造成严重影响[1,2,3,4]。2013年在安徽北部和河南东南部发生较严重,病级为3—7级,严重地块可达9级;2017年在河北省和山东中部发生普遍,但发病程度较轻,为1—3级;在安徽北部发生较重,病级普遍为3—7级,其他年份和地区仅见零星病斑。该病害的发生具有间歇性,其在我国的发生和流行规律目前尚不十分明确。研究玉米弯孢叶斑病不同年份和不同发生区域病原菌群体的遗传多样性,有助于了解各地病原菌的遗传结构,明确年度间不同地理种群间的遗传关系,可为研究病害的发生规律和流行趋势提供重要的基础信息,并为病害的田间防控和预测预报提供重要依据。【前人研究进展】物种DNA的多态性可通过多种分子标记技术快速、高效地检测出来,如RAPD、RFLP、AFLP、ISSR、SSR等[5,6,7]。其中ISSR分子标记技术可灵敏高效地检测基因组DNA的多态性,与RAPD分子标记技术相比,其具有很好的稳定性和多态性,成本低、操作简单,适合大样本的检测,现已在许多物种的遗传多样性与遗传结构、物种来源、物种形成和种质鉴定等领域广泛应用[8,9,10,11,12]。在弯孢叶斑病菌遗传多样性方面,范永山等[13]研究表明,河北省新月弯孢种内存在较大的遗传差异;张欣芳等[14]对东北地区的弯孢菌遗传多样性进行分析,发现病菌的遗传多样性与致病性有一定关系,与地理来源无密切关系;龚国淑等[15]对10个省(市)的新月弯孢菌株进行遗传多样性分析,明确了新月弯孢种内菌株的遗传多样性与致病性相关,且与地理来源无直接关系,推测可能与各地频繁调运种子所导致的病菌在各地扩散有关;王晓东等[16]使用12个ISSR引物和4个UP-PCR引物对采集自辽宁和安徽的玉米弯孢叶斑病菌进行扩增,发现ISSR技术更适合对弯孢病菌的遗传多样性进行分析,同时证明弯孢病菌存在丰富的遗传变异,且与地理来源无关。【本研究切入点】已有研究表明,新月弯孢的遗传多样性与地理来源无关,而这种现象是由何种因素所导致未见研究报道,且年度间不同地理来源的病菌遗传多样性和亲缘关系尚未明确。因此,本研究以黄淮海地区3年的弯孢病菌为研究对象,采用稳定性和多态性较好的ISSR分子标记技术,对弯孢病菌年度间不同地理种群进行遗传多样性分析,利用遗传多样性信息对病害发生的特征进行解析。【拟解决的关键问题】对采集于2013、2016和2017年黄淮海地区的玉米弯孢叶斑病菌,采用ISSR分子标记技术对不同省份和不同年份间的病菌进行遗传多样性分析,揭示黄淮海地区玉米弯孢叶斑病的流行规律,为病害防控及相关研究提供理论依据。1 材料与方法
1.1 材料
于2013、2016和2017年9月中下旬,采集黄淮海地区(河南、河北、山东、安徽、江苏)夏玉米弯孢叶斑病病叶,记录采样地点、时间等基本信息,放于标本夹中吸除水分后,干燥条件下保存于信封袋中,采用常规病害组织分离法对样品进行病原菌分离及单孢纯化[17,18],分离获得的所有菌株进行形态学鉴定和分子鉴定(ITS和EF-1α 序列分析),得到175株新月弯孢病菌(表1)。室内试验于2018年3月在河北省农林科学院植物保护研究所完成。Table 1
表1
表1供试菌株
Table 1
| 采集年份 Year | 采集地点 Collection site | 菌株编号 Strain number | |||
|---|---|---|---|---|---|
| 2013 | 安徽 Anhui | 宿州Suzhou | 萧县马井镇Majing, Xiaoxian | AH1301-1, AH1301-2 | |
| 灵璧县灵城镇Lingcheng, Lingbi | AH1301-6—AH1301-9 | ||||
| 宿县黄口镇Huangkou, Suxian | AH1301-10 | ||||
| 淮北Huaibei | 濉溪县孙疃镇Suntuan, Suixi | AH1301-3—AH1301-5 | |||
| 亳州Bozhou | 蒙城县许疃镇Xutuan, Mengcheng | AH1302-1—AH1302-3 | |||
| 蒙城县东土镇Dongtu, Mengcheng | AH1302-4 | ||||
| 蒙城县板桥集镇Banqiaoji, Mengcheng | AH1302-5, AH1302-6 | ||||
| 蒙城县王集乡Wangji, Mengcheng | AH1302-7—AH1302-9 | ||||
| 利辛县巩店镇Gongdian, Lixin | AH1302-10, AH1302-11 | ||||
| 利辛县汝集镇Ruji, Lixin | AH1302-12—AH1302-15 | ||||
| 河南 He’nan | 驻马店Zhumadian | 确山县朱古洞乡Zhugudong, Queshan | HN1301-1—HN1301-4 | ||
| 确山县三里河乡Sanlihe, Queshan | HN1301-5 | ||||
| 泌阳县盘古乡Pangu, Miyang | HN1301-6—HN1301-9 | ||||
| 泌阳县高店乡Gaodian, Miyang | HN1301-10 | ||||
| 遂平县车站镇Chezhan, Suiping | HN1301-11—HN1301-16 | ||||
| 遂平县和兴乡Hexing, Suiping | HN1301-17—HN1301-19 | ||||
| 周口Zhoukou | 商水县姚集乡Yaoji, Shangshui | HN1302-1 | |||
| 商水县城关镇Chengguan, Shangshui | HN1302-2 | ||||
| 沈丘县白集镇Baiji, Shenqiu | HN1302-3—HN1302-5 | ||||
| 沈丘县北校乡Beixiao, Shenqiu | HN1302-6—HN1302-8 | ||||
| 平顶山Pingdingshan | 叶县龚店乡Gongdian, Yexian | HN1303-1, HN1303-2 | |||
| 叶县叶邑镇Yeyi, Yexian | HN1303-3, HN1303-4 | ||||
| 叶县田庄乡Tianzhuang, Yexian | HN1303-5, HN1303-6 | ||||
| 许昌Xuchang | 许昌县尚积镇Shangji, Xuchang | HN1304-1, HN1304-2 | |||
| 许昌县蒋李集镇Jiangliji, Xuchang | HN1304-3, HN1304-4 | ||||
| 开封Kaifeng | 兰考县城关镇Chengguan, Lankao | HN1305-1, HN1305-2 | |||
| 商丘Shangqiu | 拓城县胡囊镇Hu’nang, Tuocheng | HN1306-1, HN1306-2 | |||
| 2016 | 安徽Anhui | 宿州Suzhou | 埇桥区Yongqiao | AH1601-1 | |
| 河南 He’nan | 商丘Shangqiu | 夏邑县曹集乡Caoji, Xiayi | HN1606-1 | ||
| 周口Zhoukou | 太康县张集镇Zhangji, Taikang | HN1602-1, H1N602-2 | |||
| 漯河Luohe | 舞阳县孟寨镇Mengzhai, Wuyang | HN1607-1, HN1607-2 | |||
| 开封Kaifeng | 杞县李庄村Lizhuang, Qixian | HN1605-1, HN1605-2 | |||
| 兰考县城关乡Chengguan, Lankao | HN1605-3 | ||||
| 郑州Zhengzhou | 荥阳县高村乡Gaocun, Xingyang | HN1608-1, HN1608-2 | |||
| 巩义市河洛镇Heluo, Gongyi | HN1608-3, HN1608-4 | ||||
| 焦作Jiaozuo | 沁阳市崇义镇Chongyi, Qinyang | HN1609-1 | |||
| 博爱县清化镇Qinghua, Boai | HN1609-2, HN1609-3 | ||||
| 修武县郇封镇千仓村Qiancang, Xunfeng, Xiuwu | HN1609-4, HN1609-5 | ||||
| 武陟县宁郭镇张庄村Zhangzhuang, Ningguo, Wuzhi | HN1609-6 | ||||
| 采集年份 Year | 采集地点 Collection site | 菌株编号 Strain number | |||
| 2017 | 安徽 Anhui | 亳州Bozhou | 利辛县垄店镇Gongdian, Lixin | AH1702-1 | |
| 蒙城县王集乡Wangji, Mengcheng | AH1702-2 | ||||
| 蒙城县板桥集镇Banqiaoji, Mengcheng | AH1702-3 | ||||
| 宿州Suzhou | 灵璧县灵城镇Lingcheng, Lingbi | AH1701-1, AH1701-2 | |||
| 埇桥区大店镇Dadian, Yongqiao | AH1701-3 | ||||
| 埇桥区Yongqiao | AH1701-4 | ||||
| 淮北Huaibei | 濉溪县刘桥镇Liuqiao, Suixi | AH1703-2 | |||
| 相山区Xiangshan | AH1703-3 | ||||
| 濉溪县南平镇Nanping, Suixi | AH1703-1 | ||||
| 河北 Hebei | 石家庄Shijiazhuang | 新乐市马头铺镇Matoupu, Xinle | HB1701-1 | ||
| 赵县赵州镇Zhaozhou, Zhaoxian | HB1701-2 | ||||
| 藁城区Gaocheng | HB1701-3, HB1701-5, HB1701-6 | ||||
| 灵寿县北洼乡Beiwa, Lingshou | HB1701-4 | ||||
| 保定Baoding | 望都韩庄镇Hanzhuang,Wangdu | HB1702-1 | |||
| 蠡县南庄镇Nanzhuang, Lixian | HB1702-2 | ||||
| 雄县双堂乡Shuangtang, Xiongxian | HB1702-3 | ||||
| 蠡县蠡吴镇Liwu, Lixian | HB1702-4 | ||||
| 高阳县高阳镇Gaoyang, Gaoyang | HB1702-5 | ||||
| 满城大册营镇Daceying, Mancheng | HB1702-6 | ||||
| 易县高村镇Gaocun, Yixian | HB1702-7 | ||||
| 涞源王安镇Wang’an, Laiyuan | HB1702-8 | ||||
| 易县尉都乡Weidu, Yixian | HB1702-9 | ||||
| 涞源县胡家庄乡Hujiazhuang, Laiyuan | HB1702-10 | ||||
| 涿州市松林店镇Songlindian, Zhuozhou | HB1702-11 | ||||
| 定兴县定兴镇Dingxing, Dingxing | HB1702-12 | ||||
| 徐水区Xushui | HB1702-13 | ||||
| 邯郸Handan | 邯郸县丛台区Congtai, Handan | HB1703-1 | |||
| 峰峰县新坡镇Xinpo, Fengfeng | HB1703-2 | ||||
| 大城县大尚屯镇Dashangtun, Dacheng | HB1706-3 | ||||
| 大城县广安镇Guang’an, Dacheng | HB1706-4 | ||||
| 衡水Hengshui | 武邑县肖桥头镇Xiaoqiaotou, Wuyi | HB1704-3, HB1704-4 | |||
| 故城县故城镇Gucheng, Gucheng | HB1704-1 | ||||
| 深州市Shenzhou | HB1704-2 | ||||
| 沧州Cangzhou | 东光县找王镇Zhaowang, Dongguang | HB1705-1 | |||
| 青县新兴镇Xinxing, Qingxian | HB1705-2 | ||||
| 沧县崔尔庄镇Cuierzhuang, Cangxian | HB1705-3 | ||||
| 河间市时村乡Shicun, Hejian | HB1705-4 | ||||
| 任丘市麻家坞镇Majiawu, Renqiu | HB1705-5 | ||||
| 青县曹寺镇Caosi, Qingxian | HB1705-6 | ||||
| 献县淮镇Huaizhen, Xianxian | HB1705-7 | ||||
| 廊坊Langfang | 固安县马庄镇Mazhuang, Gu’an | HB1706-1 | |||
| 固安县Gu’an | HB1706-2 | ||||
| 固安县牛驼镇Niutuo, Gu’an | HB1706-5 | ||||
| 采集年份 Year | 采集地点 Collection site | 菌株编号 Strain number | |||
| 唐山Tangshan | 玉田县杨家套乡Yangjiatao, Yutian | HB1707-1 | |||
| 昌黎县葛条港乡Getiaogang, Changli | HB1707-2 | ||||
| 玉田县石臼窝镇Shijiuwo, Yutian | HB1707-3 | ||||
| 滦州市滦州镇Luanzhou, Luanzhou | HB1707-6 | ||||
| 滦州市Luanzhou | HB1707-7 | ||||
| 遵化市团瓢庄乡Tuanpiaozhuang, Zunhua | HB1707-8 | ||||
| 迁安市Qian’an | 五重安乡Wuchongan | HB1707-4 | |||
| 赵新庄Zhaoxinzhuang | HB1707-5 | ||||
| 承德Chengde | 承德县下板城镇Xiabancheng, Chengde | HB1709-1 | |||
| 秦皇岛Qinhuangdao | 卢龙县Lulong | HB1708-1 | |||
| 抚宁区Funing | HB1708-2 | ||||
| 河南 He’nan | 安阳Anyang | 安阳县崔家桥镇Cuijiaqiao, Anyang | HN1710-1 | ||
| 鹤壁Hebi | 淇县西岗镇Xigang, Qixian | HN1711-1 | |||
| 新乡Xinxiang | 辉县胡桥乡Huqiao, Huixian | HN1712-1 | |||
| 新乡县Xinxiang | HN1712-2 | ||||
| 漯河Luohe | 源汇区Yuanhui | HN1707-1 | |||
| 驻马店Zhumadian | 西平县二郎镇Erlang, Xiping | HN1701-1 | |||
| 周口Zhoukou | 西华县田口乡Tiankou, Xihua | HN1702-1 | |||
| 平顶山Pingdingshan | 宝丰县杨庄镇Yangzhuang, Baofeng | HN1703-1 | |||
| 商丘Shangqiu | 柘城县Zhecheng | HN1706-1 | |||
| 江苏 Jiangsu | 徐州Xuzhou | 铜山区大彭镇Dapeng, Tongshan | JS1701-1, JS1701-2 | ||
| 沛县栖山镇Qishan, Peixian | JS1701-3 | ||||
| 沛县鹿楼镇Lulou, Peixian | JS1701-4 | ||||
| 沛县朱寨镇Zhuzhai, Peixian | JS1701-5 | ||||
| 山东 Shandong | 滨州Binzhou | 阳信县劳店镇Laodian, Yangxin | SD1701-1 | ||
| 德州Dezhou | 乐陵县Leling | SD1702-1 | |||
| 平原县Pingyuan | SD1702-2 | ||||
| 德城区Decheng | SD1702-3 | ||||
| 济宁Jining | 微山县Weishan | SD1704-1 | |||
| 曲阜市石门山镇Shimenshan, Qufu | SD1704-2 | ||||
| 汶上县苑庄镇Yuanzhuang, Wenshang | SD1704-3 | ||||
| 嘉祥县瞳里镇Tongli, Jiaxiang | SD1704-4 | ||||
| 任城区Rencheng | SD1704-5 | ||||
| 枣庄Zaozhuang | 滕州市西岗镇Xigang,Tengzhou | SD1703-1 | |||
| 峄城区峨山镇Eshan, Yicheng | SD1703-2 | ||||
| 临沂Linyi | 兰陵县金陵镇Jinling, Lanling | SD1705-1 | |||
| 兰山区汪沟镇Wanggou, Lanshan | SD1705-2 | ||||
| 莱芜Laiwu | 莱芜区杨庄镇Yangzhuang, Laiwu | SD1706-1 | |||
| 泰安Taian | 东平县彭集镇Pengji, Dongping | SD1707-1 | |||
| 菏泽Heze | 鄄城县红船镇Hongchuan, Juancheng | SD1708-1 | |||
| 聊城Liaocheng | 聊城市东昌府区Dongchangfu, Liaocheng | SD1709-1 | |||
| 潍坊Weifang | 临朐县Linqu | SD1710-1 | |||
新窗口打开|下载CSV
1.2 模板DNA的制备
将分离到的175个菌株分别接种到PDA平板培养基上,28℃培养5 d,用打孔器(直径为1 cm)取3个菌饼放入PD培养基中,在28℃,180 r/min条件下培养5 d,过滤收集菌丝。收集的菌丝经冷冻干燥后,采用真菌基因组提取试剂盒(北京艾德莱生物科技有限公司)提取DNA。采用ND2000紫外分光光度(Thermo,USA)测定DNA的浓度,统一稀释至50 ng·μL-1,贮存于-20℃保存,备用。1.3 ISSR分析
1.3.1 ISSR引物筛选 随机选取2013和2016年各1个菌株和2017年的2个菌株的DNA作为模板,对加拿大哥伦比亚大学(University of British Columbia,UBC)60条引物进行筛选,选取扩增条带清晰、多态性高、重复性好的13条引物用于本试验175个DNA样品的扩增。1.3.2 ISSR-PCR体系及程序 PCR反应体系:2×Es TaqMasterMix 10 μL,引物(10 μmol·L-1)1.5 μL,DNA模板1 μL,加ddH2O补足至20 μL。采用的13条引物见表2。PCR反应程序:95℃预变性5 min,94℃变性30 s,退火(视不同引物而定)45 s,72℃延伸90 s,35个循环,最后72℃延伸10 min。
Table 2
表2
表2ISSR引物扩增
Table 2
| 引物 Primer | 序列 Sequence (5′-3′) | 退火温度 Tm (℃) | 扩增条带数 Number of amplified bands | 多态性条带 Number of polymorphic bands | 多态性比率 Polymorphism ratio (%) |
|---|---|---|---|---|---|
| 808 | (AG)8C | 49 | 10 | 10 | 100 |
| 810 | (GA)8T | 45 | 7 | 7 | 100 |
| 823 | (TC)8C | 50 | 6 | 6 | 100 |
| 827 | (AC)8G | 52 | 7 | 7 | 100 |
| 835 | (AG)8YC | 53 | 10 | 10 | 100 |
| 841 | (GA)8YC | 50 | 11 | 11 | 100 |
| 842 | (GA)8YG | 50 | 10 | 10 | 100 |
| 847 | (CA)8RC | 54 | 6 | 6 | 100 |
| 855 | (AC)8YT | 46 | 7 | 7 | 100 |
| 873 | (GACA)4 | 49 | 7 | 7 | 100 |
| 880 | (GGAGA)3 | 46 | 7 | 7 | 100 |
| 888 | BDB(CA)7 | 48 | 9 | 9 | 100 |
| 891 | HVH(TG)7 | 47 | 8 | 8 | 100 |
| 总计Total | 105 | 105 | 100 |
新窗口打开|下载CSV
PCR扩增产物在1.8%琼脂糖凝胶上以电压100 V,电流100 mA,进行电泳50 min,随后使用GelDoc XR+凝胶成像系统进行拍照。
1.3.3 数据分析 对ISSR电泳谱带进行统计,分别用1和0表示条带的有无,缺失条带记为“9”或“.”。使用Popgen32生物软件进行数据运算,计算群体内和群体间的Shannon’s指数和Nei’s指数、等位基因数、有效等位基因数、群体间的遗传距离和遗传相似性[19,20,21];利用NTsys2.10e软件,采用clustering模块中的SAHN进行UPGMA聚类分析,构建供试菌株的系统聚类图,并进行基于遗传相似系数的主坐标分析(principal coordinates analysis,PCA)。
2 结果
2.1 ISSR-PCR扩增
从60个引物中筛选出13条扩增条带清晰、多态性高、重复性好的引物(表2),对175个采集自不同年度和不同地区的菌株进行ISSR-PCR扩增(图1)。这些引物都具有较好的扩增多态性,共获得105个条带,多分布在200—2 000 bp,其中多态性条带105个,占100%。不同引物扩增出的条带数目不同,多为6—11条,平均每个引物产生8个多态性条带。其中841号引物扩增出的条带数最多(11条),而引物823和847扩增出的条带数最少(6条),13条引物的多态性条带比例均为100%(表2)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图14个引物对部分新月弯孢菌株基因组DNA的ISSR-PCR扩增结果
Fig. 1Amplification results of genome DNA of some C. lunata strains with four primers
2.2 新月弯孢遗传多态性分析
将分离到的175株新月弯孢按省份分为不同地理来源的5个种群,其遗传多样性分析如表3所示。在群体平均水平上,多态位点为105个,多态性位点平均百分率为100%,观察等位基因数(Na)为2.0000,有效等位基因数(Ne)为1.6761,Nei’s基因多样性指数(H)为0.3867,Shannon’s信息指数(I)为0.5682,表明新月弯孢群体存在丰富的遗传变异。Table 3
表3
表3新月弯孢不同地理种群的遗传多样性水平
Table 3
| 多态性位点 Number of polymorphic loci | 多态性比率 Polymorphism ratio (%) | 观察等位基因数 Observed number of alleles (NA) | 有效等位基因 Effective number of alleles (NE) | 基因多样性指数Nei’s gene diversity (H) | Shannon’s信息指数 Shannon’s information index (I) | |
|---|---|---|---|---|---|---|
| 安徽AH | 103 | 98.10 | 1.9810 | 1.6344 | 0.3665 | 0.5428 |
| 河南HN | 105 | 100.00 | 2.0000 | 1.6309 | 0.3662 | 0.5438 |
| 河北HB | 96 | 91.43 | 1.9143 | 1.5855 | 0.3415 | 0.5072 |
| 江苏JS | 58 | 55.24 | 1.5524 | 1.3904 | 0.2170 | 0.3171 |
| 山东SD | 90 | 85.71 | 1.8571 | 1.5355 | 0.3109 | 0.4623 |
| 群体水平 Population level | 105 | 100.00 | 2.0000 | 1.6761 | 0.3867 | 0.5682 |
新窗口打开|下载CSV
不同地理来源的种群间,H值在0.2170—0.3665,I值在0.3171—0.5438,表明新月弯孢的遗传多样性在不同地理种群间存在一定差异。总体而言,河南(0.3662)和安徽(0.3665)种群遗传多样性最高,山东(0.3109)和河北(0.3415)种群次之,江苏(0.2170)种群较低。
2.3 新月弯孢遗传相似性分析
为进一步分析新月弯孢各地理种群间的遗传分化程度,将分离到的175株新月弯孢菌株按不同年度和省份分组,计算了Nei’s遗传相似性GS和遗传距离GD(表4),以及群体间的基因分化系数Gst值。不同年份各地理种群间的遗传相似系数为0.6316—0.9869,遗传距离为0.0132—0.4594,表明不同年份地理种群间的分化程度不同,遗传距离不一致。2013年河南与安徽的菌株群体遗传相似系数最高(GS=0.9869),遗传距离最近(GD=0.0132),2017年江苏与安徽的菌株群体,山东与河南、河北的菌株群体遗传相似系数较高,表明同一年份不同地理种群间相关性较高;江苏2017年菌株群体与安徽2016年菌株群体遗传相似系数最小(GS=0.6316),遗传距离最远(GD=0.4594),2016年安徽与2013年安徽、河南菌株群体遗传相似系数较小,表明相关性较低。Table 4
表4
表4新月弯孢不同地理种群间的遗传相似性和遗传距离
Table 4
| 种群Group | 安徽AH13 | 河南HN13 | 安徽AH16 | 河南HN16 | 安徽AH17 | 河北HB17 | 河南HN17 | 江苏JS17 | 山东SD17 |
|---|---|---|---|---|---|---|---|---|---|
| 安徽AH13 | 0.9869 | 0.6550 | 0.8571 | 0.7644 | 0.8767 | 0.8621 | 0.7855 | 0.8652 | |
| 河南HN13 | 0.0132 | 0.6467 | 0.8559 | 0.7682 | 0.8829 | 0.8623 | 0.7854 | 0.8669 | |
| 安徽AH16 | 0.4232 | 0.4358 | 0.7510 | 0.6771 | 0.6893 | 0.6751 | 0.6316 | 0.6953 | |
| 河南HN16 | 0.1542 | 0.1557 | 0.2863 | 0.7542 | 0.8292 | 0.7847 | 0.7424 | 0.8173 | |
| 安徽AH17 | 0.2687 | 0.2637 | 0.3899 | 0.2821 | 0.8698 | 0.9014 | 0.9435 | 0.8947 | |
| 河北HB17 | 0.1316 | 0.1246 | 0.3720 | 0.1873 | 0.1395 | 0.8924 | 0.8809 | 0.9491 | |
| 河南HN17 | 0.1483 | 0.1481 | 0.3929 | 0.2424 | 0.1038 | 0.1139 | 0.8897 | 0.9221 | |
| 江苏JS17 | 0.2415 | 0.2415 | 0.4594 | 0.2978 | 0.0582 | 0.1269 | 0.1168 | 0.9014 | |
| 山东SD17 | 0.1448 | 0.1429 | 0.3635 | 0.2018 | 0.1112 | 0.0522 | 0.0811 | 0.1038 |
新窗口打开|下载CSV
2013和2016年各省份之间的遗传相似系数分别为0.9869和0.7510,遗传距离分别为0.0132和0.2863;2017年各省份之间的遗传相似系数为0.8698—0.9491,遗传距离为0.0522—0.1395;表明相同年份不同地理来源的菌株具有较小的遗传差异。
安徽省2013与2016、2017年的菌群遗传相似系数分别为0.6550和0.7644,2016与2017年为0.6771;河南省2013与2016、2017年的菌群遗传相似系数分别为0.8559和0.8623,2016与2017年为0.7847,表明年度间菌群的亲缘关系相对较远。
2013、2016和2017年菌株群体间的Gst值为0.1569,表明不同年度间遗传变异占种群总遗传变异的15.69%,大部分遗传变异(84.31%)发生在种群内。菌株群体间的基因流Nm为2.6869(<4),表明不同年度间种群有一定的遗传差异;5个地理种群总基因多样性Ht为0.3857,各个种群内基因多样性Hs为0.2582,群体间的基因多样性Dst为0.1275,表明相同年份和省份来源的菌株具有较近的亲缘关系,各地区新月弯孢的基因流Nm为2.9420,表明不同地区间存在一定的基因交流。
2.4 新月弯孢ISSR聚类分析及主坐标分析
利用NTsys2.10e软件对175个菌株进行聚类分析,结果如图2所示,菌株间具有丰富的遗传多样性,相似系数在0.51—0.93。在相似系数0.59水平上,所有的菌株被分为2个大类群(Group 1和Group 2)5个亚群(subgroup)。Group 1包含173个菌株;Group 2包括河南(2013和2017年各1株)2个菌株。Group 1又分为5个亚群,subgroupⅠ为2013年采集自河南和安徽省的63个菌株和2016年采集自河南开封市的1个菌株;subgroupⅡ为2016年采集自河南和安徽的18个菌株;subgroupⅢ为2013年采集自河南许昌市的1个菌株;subgroupⅣ为2017年采集自河南、河北、山东、江苏和安徽的89个菌株;subgroupⅤ为2013年采集自河南周口市的1个菌株。地理种群之间新月弯孢表现出地域相邻,其遗传关系较近,遗传相似系数较高的特点,如河南与安徽,河北与山东,表明不同地域的菌株来源可能相近;同一地区的菌株在年度间表现出一定遗传距离,遗传相似系数较低,表明年度间的菌株初侵染来源可能存在不同。图2
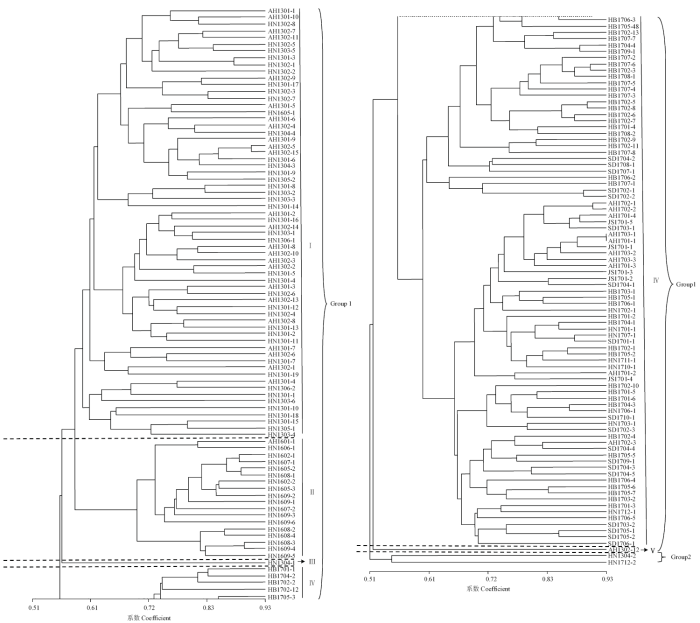 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2新月弯孢菌株的UPGMA聚类图
Fig. 2Dendrogram obtained by UPGMA clustering for C. lunata
利用NTSY2.10软件对175份新月弯孢菌株进行基于遗传距离的主坐标分析(PCA),前3个主坐标显示遗传变异分别为11.81%、6.79%、5.79%。三维散点图(图3)显示,175个菌株形成了3个较为独立的分布区,2013、2016、2017 3个年份分别聚合为一个独立的分布区,这与聚类分析结果基本一致。
图3
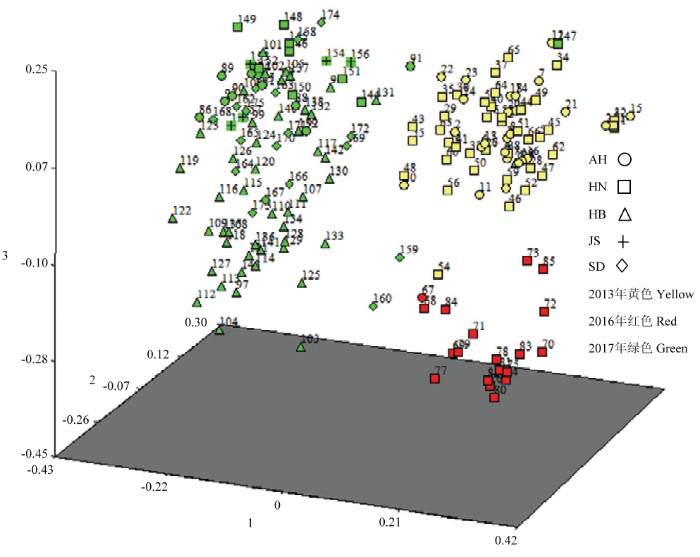 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3新月弯孢菌株主坐标分析的三维散点图
1—175:新月弯孢菌株编号坐标轴1、2、3分别代表前3个主成分对遗传变异的贡献率
Fig. 3Three-dimensional scatter plot of principal coordinate analysis of C. lunata
Number of C. lunata strains;Coordinate axis 1, 2, 3 represent the contribution rate of the first three principal components to genetic variation, respectively
3 讨论
3.1 新月弯孢的遗传多样性
基于PCR的分子标记技术广泛应用于植物、病原菌等物种遗传多样性和亲缘关系的研究,马红霞等[22]利用ISSR分子标记技术证明禾谷镰孢(Fusarium graminearum )复合种菌株群体内存在丰富的遗传变异,且遗传多样性与地理来源有关;郭云燕、鄢洪海等[23,24]利用ISSR分子标记技术分析了玉米南方锈病病原菌(Puccinia polysora )的遗传多样性,并对其初侵染源进行了探讨。本研究通过对2013、2016和2017年分离自黄淮海地区5省的175个新月弯孢的ISSR片段多样性进行分析,得到105个多态性条带,多态性比率为100%,表明新月弯孢具有丰富的遗传变异,这一结果与王晓东等[16]的研究结果一致。在群体平均水平上,Nei’s基因多样性指数(H)为0.3867,Shannon’s信息指数(I)为0.5682,表明新月弯孢群体存在丰富的遗传多样性。5个地理种群的H值和I值的结果趋势一致,河南和安徽省群体具有丰富的遗传多样性水平,江苏省群体遗传多样性较低,可能与江苏种群菌株数目较少有关。UPGMA聚类分析显示,在相似系数0.59水平上,所有的菌株被分为2个大群5个亚群,2013、2016和2017年的菌株明显聚为3支,分别分布于subgroupⅠ、subgroupⅡ、subgroupⅣ。但2013年有2个菌株分别分布于subgroupⅢ和subgroupⅤ,2016年有1个菌株分布于subgroupⅠ,基因流分析结果也表明,不同年度间种群存在一定的基因交流。遗传相似性分析表明,年度间相同地理来源的菌群亲缘关系较远,相同年份不同地理来源菌群亲缘关系较近。
3.2 玉米弯孢叶斑病的初侵染源分析
玉米弯孢叶斑病病菌主要通过气流进行远距离传播,以菌丝体或分生孢子随病残体越冬,有关越冬场所和越冬条件等也有很多相关研究报道。张定法[25]通过对病残体上病原菌越冬存活检测,表明在河南新乡地区病菌的菌丝体在土壤10 cm处越冬率较低;暴增海等[26]对冀东地区玉米弯孢叶斑病菌的初侵染源进行研究,发现该病原菌可随病残体在地表越冬,但存在于10—30 cm土层中的病残体不能越冬;戴法超等[27]研究指出,干燥是病残体中病菌存活和越冬的首要条件,在潮湿条件下病残体容易腐烂,因此埋于土层的病残体未分离到弯孢菌。近年来,由于黄淮海地区耕作制度的改变,多地均采取玉米秸秆深翻还田,而后进行冬小麦种植的耕作措施。弯孢病菌随含水量较高的病残体被深翻埋于土层之中,而冬小麦的种植需进行2—3次灌溉,因此,黄淮海地区的栽培模式未能提供病原菌越冬的适宜条件,本团队多年的田间调查表明该病的发生具有间歇性,且病斑从植株上部向中下部蔓延,具有病原菌的初侵染来源为外来菌源的特点,但并无相关研究证明。本研究结果表明,年度间相同地理来源的菌群亲缘关系较远,相同年份不同地理来源菌群亲缘关系较近,且不同年度间种群存在一定的基因交流。因此推测,黄淮海地区引起玉米弯孢叶斑病的新月弯孢菌株不是以本地菌源为主,同时存在少量存活于田间地表病残体上的菌株可作为本地菌源在翌年进行侵染。徐辉[28]研究发现,新月弯孢可引起水稻弯孢叶斑病,且在安徽地区有蔓延的趋势;在长江以南地区的水稻田,新月弯孢菌株还可以引起水稻叶鞘黑斑病、穗腐病、穗枯病等[29,30,31,32];也可以引起几种禾草草坪草的叶部病害[33,34,35,36,37]。因此南方地区冬季未处理的水稻带菌稻秸和多年生草坪草等可能作为弯孢病菌的越冬场所,第二年病菌随东南气流进入黄淮海玉米生产区,为该地区玉米弯孢叶斑病的发生提供初侵染菌来源;另一种可能是病菌随台风来自东南亚玉米种植区。因此,下一步研究应采集来自南方不同地区不同作物上的新月弯孢菌株进行验证。
3.3 影响病菌变异及群体遗传多样性的因子
遗传多样性及聚类分析结果表明,年度间菌群遗传差异较大,但遗传多样性与地理种群无关。推测可能是由于不是以本地菌源为主所导致;其次病原菌可能随气流或台风从不同菌源地远距离传入黄淮海玉米种植区。同时病原菌变异较快的原因可能是由环境等外界因素所导致。寄主品种和抗病性差异、栽培模式以及不同地域对病原菌的遗传基因型、毒力表型具有显著的影响[38,39]。刘二明对稻瘟病菌(Magnaporthe grisea )遗传多样性研究发现,水稻种植品种越多的地区,该地区的群体遗传多样性越丰富[40];对于小麦锈菌(Puccinia triticina )和麦类白粉菌(Blumeria graminis )等专性寄生菌,由于周年循环过程需与寄主相互作用,因此寄主与病原菌之间也可产生互作、变异、适应性改变等[41,42]。黄淮海地区玉米种植品种繁多,且本研究采取田间随机采样模式,因此寄主遗传背景的差异可能会对弯孢病菌的基因型有一定影响。病原物、寄主与环境条件相互作用对病原菌的基因型也产生一定影响。病原菌菌丝体普遍为多核,且每个细胞核均能发育成为单个菌丝体[43,44],这种现象有利于病菌遗传差异的产生,且菌群结构易受环境等外界因素的影响[45],因此环境因素也是造成弯孢病菌年度间差异较大的原因之一。同时本研究中年度间菌群差异明显,可能是由于年度间菌群不是以本地菌源为主所致,以及随气流或台风在高空漂移过程中,来自空中的各种辐射也是导致遗传差异的一个重要因素[23,41]。
4 结论
黄淮海地区玉米弯孢叶斑病菌(新月弯孢)群体内具有丰富的遗传多样性,即存在种内遗传分化现象。聚类分析表明,地域相邻的新月弯孢遗传关系较近,遗传相似系数较高;同一地区的菌株在年度间表现出一定遗传距离,遗传相似系数较低。遗传多样性分析结果表明,引起玉米弯孢叶斑病的新月弯孢菌株不是以本地菌源为主,但也存在少量存活于地表病残体上的菌株可作为翌年的初侵染源。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1007/BF02007173URL [本文引用: 1]

正 1944年在北京郊区夏玉米发生一种异于小斑病和园斑病的叶斑病,严重发生的玉米杂交种有京早10号、西玉3号等。顺义县一乡镇种植西玉3号6000余亩,全部发病,造成减产。9月上旬,田间观察该病症状,病斑卵园形或椭园形,大小为1~4mm,中央乳白色,外围有褐色环并带有褪色晕圈。严重感病的品种全株叶片密布病斑,病斑偶有联合,引致全叶枯死。这种病害症状极易与眼斑病的症状混淆。 用采自田间西玉3号植株病叶进行分离接种,试验在中国农科院引种隔离检疫基地温室进行。第一种接种试验
DOI:10.1007/BF02007173URL [本文引用: 1]
正 1944年在北京郊区夏玉米发生一种异于小斑病和园斑病的叶斑病,严重发生的玉米杂交种有京早10号、西玉3号等。顺义县一乡镇种植西玉3号6000余亩,全部发病,造成减产。9月上旬,田间观察该病症状,病斑卵园形或椭园形,大小为1~4mm,中央乳白色,外围有褐色环并带有褪色晕圈。严重感病的品种全株叶片密布病斑,病斑偶有联合,引致全叶枯死。这种病害症状极易与眼斑病的症状混淆。 用采自田间西玉3号植株病叶进行分离接种,试验在中国农科院引种隔离检疫基地温室进行。第一种接种试验
URL [本文引用: 1]
经作者分离接种鉴定,确认近年北京郊区和河北省的一些玉米产区发生的叶斑病为玉米弯孢菌叶斑病,病原菌是Curvu larialunata。发生此病玉米可减产20%~60%,且有逐年加重的趋势,经人工接种鉴定不同品种(系)抗性不同Mo17,黄早四,丹340,478,5003和E28等骨干自交系均为感病类型(3~4级)。西玉3号,京早10号不同生育期抗性不同,以3~4叶期为抗病类型(0.5级)7~8叶期,
URL [本文引用: 1]
经作者分离接种鉴定,确认近年北京郊区和河北省的一些玉米产区发生的叶斑病为玉米弯孢菌叶斑病,病原菌是Curvu larialunata。发生此病玉米可减产20%~60%,且有逐年加重的趋势,经人工接种鉴定不同品种(系)抗性不同Mo17,黄早四,丹340,478,5003和E28等骨干自交系均为感病类型(3~4级)。西玉3号,京早10号不同生育期抗性不同,以3~4叶期为抗病类型(0.5级)7~8叶期,
DOI:10.3969/j.issn.1672-6820.2003.04.023URL [本文引用: 1]
综合作者研究成果和国内外研究资料,阐述了玉米弯孢菌叶斑病的病原、发生症状、为害损失、影响发生的主要因素和防治措施等.
DOI:10.3969/j.issn.1672-6820.2003.04.023URL [本文引用: 1]
综合作者研究成果和国内外研究资料,阐述了玉米弯孢菌叶斑病的病原、发生症状、为害损失、影响发生的主要因素和防治措施等.
DOI:10.3969/j.issn.1005-0906.2004.02.030URL [本文引用: 1]
玉米弯孢叶斑病[Curvularia lunata(Wakker)Boed]是我国近几年玉米上发生的重要病害.其危害在一些地区已经超过大小斑病,成为玉米主要病害.从目前国内外的研究现状、症状、病原、发生规律、危害损失、抗病性以及综合防治等方面对该病进行了综述,同时讨论了今后玉米弯孢叶斑病的研究方向.
DOI:10.3969/j.issn.1005-0906.2004.02.030URL [本文引用: 1]
玉米弯孢叶斑病[Curvularia lunata(Wakker)Boed]是我国近几年玉米上发生的重要病害.其危害在一些地区已经超过大小斑病,成为玉米主要病害.从目前国内外的研究现状、症状、病原、发生规律、危害损失、抗病性以及综合防治等方面对该病进行了综述,同时讨论了今后玉米弯孢叶斑病的研究方向.
.
DOI:10.1684/agr.2013.0625URL [本文引用: 1]
Molecular characterization of 19 Jatropha accessions that included 15 accessions ofJ.curcas and 4 different species was carried out using 3 different markers systems.Highest polymorphism (96.67%) was recorded by RAPD followed by DAMD (91.02%)and ISSR (90%). Polymorphism Information Content (PIC) was higher for DAMD(0.873) and almost equal for RAPD (0.863) and ISSR (0.862) markers, whereas ResolvingPower (Rp) was found to be higher for RAPD as compared to the other two markersystems. Marker Index (MI) values varied greatly with highest (19.07) in RAPD. Shannonindex (i), observed number of alleles (na), effective number of alleles (ne) and Nei genetic diversity (h) values were found to be significantly higher for ISSR as compared toRAPD and DAMD markers. Thus, all the markers proved to be equally efficient fordiversity studies in Jatropha. Several alleles in all the markers indicated J. gossypiifolia asone of the parents of J. tanjorensis. Dendrograms and PCA plots generated based onRAPD showed three major clusters with J. integerrima and J. podagrica falling in group I,fifteen J. curcas accessions in group II, and J. gossypiifolia as an outlier in group III.DAMD markers also showed similar clustering pattern whereas ISSR showed last clusterof J. gossypiifolia and J. tanjorensis. These results may provide a future base forconservation and characterization of available Jatropha genetic resources.Keywords: Genetic diversity, Jatropha, Molecular markers, Polymorphism.
DOI:10.3969/j.issn.0517-6611.2010.36.060URL [本文引用: 1]
ISSR分子标记是PCR基础 上发展起来的一种新技术,其基本原理是在SSR的5′或3′端加锚1~4个嘌呤或嘧啶碱基,然后以此为引物对两侧具有反向排列的一段基因组序列进行扩增。 阐述了ISSR的原理和特点,并论述了ISSR标记技术目前在植物病原菌分类、亲缘关系和遗传多样性等方面的研究及应用。
DOI:10.3969/j.issn.0517-6611.2010.36.060URL [本文引用: 1]
ISSR分子标记是PCR基础 上发展起来的一种新技术,其基本原理是在SSR的5′或3′端加锚1~4个嘌呤或嘧啶碱基,然后以此为引物对两侧具有反向排列的一段基因组序列进行扩增。 阐述了ISSR的原理和特点,并论述了ISSR标记技术目前在植物病原菌分类、亲缘关系和遗传多样性等方面的研究及应用。
.
DOI:10.1016/j.rsci.2016.08.001URL [本文引用: 1]
Inter simple sequence repeat (ISSR) polymorphism was used to determine genetic diversity and phylogenetic relationships in 90 genotypes of wild and cultivated species ofOryzafrom different geographical regions of the world. In all the 17 primers used in ISSR-PCR, a total of 11 464 bands were amplified at 253 band positions/loci. The primer UBC-809 amplified the maximum bands (1 059) at 21 band positions. UBC-810 and UBC-835 amplified the minimum of 391 bands each at 7 and 14 band positions, respectively. The mean polymorphism information content ranged from 0.44 to 0.84 and resolving power ranged from 8.69 to 23.53. Un-weighted pair group method with arithmetic mean dendrogram and population structure based on the 17 primers separated all genotypes into 4 major clusters with a genetic similarity of 53% 100%. The first two clusters consisted of 30O. rufipogonaccessions each. In the third cluster,O. nivaraandO. longistaminatagrouped as one sub-cluster and all otherO. nivaraaccessions and cultivars grouped as another sub-cluster. The fourth cluster had only fiveO. rufipogonaccessions which can be a source of new genes. Four sub-populations were identified withinO. rufipogonand two sub-populations withinO. nivaraatK=7. A subset of six primers with high resolving power values were the most informative and grouped all genotypes almost similarly as the 17 primers did. Use of these six highly informative primers in ISSR-PCR is a cost effective and robust method for assessing genetic diversity in large germplasm collections of wild rice species.
DOI:10.3969/j.issn.0006-3193.2004.02.008URL [本文引用: 1]
ISSR分子标记技术是近年发展而来的一种DNA多态性分子标记,具有简单、经济、信息量大、稳定性高等特点。本文论述了ISSR的反应原理及ISSR实验过程中的技术要点,并总结了ISSR在植物品种鉴定、亲缘关系分析,群体遗传结构、遗传多样性检测,以及植物育种研究中的应用。
DOI:10.3969/j.issn.0006-3193.2004.02.008URL [本文引用: 1]
ISSR分子标记技术是近年发展而来的一种DNA多态性分子标记,具有简单、经济、信息量大、稳定性高等特点。本文论述了ISSR的反应原理及ISSR实验过程中的技术要点,并总结了ISSR在植物品种鉴定、亲缘关系分析,群体遗传结构、遗传多样性检测,以及植物育种研究中的应用。
URL [本文引用: 1]
为明确我国烟草赤星病的2种主要致病菌链格孢菌Alternaria alternata和长柄链格孢菌A.longipes的地理差异与遗传结构,采用ISSR标记对分离自9个省市的135株烟草赤星病菌进行遗传多样性分析。结果显示,通过正交优化试验建立的烟草赤星病菌ISSR-PCR最佳反应体系稳定性较好,筛选出17条多态性高且稳定的引物,共扩增出192条谱带,其中有177条具有多态性,多态率为92.19%。UPGMA聚类分析结果显示,链格孢菌和长柄链格孢菌的遗传相似性系数分别在0.67~1.00和0.66~1.00之间,遗传相似性系数为0.83时可使链格孢菌和长柄链格孢菌分别划分为5个和6个亚群,其中前者不同地理种群间表现出地理相关性,后者不同菌株随机分组。烟草赤星病菌种群的基因多态性和遗传多样性丰富,链格孢菌和长柄链格孢菌的群体间遗传分化系数分别为0.36和0.37,均存在遗传分化;群居每代迁移数分别为0.89和0.85,不同地理种群间存在基因交流;2种烟草赤星病菌的遗传分化结构表现出相似性。表明我国烟草赤星病菌中的链格孢菌和长柄链格孢菌均存在丰富的遗传多样性,且二者进化方向相似,ISSR标记能较好地揭示烟草赤星病菌种群间的亲缘关系和遗传差异性,可用于其遗传多样性分析。
URL [本文引用: 1]
为明确我国烟草赤星病的2种主要致病菌链格孢菌Alternaria alternata和长柄链格孢菌A.longipes的地理差异与遗传结构,采用ISSR标记对分离自9个省市的135株烟草赤星病菌进行遗传多样性分析。结果显示,通过正交优化试验建立的烟草赤星病菌ISSR-PCR最佳反应体系稳定性较好,筛选出17条多态性高且稳定的引物,共扩增出192条谱带,其中有177条具有多态性,多态率为92.19%。UPGMA聚类分析结果显示,链格孢菌和长柄链格孢菌的遗传相似性系数分别在0.67~1.00和0.66~1.00之间,遗传相似性系数为0.83时可使链格孢菌和长柄链格孢菌分别划分为5个和6个亚群,其中前者不同地理种群间表现出地理相关性,后者不同菌株随机分组。烟草赤星病菌种群的基因多态性和遗传多样性丰富,链格孢菌和长柄链格孢菌的群体间遗传分化系数分别为0.36和0.37,均存在遗传分化;群居每代迁移数分别为0.89和0.85,不同地理种群间存在基因交流;2种烟草赤星病菌的遗传分化结构表现出相似性。表明我国烟草赤星病菌中的链格孢菌和长柄链格孢菌均存在丰富的遗传多样性,且二者进化方向相似,ISSR标记能较好地揭示烟草赤星病菌种群间的亲缘关系和遗传差异性,可用于其遗传多样性分析。
DOI:10.13802/j.cnki.zwbhxb.2015.06.007URL [本文引用: 1]
为明确我国发生的玉米灰斑病菌地理差异及遗传结构,利用简单序列重复区间(ISSR)对玉米灰斑病菌遗传多样性进行了分析,并利用尾孢菌特异引物对分离自四川、云南、湖北、贵州等西南地区的16个玉米灰斑病菌菌株进行了分子鉴定。结果显示,通过ISSR标记筛选出10个扩增多态性好且稳定的通用引物,共扩增出81条DNA条带,均为多态性条带,扩增片段大小在200-2 000 bp之间,菌株遗传相似系数为0.19-1.00。在遗传相似系数为0.19时,供试菌株被聚为2大类群,来自西南地区和东北地区的菌株各自聚为一组,在DNA水平上表现出明显差异,认为是2类不同的致病类群。分子鉴定结果显示引起西南各地区玉米灰斑病的主要致病菌均为玉米尾孢菌Cercospora zeina。表明我国玉米灰斑病菌存在丰富的遗传多样性,ISSR标记可揭示出玉米灰斑病菌株间的亲缘关系及遗传差异性,可用于其遗传多样性研究。
.
DOI:10.13802/j.cnki.zwbhxb.2015.06.007URL [本文引用: 1]
为明确我国发生的玉米灰斑病菌地理差异及遗传结构,利用简单序列重复区间(ISSR)对玉米灰斑病菌遗传多样性进行了分析,并利用尾孢菌特异引物对分离自四川、云南、湖北、贵州等西南地区的16个玉米灰斑病菌菌株进行了分子鉴定。结果显示,通过ISSR标记筛选出10个扩增多态性好且稳定的通用引物,共扩增出81条DNA条带,均为多态性条带,扩增片段大小在200-2 000 bp之间,菌株遗传相似系数为0.19-1.00。在遗传相似系数为0.19时,供试菌株被聚为2大类群,来自西南地区和东北地区的菌株各自聚为一组,在DNA水平上表现出明显差异,认为是2类不同的致病类群。分子鉴定结果显示引起西南各地区玉米灰斑病的主要致病菌均为玉米尾孢菌Cercospora zeina。表明我国玉米灰斑病菌存在丰富的遗传多样性,ISSR标记可揭示出玉米灰斑病菌株间的亲缘关系及遗传差异性,可用于其遗传多样性研究。
.
DOI:10.1007/s001220051637URL [本文引用: 1]
Inter-simple sequence repeat (ISSR) analysis was evaluated for its usefulness in generating markers to extend the genetic linkage map of Citrus using a backcross population previously mapped with restriction fragment length polymorphism (RFLP), random amplified polymorphic DNA (RAPD) and isozyme markers. ISSR markers were obtained through the simple technique of PCR followed by analysis on agarose gels, using simple sequence repeat (SSR) primers. Optimization of reaction conditions was achieved for 50% of the SSR primers screened, and the primers amplified reproducible polymorphic bands in the parents and progeny of the backcross population. Mendelian segregation of the polymorphic bands was demonstrated, with an insignificant number of skewed loci. Most of the SSR primers produced dominant loci; however co-dominance was observed with loci derived from three primers. A new genetic map was produced by combining the segregation data for the ISSR markers and data for the RFLP, RAPD and isozyme markers from the previous map and creating genetic linkages among all the markers using JoinMap 2.0 mapping software. The new map has an improved distribution of markers along the linkage groups with fewer gaps, and marker order showed partial or complete conservation in the linkage groups. The incorporation of ISSR markers into the genetic linkage map demonstrates that ISSR markers are suitable for genetic mapping in Citrus .
.
DOI:10.1017/S1479262115000568URL [本文引用: 1]
Substantial variation in phenotypic traits and ISSR fingerprinting was observed among twenty populations of Achillea fragrantissima (Forssk.) Sch. Bip. in Egypt. Such variation was reflected in the clustering of the examined populations into two major groups: one representing populations in the mountainous area of South Sinai and the other comprising populations growing at low elevations in the middle of Sinai and the desert west of the Suez Canal from Suez in the east to Cairo in the west. Five populations in the eastern part of Sinai near Nuwieba and Taba on the Gulf of Aqaba were loosely assigned to the first group. The populations growing at high elevations in South Sinai, under lower temperature and higher humidity, were characterized by a higher number of total and polymorphic ISSR markers compared with other populations. Unique ISSR markers were more often observed in the fingerprinting of seven populations including five populations growing in the high mountains of Saint Catherine in South Sinai and two populations growing at low elevations but at Wadi Hof south-east of Cairo. Interestingly, unique bands were found in the populations that possessed traits associated with larger plant size and seed yield as well as better vigour. These are important criteria for the selection of A. fragrantissima populations for conservation and sustainable commercial use.
DOI:10.3969/j.issn.1000-1573.2008.01.009URL [本文引用: 1]
对采自河北省14个玉米产区的28个玉米弯孢霉叶斑病标样进行了 分离鉴定,发现新月弯孢(Curuularia lunata)占57.1%,棒状弯孢(C.clauata)占17.9%,画眉草弯孢(C.eragrostidis)占14.3%,中隔弯孢 (C.intermedia)占10.7%.将4种玉米弯孢霉叶斑病菌接种在9个玉米自交系和杂交种上进行寄生适合度测定,发现新月弯孢和中隔弯孢的致病 性最强,其次为棒状弯孢,画眉草弯孢的致病性最弱.用23条随机引物对28个菌株进行RAPD扩增,共获得了185个RAPD标记,其中多态性标记142 个,多态性几率P=76.8%.利用菌株间的遗传相似性进行聚类分析,发现4种弯孢在种间存在的遗传差异可能是导致致病性差异的一个原因.在新月弯孢种内 也存在较大的遗传差异,部分新月弯孢菌株与其它3种弯孢霉亲缘关系较近.
.
DOI:10.3969/j.issn.1000-1573.2008.01.009URL [本文引用: 1]
对采自河北省14个玉米产区的28个玉米弯孢霉叶斑病标样进行了 分离鉴定,发现新月弯孢(Curuularia lunata)占57.1%,棒状弯孢(C.clauata)占17.9%,画眉草弯孢(C.eragrostidis)占14.3%,中隔弯孢 (C.intermedia)占10.7%.将4种玉米弯孢霉叶斑病菌接种在9个玉米自交系和杂交种上进行寄生适合度测定,发现新月弯孢和中隔弯孢的致病 性最强,其次为棒状弯孢,画眉草弯孢的致病性最弱.用23条随机引物对28个菌株进行RAPD扩增,共获得了185个RAPD标记,其中多态性标记142 个,多态性几率P=76.8%.利用菌株间的遗传相似性进行聚类分析,发现4种弯孢在种间存在的遗传差异可能是导致致病性差异的一个原因.在新月弯孢种内 也存在较大的遗传差异,部分新月弯孢菌株与其它3种弯孢霉亲缘关系较近.
URL [本文引用: 1]
在东北三省15个地区采集分离26个玉米弯孢菌菌株,通过鉴别寄主技术与RAPD分子标记技 术进行致病性与遗传分化的研究。结果表明,东北地区玉米弯孢菌在致病性与遗传上都存在明显的变异,二者有一定的相关性,未发现变异程度与菌株地理来源有明 显的直接关系。26个菌株的致病性分为5种类型,其中中等致病类型分布遍及东北三省,为优势致病类群;强致病类型分布在吉林省的白城市;弱致病类型主要分 布在吉林省的白山和梨树。用8个引物对供试菌株进行RAPD扩增,共获得77个RAPD标记,其中多态性标记71个,多态性比例92.2%。供试菌株在相 似系数约0.71处共聚为4组,大多数致病性较强的菌株聚在一起,而弱致病株在相似系数相对较低时和其他菌株聚在一起。
URL [本文引用: 1]
在东北三省15个地区采集分离26个玉米弯孢菌菌株,通过鉴别寄主技术与RAPD分子标记技 术进行致病性与遗传分化的研究。结果表明,东北地区玉米弯孢菌在致病性与遗传上都存在明显的变异,二者有一定的相关性,未发现变异程度与菌株地理来源有明 显的直接关系。26个菌株的致病性分为5种类型,其中中等致病类型分布遍及东北三省,为优势致病类群;强致病类型分布在吉林省的白城市;弱致病类型主要分 布在吉林省的白山和梨树。用8个引物对供试菌株进行RAPD扩增,共获得77个RAPD标记,其中多态性标记71个,多态性比例92.2%。供试菌株在相 似系数约0.71处共聚为4组,大多数致病性较强的菌株聚在一起,而弱致病株在相似系数相对较低时和其他菌株聚在一起。
[本文引用: 1]
.
[本文引用: 1]
DOI:10.7668/hbnxb.2014.03.041Magsci [本文引用: 2]
为明确辽宁省和安徽省玉米弯孢叶斑病菌的生理分化和遗传多样性及其近缘关系。利用UP-PCR、ISSR分子标记方法,对采集于辽宁省和安徽省30株玉米弯孢叶斑病菌进行了遗传多样性分析。从供试引物中筛选获得具有多态性好且稳定的UP-PCR引物4个、ISSR引物12个,分别扩增出 27条谱带和76条谱带,多态性条带比率分别为77.78%和89.47%。聚类分析结果表明,UP-PCR阈值在0.662处菌株被分为7个类群,ISSR阈值在0.681处菌株被分为8个类群,玉米弯孢叶斑病菌存在丰富的遗传变异,与地理来源无明显相关性。从稳定性、可操作性和多态性水平来看,ISSR技术更适合于分析玉米弯孢叶斑病菌的遗传多样性。
.
DOI:10.7668/hbnxb.2014.03.041Magsci [本文引用: 2]
为明确辽宁省和安徽省玉米弯孢叶斑病菌的生理分化和遗传多样性及其近缘关系。利用UP-PCR、ISSR分子标记方法,对采集于辽宁省和安徽省30株玉米弯孢叶斑病菌进行了遗传多样性分析。从供试引物中筛选获得具有多态性好且稳定的UP-PCR引物4个、ISSR引物12个,分别扩增出 27条谱带和76条谱带,多态性条带比率分别为77.78%和89.47%。聚类分析结果表明,UP-PCR阈值在0.662处菌株被分为7个类群,ISSR阈值在0.681处菌株被分为8个类群,玉米弯孢叶斑病菌存在丰富的遗传变异,与地理来源无明显相关性。从稳定性、可操作性和多态性水平来看,ISSR技术更适合于分析玉米弯孢叶斑病菌的遗传多样性。
URL [本文引用: 1]
玉米弯孢霉叶斑病是近年我国玉米生产中发生的一种叶部病害,在黑龙江省已演变为主要病害.通过采集黑龙江省哈尔滨、绥化、大庆、齐齐哈尔和牡丹江等玉米产区病叶标样,经病原菌分离培养、致病性测定、形态鉴定和rDNA-ITS序列分析,明确黑龙江省玉米弯孢霉叶斑病可由新月弯孢Curvularia lunata、棒状弯孢Curvularia clavata、不等弯孢Curvularia inaequalis、苍白弯孢Curvularia pallescens4种弯孢霉引起,且对玉米叶片均具有致病性,其中新月弯孢为优势种,致病力最强.
URL [本文引用: 1]
玉米弯孢霉叶斑病是近年我国玉米生产中发生的一种叶部病害,在黑龙江省已演变为主要病害.通过采集黑龙江省哈尔滨、绥化、大庆、齐齐哈尔和牡丹江等玉米产区病叶标样,经病原菌分离培养、致病性测定、形态鉴定和rDNA-ITS序列分析,明确黑龙江省玉米弯孢霉叶斑病可由新月弯孢Curvularia lunata、棒状弯孢Curvularia clavata、不等弯孢Curvularia inaequalis、苍白弯孢Curvularia pallescens4种弯孢霉引起,且对玉米叶片均具有致病性,其中新月弯孢为优势种,致病力最强.
.
DOI:10.4238/2015.October.16.21URLPMID:26505404 [本文引用: 1]
Abstract To understand the effects of disease-resistant maize varieties and new cropping systems on the population of Curvularia lunata, 52 isolates of C. lunata were collected in China from 2011 to 2013. The isolates were analyzed in terms of phylogenetic relationships, morphology, and pathogenicity. Phylogenetic analysis showed that the 52 isolates clustered into 2 distinct clusters with further subdivisions, suggesting the emergence of new genetic divergence within C. lunata. Results of morphology and pathogenicity analyses demonstrated that there were significant differences among these isolates: 27 isolates were classified as fast growing, 5 as slow growing, and 20 as moderate growing. Three isolates had white-colored colonies, 13 had yellowish green-colored colonies, and the remaining isolates had dark green-colored colonies. Furthermore, conidiation rates were assessed: 30 isolates were characterized as having low conidiation rates, 15 as having medium conidiation rates, and the remaining 7 isolates as having high conidiation rates. Eleven of the isolates appeared to be strongly pathogenic against maize, 15 isolates proved to be weakly pathogenic against maize, and the remaining isolates were regarded to be moderately pathogenic. Interestingly, correlation analysis demonstrated a negative correlation between the growth rate and the pathogenicity of the isolates, while a positive correlation was observed between the conidiation rate and the pathogenicity. No correlation was observed between the colony color and the pathogenicity of the isolates.
DOI:10.1086/282771URL [本文引用: 1]
DOI:10.1073/pnas.70.12.3321URL [本文引用: 1]
[本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2018.01.008URL [本文引用: 1]
【目的】明确不同省(自治区)引起玉米穗腐病的禾谷镰孢复合种(Fusarium graminearum species complex,FGSC)的毒素化学型和它们之间的遗传差异以及亲缘关系。【方法】选用根据基因Tri13和Tri3序列设计的特异性引物分析来自11个省(自治区)的92株禾谷镰孢复合种菌株的毒素化学型;从100条哥伦比亚大学(UBC)开发的通用引物中筛选出13条扩增条带丰富、重复性好、信号强、背景清晰的引物,利用筛选出的引物对供试菌株进行ISSR-PCR扩增,使用Popgen32软件计算多态性位点百分率、Shannon’s多样性指数、群体间的遗传距离和遗传相似性;依据Nei’s遗传距离,利用NTsys2.10e软件进行UPGMA聚类分析,并构建供试菌株的聚类图。【结果】供试的92株禾谷镰孢复合种菌株的产毒素化学型有4种:DON、15-ADON、DON+15-ADON和NIV+15-ADON。其中产生DON和15-ADON的菌株分别为1株和20株;同时产生15-ADON和NIV的有1株;同时产生15-ADON和DON的有55株。筛选出的13条引物对所有供试菌株进行PCR扩增,共获得102条带,其中多态性条带101条,多态性比率为99.02%,平均每条引物产生条带为7.85条。在群体平均水平上,基因多样性指数(H)为0.3129,Shannon’s的信息指数(I)为0.4774,表明禾谷镰孢复合种存在较高的遗传多样性水平;不同地理种群间,各群体的遗传多样性水平具有一定差异,河北、山西、黑龙江和吉林种群遗传多样性最高,安徽和河南种群最低。地理种群间的基因分化系数(Gst)为0.2722,说明不同地理种群间存在一定的遗传变异,但大部分遗传变异(72.78%)发生在种群内。遗传分化系数估算的基因流值Nm=1.3372(〉1),表明不同地理种群间存在一定的基因流动;遗传关系结果表明河北、山西、黑龙江、吉林、辽宁、内蒙古和甘肃菌株群体间亲缘关系较近,山东、江苏、河南和安徽菌株间亲缘关系较近,安17
.
DOI:10.3864/j.issn.0578-1752.2018.01.008URL [本文引用: 1]
【目的】明确不同省(自治区)引起玉米穗腐病的禾谷镰孢复合种(Fusarium graminearum species complex,FGSC)的毒素化学型和它们之间的遗传差异以及亲缘关系。【方法】选用根据基因Tri13和Tri3序列设计的特异性引物分析来自11个省(自治区)的92株禾谷镰孢复合种菌株的毒素化学型;从100条哥伦比亚大学(UBC)开发的通用引物中筛选出13条扩增条带丰富、重复性好、信号强、背景清晰的引物,利用筛选出的引物对供试菌株进行ISSR-PCR扩增,使用Popgen32软件计算多态性位点百分率、Shannon’s多样性指数、群体间的遗传距离和遗传相似性;依据Nei’s遗传距离,利用NTsys2.10e软件进行UPGMA聚类分析,并构建供试菌株的聚类图。【结果】供试的92株禾谷镰孢复合种菌株的产毒素化学型有4种:DON、15-ADON、DON+15-ADON和NIV+15-ADON。其中产生DON和15-ADON的菌株分别为1株和20株;同时产生15-ADON和NIV的有1株;同时产生15-ADON和DON的有55株。筛选出的13条引物对所有供试菌株进行PCR扩增,共获得102条带,其中多态性条带101条,多态性比率为99.02%,平均每条引物产生条带为7.85条。在群体平均水平上,基因多样性指数(H)为0.3129,Shannon’s的信息指数(I)为0.4774,表明禾谷镰孢复合种存在较高的遗传多样性水平;不同地理种群间,各群体的遗传多样性水平具有一定差异,河北、山西、黑龙江和吉林种群遗传多样性最高,安徽和河南种群最低。地理种群间的基因分化系数(Gst)为0.2722,说明不同地理种群间存在一定的遗传变异,但大部分遗传变异(72.78%)发生在种群内。遗传分化系数估算的基因流值Nm=1.3372(〉1),表明不同地理种群间存在一定的基因流动;遗传关系结果表明河北、山西、黑龙江、吉林、辽宁、内蒙古和甘肃菌株群体间亲缘关系较近,山东、江苏、河南和安徽菌株间亲缘关系较近,安17
DOI:10.3864/j.issn.0578-1752.2013.21.015URLMagsci [本文引用: 2]
【目的】针对近年频繁暴发的玉米南方锈病,通过分子生物学技术,明确不同地区玉米南方锈病菌的遗传相似性,为研究该病害流行与发生规律和抗病育种提供信息。【方法】利用18对多态性ISSR引物对2011和2012年采集自12个省(自治区、直辖市)的72个菌株进行扩增,分析种群的遗传结构与特征。【结果】在相似系数0.81水平上,72个菌株被划为2群5亚群10组,亚群间表现出年度与地域的一定差异;遗传多样性分析表明菌株群体间存在较高的遗传变异;遗传相似性和遗传距离分析揭示山东与安徽、福建与浙江群体分别具有相对较高的相似性,而海南群体与其它群体遗传距离最远;不同地区病菌群体间缺乏基因交流,遗传相关性较低。【结论】引起中国不同地区玉米南方锈病的多堆柄锈菌群体具有较高的遗传变异;病菌遗传多样性分析结果初步证明,各地病菌无法在当地完成周年侵染循环;海南三亚冬季发生的玉米南方锈病不是中国其它地区病害的初侵染源,广东和广西等华南地区的玉米锈病也不是黄淮夏玉米区病害的初侵染源,中国大陆地区和海南岛的玉米南方锈病的初侵染源可能来自于中国大陆以外的其它地区。
DOI:10.3864/j.issn.0578-1752.2013.21.015URLMagsci [本文引用: 2]
【目的】针对近年频繁暴发的玉米南方锈病,通过分子生物学技术,明确不同地区玉米南方锈病菌的遗传相似性,为研究该病害流行与发生规律和抗病育种提供信息。【方法】利用18对多态性ISSR引物对2011和2012年采集自12个省(自治区、直辖市)的72个菌株进行扩增,分析种群的遗传结构与特征。【结果】在相似系数0.81水平上,72个菌株被划为2群5亚群10组,亚群间表现出年度与地域的一定差异;遗传多样性分析表明菌株群体间存在较高的遗传变异;遗传相似性和遗传距离分析揭示山东与安徽、福建与浙江群体分别具有相对较高的相似性,而海南群体与其它群体遗传距离最远;不同地区病菌群体间缺乏基因交流,遗传相关性较低。【结论】引起中国不同地区玉米南方锈病的多堆柄锈菌群体具有较高的遗传变异;病菌遗传多样性分析结果初步证明,各地病菌无法在当地完成周年侵染循环;海南三亚冬季发生的玉米南方锈病不是中国其它地区病害的初侵染源,广东和广西等华南地区的玉米锈病也不是黄淮夏玉米区病害的初侵染源,中国大陆地区和海南岛的玉米南方锈病的初侵染源可能来自于中国大陆以外的其它地区。
URL [本文引用: 1]
利用11对多态性ISSR引物对2014–2016年间采自山东、江苏、浙江、河南及海南等5省的60个玉米多堆柄锈菌菌株进行了种群遗传多样性分析。结果表明,分离自山东的玉米多堆柄锈菌菌株的遗传多样性丰富,多态性百分率(P)达96.34%,与江苏、浙江、河南锈菌种群具有较高的相似性,遗传距离相对较近,其中与江苏的群体遗传相似系数最高(GS=0.7829),遗传距离最近(GD=0.2447),而与分离自海南的锈菌种群相似性最低(GS=0.0148),遗传距离最远(GD=4.2999)。在相似性系数0.74水平上,供试60个菌株被划为3个群和多个亚群组,各群和亚群间表现出较明显的年度与地理分布上的差异。2015年采集的菌株之间G_(st)值为0.8694,2015与2016年采集的菌株群体间的Gst值为0.4562,说明病菌的遗传变异不仅表现在地域上,也体现在年度间。基于ISSR标记分析,作者认为发生在山东省的玉米南方锈病其初侵染菌源不是来自我国周年发生玉米南方锈病的海南省,而最有可能来自同样能越冬的菲律宾或我国台湾地区。
.
URL [本文引用: 1]
利用11对多态性ISSR引物对2014–2016年间采自山东、江苏、浙江、河南及海南等5省的60个玉米多堆柄锈菌菌株进行了种群遗传多样性分析。结果表明,分离自山东的玉米多堆柄锈菌菌株的遗传多样性丰富,多态性百分率(P)达96.34%,与江苏、浙江、河南锈菌种群具有较高的相似性,遗传距离相对较近,其中与江苏的群体遗传相似系数最高(GS=0.7829),遗传距离最近(GD=0.2447),而与分离自海南的锈菌种群相似性最低(GS=0.0148),遗传距离最远(GD=4.2999)。在相似性系数0.74水平上,供试60个菌株被划为3个群和多个亚群组,各群和亚群间表现出较明显的年度与地理分布上的差异。2015年采集的菌株之间G_(st)值为0.8694,2015与2016年采集的菌株群体间的Gst值为0.4562,说明病菌的遗传变异不仅表现在地域上,也体现在年度间。基于ISSR标记分析,作者认为发生在山东省的玉米南方锈病其初侵染菌源不是来自我国周年发生玉米南方锈病的海南省,而最有可能来自同样能越冬的菲律宾或我国台湾地区。
URL [本文引用: 1]
对种子带菌检验、病原菌在病残 体上越冬形态观察及存活检测、越冬病菌的苗期致病力测定表明,玉米种子不携带弯孢霉叶斑病菌(Curvularialunata),其分生孢子能在玉米秸 秆垛中越冬,病残体内的菌丝体能在多种场所越冬,越冬病菌接种幼苗显示了该病典型的病斑。说明玉米秸秆垛、田间和堆肥中的病残体是玉米弯孢霉叶斑病的初侵 染来源。
.
URL [本文引用: 1]
对种子带菌检验、病原菌在病残 体上越冬形态观察及存活检测、越冬病菌的苗期致病力测定表明,玉米种子不携带弯孢霉叶斑病菌(Curvularialunata),其分生孢子能在玉米秸 秆垛中越冬,病残体内的菌丝体能在多种场所越冬,越冬病菌接种幼苗显示了该病典型的病斑。说明玉米秸秆垛、田间和堆肥中的病残体是玉米弯孢霉叶斑病的初侵 染来源。
DOI:10.3969/j.issn.1000-5684.2002.04.013URLMagsci [本文引用: 1]
采用不同的方法对玉米弯孢霉叶斑病(<i>Curvlarialunata</i>)的初侵染来源进行了初步研究。结果表明:玉米种子内外均不带菌,不能作为病害的初侵染来源;病残体内的病菌能在地表越冬,作为翌年的初侵染来源;不同深度土壤中病残体上的病菌存活时间较短,不能越冬作为下翌年的初侵染来源。生长于PSA培养基上的病菌培养物在地表和不同深度的土壤中均能越冬,说明病菌的分生孢子可在不同环境下越冬,作为翌年的初侵染源。对4种杀菌剂进行室内毒力测定的结果表明:供试药剂中以50%福美双可湿性粉剂的EC<sub>50</sub>值最小,为11.79mg/kg,毒力最高,其毒力回归方程为<i>y=3.33+1.56x,相关系数r</i>为0.9912,达极显著水平。其余供试药剂毒力大小依次为:75%百菌清可湿性粉剂,其毒力回归方程为<i>y=3.78+1.04x,相关系数r</i>为0.9579,达极显著水平,EC<sub>50</sub>值为14.89mg/kg;75%代森锌,其毒力回归方程为<i>y=4.01+0.63x,相关系数r</i>为0.9555,达极显著水平,EC<sub>50</sub>值为44.57mg/kg;75%代森锰锌可湿性粉剂,其毒力回归方程为<i>y=2.08+1.61x,相关系数r</i>为0.9670,达极显著水平,EC<sub>50</sub>值为64.10mg/kg。
.
DOI:10.3969/j.issn.1000-5684.2002.04.013URLMagsci [本文引用: 1]
采用不同的方法对玉米弯孢霉叶斑病(<i>Curvlarialunata</i>)的初侵染来源进行了初步研究。结果表明:玉米种子内外均不带菌,不能作为病害的初侵染来源;病残体内的病菌能在地表越冬,作为翌年的初侵染来源;不同深度土壤中病残体上的病菌存活时间较短,不能越冬作为下翌年的初侵染来源。生长于PSA培养基上的病菌培养物在地表和不同深度的土壤中均能越冬,说明病菌的分生孢子可在不同环境下越冬,作为翌年的初侵染源。对4种杀菌剂进行室内毒力测定的结果表明:供试药剂中以50%福美双可湿性粉剂的EC<sub>50</sub>值最小,为11.79mg/kg,毒力最高,其毒力回归方程为<i>y=3.33+1.56x,相关系数r</i>为0.9912,达极显著水平。其余供试药剂毒力大小依次为:75%百菌清可湿性粉剂,其毒力回归方程为<i>y=3.78+1.04x,相关系数r</i>为0.9579,达极显著水平,EC<sub>50</sub>值为14.89mg/kg;75%代森锌,其毒力回归方程为<i>y=4.01+0.63x,相关系数r</i>为0.9555,达极显著水平,EC<sub>50</sub>值为44.57mg/kg;75%代森锰锌可湿性粉剂,其毒力回归方程为<i>y=2.08+1.61x,相关系数r</i>为0.9670,达极显著水平,EC<sub>50</sub>值为64.10mg/kg。
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
水稻黑粒米是由弯孢霉属真菌引起的一种水稻生育后期的重要病害,影响稻米品质和稻谷产量,作者从我国主要产稻省稻田采集的变色谷样品上,分离到大量弯孢霉,并鉴定出13个种。水稻及禾本科杂草的下部枯黄叶上均带有较多的弯孢霉孢子,禾本科杂草带菌远较水稻为多。水稻穗期稻谷上弯孢霉的平均检出率早稻为17.2%,晚稻为40.2%,可用70%代森锰锌等药进行防治。
.
URL [本文引用: 1]
水稻黑粒米是由弯孢霉属真菌引起的一种水稻生育后期的重要病害,影响稻米品质和稻谷产量,作者从我国主要产稻省稻田采集的变色谷样品上,分离到大量弯孢霉,并鉴定出13个种。水稻及禾本科杂草的下部枯黄叶上均带有较多的弯孢霉孢子,禾本科杂草带菌远较水稻为多。水稻穗期稻谷上弯孢霉的平均检出率早稻为17.2%,晚稻为40.2%,可用70%代森锰锌等药进行防治。
DOI:10.3969/j.issn.10017216.2015.02.014URLMagsci [本文引用: 1]
水稻穗腐病和穗枯病是我国新上升或新出现的两个水稻后期穗部病害。近年来,随着气候的变化、耕作栽培制度及肥水管理的改变以及品种(组合)的更替,两病在我国的发生和危害程度越来越严重。穗腐病和穗枯病病原菌有色,可产生毒素,不但直接影响水稻产量,还导致稻米品质降低,危害食用者的健康。穗腐病和穗枯病在发病时期、发生部位和危害程度上具有相似性。综述了两病害的发生、流行规律、危害状况、病原菌生物特性和毒素研究以及防治方法,提出了研究上目前存在的问题和今后的研究方向。
DOI:10.3969/j.issn.10017216.2015.02.014URLMagsci [本文引用: 1]
水稻穗腐病和穗枯病是我国新上升或新出现的两个水稻后期穗部病害。近年来,随着气候的变化、耕作栽培制度及肥水管理的改变以及品种(组合)的更替,两病在我国的发生和危害程度越来越严重。穗腐病和穗枯病病原菌有色,可产生毒素,不但直接影响水稻产量,还导致稻米品质降低,危害食用者的健康。穗腐病和穗枯病在发病时期、发生部位和危害程度上具有相似性。综述了两病害的发生、流行规律、危害状况、病原菌生物特性和毒素研究以及防治方法,提出了研究上目前存在的问题和今后的研究方向。
URL [本文引用: 1]
2011年7月,在湖南省花垣县水稻上发现一种新的病害,症状表现为叶鞘上出现长椭圆形黑斑,病健交界模糊.分离到的4隔孢弯孢霉(HNHY001)经离体和活体人工接种均可产生黑斑;另一种黑孢霉菌株(HNHY002)不引致叶鞘黑斑.从湖南省栽培水稻150份种子样本中共分离到弯孢霉菌株27个,其中4个菌株具有4隔孢子.这4个菌株中1个菌株不产生子座,其余3个均产生分枝子座.经人工接种除不产生子座的外,其余3个菌株均能产生典型的叶鞘黑斑.HNHY001的rDNA-ITS序列(GenBank登录号JQ360963)经BLAST搜索,与之最接近的几个序列为膝曲旋孢腔菌(Cochliobolusgeniculatus)及其有丝分裂产孢种—4隔孢弯孢霉.鉴于单独以及与其它4隔孢弯孢霉菌株进行对峙培养均未产生有性阶段,建议根据此菌无性阶段特征即分生孢子较直、4横隔、子座分枝等特征,鉴定为膝曲旋孢腔菌有丝分裂产孢种之一的假弯孢(C.fallax).水稻叶鞘黑斑病系国内外首次报道.
URL [本文引用: 1]
2011年7月,在湖南省花垣县水稻上发现一种新的病害,症状表现为叶鞘上出现长椭圆形黑斑,病健交界模糊.分离到的4隔孢弯孢霉(HNHY001)经离体和活体人工接种均可产生黑斑;另一种黑孢霉菌株(HNHY002)不引致叶鞘黑斑.从湖南省栽培水稻150份种子样本中共分离到弯孢霉菌株27个,其中4个菌株具有4隔孢子.这4个菌株中1个菌株不产生子座,其余3个均产生分枝子座.经人工接种除不产生子座的外,其余3个菌株均能产生典型的叶鞘黑斑.HNHY001的rDNA-ITS序列(GenBank登录号JQ360963)经BLAST搜索,与之最接近的几个序列为膝曲旋孢腔菌(Cochliobolusgeniculatus)及其有丝分裂产孢种—4隔孢弯孢霉.鉴于单独以及与其它4隔孢弯孢霉菌株进行对峙培养均未产生有性阶段,建议根据此菌无性阶段特征即分生孢子较直、4横隔、子座分枝等特征,鉴定为膝曲旋孢腔菌有丝分裂产孢种之一的假弯孢(C.fallax).水稻叶鞘黑斑病系国内外首次报道.
URL [本文引用: 1]
本文报道了弯孢菌引起的变色米的症状,弯孢苗的种类及其对水稻的致病性。变色米的症状,因水稻品种、弯孢菌种类和接种方法而有差异,但一般在糙米上为浅至深色的褐色米;在稻谷上产生全褐型、半褐型、褐斑型、褐点型和退色型五种类型。后各地采集的变色谷(米)上分离到弯孢属9个种,即新月弯孢、膝曲弯孢、近缘弯孢、不正弯孢、中隔弯孢、棒状弯孢、苍白弯孢、画眉弯孢、假弯孢,其中6个种为国内首次报道。据在水稻孕穗至开花期人工接种结果,上述9个种对水稻均有致病性,并产生大量的变色谷(米)。
URL [本文引用: 1]
本文报道了弯孢菌引起的变色米的症状,弯孢苗的种类及其对水稻的致病性。变色米的症状,因水稻品种、弯孢菌种类和接种方法而有差异,但一般在糙米上为浅至深色的褐色米;在稻谷上产生全褐型、半褐型、褐斑型、褐点型和退色型五种类型。后各地采集的变色谷(米)上分离到弯孢属9个种,即新月弯孢、膝曲弯孢、近缘弯孢、不正弯孢、中隔弯孢、棒状弯孢、苍白弯孢、画眉弯孢、假弯孢,其中6个种为国内首次报道。据在水稻孕穗至开花期人工接种结果,上述9个种对水稻均有致病性,并产生大量的变色谷(米)。
DOI:10.3321/j.issn:0577-7518.2005.03.011URLMagsci [本文引用: 1]
杂交狗牙根叶枯病是武汉地区高尔夫球场草坪上的一种重要病害。为了查明该病害的病原和控制该病害的发生,对叶枯病的病原物进行了分离鉴定、生物学特性研究以及室内药剂筛选。形态学观察结果表明,引起杂交狗牙根叶枯病的病原物为新月弯孢<em>Curvularia lunata </em>(Wakker)Boedijn;生物学特性研究显示<em>C.lunata</em>生长的适宜温度为20~35℃,最适温度为25℃,适宜pH值为5~8,最适pH值为6;在所测试的碳源中,<em>C.lunata</em>对葡萄糖和蔗糖的利用最好,对半乳糖的利用效果最差;在所测试的氮源中,对KNO<sub>3</sub>利用最好,对NH<sub>4</sub>Cl和氨基乙酸的利用最差;菌丝的致死温度为55℃ 10min。室内PDA平板药剂筛选结果表明,10%苯醚甲环唑对病菌菌丝生长的抑制效果最好。
DOI:10.3321/j.issn:0577-7518.2005.03.011URLMagsci [本文引用: 1]
杂交狗牙根叶枯病是武汉地区高尔夫球场草坪上的一种重要病害。为了查明该病害的病原和控制该病害的发生,对叶枯病的病原物进行了分离鉴定、生物学特性研究以及室内药剂筛选。形态学观察结果表明,引起杂交狗牙根叶枯病的病原物为新月弯孢<em>Curvularia lunata </em>(Wakker)Boedijn;生物学特性研究显示<em>C.lunata</em>生长的适宜温度为20~35℃,最适温度为25℃,适宜pH值为5~8,最适pH值为6;在所测试的碳源中,<em>C.lunata</em>对葡萄糖和蔗糖的利用最好,对半乳糖的利用效果最差;在所测试的氮源中,对KNO<sub>3</sub>利用最好,对NH<sub>4</sub>Cl和氨基乙酸的利用最差;菌丝的致死温度为55℃ 10min。室内PDA平板药剂筛选结果表明,10%苯醚甲环唑对病菌菌丝生长的抑制效果最好。
DOI:10.3969/j.issn.1000-4440.2005.03.015URL [本文引用: 1]
以新月弯孢[Curvularia lunata(Wakker) Boedijn]为病原,接种亚热带地区9种常见禾草,观测它们的生长情况、潜育期、发病部位、病斑大小、病斑密度等指标,综合评价供试草的抗感性.结果表明,弯孢叶斑病菌可以侵染供试草种,使禾草生长速度降低、新生叶片形成慢、叶片短而窄、节间缩短,但供试禾草的潜育期(4~10 d,变异系数29.54%)和症状的明显程度不同,潜育期由短到长依次为:剪股颖、狗牙根、高羊茅、牛鞭草、鸭茅、多花黑麦草、多年生黑麦草、一年生早熟禾、假俭草;症状明显程度依次为:剪股颖、狗牙根、多花黑麦草、鸭茅、高羊茅、牛鞭草、一年生早熟禾、多年生黑麦草、假俭草.供试禾草的抗感性:假俭草为高抗(R),多年生黑麦草、一年生早熟禾为中抗(MR);高羊茅为中感(MS);牛鞭草为感病(S);剪股颖、狗牙根、多花黑麦草和鸭茅为高感(HS).
DOI:10.3969/j.issn.1000-4440.2005.03.015URL [本文引用: 1]
以新月弯孢[Curvularia lunata(Wakker) Boedijn]为病原,接种亚热带地区9种常见禾草,观测它们的生长情况、潜育期、发病部位、病斑大小、病斑密度等指标,综合评价供试草的抗感性.结果表明,弯孢叶斑病菌可以侵染供试草种,使禾草生长速度降低、新生叶片形成慢、叶片短而窄、节间缩短,但供试禾草的潜育期(4~10 d,变异系数29.54%)和症状的明显程度不同,潜育期由短到长依次为:剪股颖、狗牙根、高羊茅、牛鞭草、鸭茅、多花黑麦草、多年生黑麦草、一年生早熟禾、假俭草;症状明显程度依次为:剪股颖、狗牙根、多花黑麦草、鸭茅、高羊茅、牛鞭草、一年生早熟禾、多年生黑麦草、假俭草.供试禾草的抗感性:假俭草为高抗(R),多年生黑麦草、一年生早熟禾为中抗(MR);高羊茅为中感(MS);牛鞭草为感病(S);剪股颖、狗牙根、多花黑麦草和鸭茅为高感(HS).
DOI:10.3969/j.issn.1001-411X.2005.02.008URL [本文引用: 1]
2 0 0 1~2 0 0 3年,对广东省境内狗牙根草Cynodondactylon (L .)Pers.的真菌病害进行了调查,共鉴定出9种病原真菌,分别是灰葡萄孢BotrytiscinereaPers.exFr.、禾生炭疽 Colletotrichumgraminicola (Ces.)Wilson、棒弯孢霉CurvulariaclavataJain、新月弯孢霉C .lunataNelsonHaasis、疣弯孢霉C .verruculosus (TsudaUeyama)Sivan .、球黑孢Nigrosporasphaerica (Sacc.)Mason、侧生团黑孢PericonialateralisEllisErerh、狗牙根柄锈菌 PucciniacynodontisLacroixexDesm .和立枯丝核菌RhizoctoniasolaniK櫣hn.,生产上以3种弯孢霉引发的病害普遍且较重
DOI:10.3969/j.issn.1001-411X.2005.02.008URL [本文引用: 1]
2 0 0 1~2 0 0 3年,对广东省境内狗牙根草Cynodondactylon (L .)Pers.的真菌病害进行了调查,共鉴定出9种病原真菌,分别是灰葡萄孢BotrytiscinereaPers.exFr.、禾生炭疽 Colletotrichumgraminicola (Ces.)Wilson、棒弯孢霉CurvulariaclavataJain、新月弯孢霉C .lunataNelsonHaasis、疣弯孢霉C .verruculosus (TsudaUeyama)Sivan .、球黑孢Nigrosporasphaerica (Sacc.)Mason、侧生团黑孢PericonialateralisEllisErerh、狗牙根柄锈菌 PucciniacynodontisLacroixexDesm .和立枯丝核菌RhizoctoniasolaniK櫣hn.,生产上以3种弯孢霉引发的病害普遍且较重
DOI:10.3969/j.issn.1001-0629.2003.12.021URL [本文引用: 1]
以我国北方常见的冷季型草坪草高羊茅(Festucaarundinacea)、多年生黑麦草(Loliumperenne)和草地早熟禾(Poapratensis)为材料,将细交链孢(Alternariaalternata)、根腐离蠕孢(Bipolarissorokiniana)、枝孢(Cladosporiumherbarum)、新月弯孢(Curvularialunata)、德氏霉(Drechslerasp )、燕麦镰孢(Fusariumave naceum)、锐顶镰孢(F acuminatum)、尖镰孢(F oxysporum),腐皮镰孢(F solani)和粉红粘帚霉(Glio cldiumro
DOI:10.3969/j.issn.1001-0629.2003.12.021URL [本文引用: 1]
以我国北方常见的冷季型草坪草高羊茅(Festucaarundinacea)、多年生黑麦草(Loliumperenne)和草地早熟禾(Poapratensis)为材料,将细交链孢(Alternariaalternata)、根腐离蠕孢(Bipolarissorokiniana)、枝孢(Cladosporiumherbarum)、新月弯孢(Curvularialunata)、德氏霉(Drechslerasp )、燕麦镰孢(Fusariumave naceum)、锐顶镰孢(F acuminatum)、尖镰孢(F oxysporum),腐皮镰孢(F solani)和粉红粘帚霉(Glio cldiumro
URL [本文引用: 1]
1993-1995年调查了福建暖季型草坪草狗牙根、细叶结缕草、沟叶结缕草结缕草、假俭草和钝叶草栽培品种的产现害、病原种类。发现病害14种,确证病原13种。其中狗牙根白化病的危害面积最大,黑斑病病原新月弯孢霉的分离频率最高,结缕草锈病发生较普遍。同时,明确了棉蚜虫害也可以引直民狗牙根白化。
URL [本文引用: 1]
1993-1995年调查了福建暖季型草坪草狗牙根、细叶结缕草、沟叶结缕草结缕草、假俭草和钝叶草栽培品种的产现害、病原种类。发现病害14种,确证病原13种。其中狗牙根白化病的危害面积最大,黑斑病病原新月弯孢霉的分离频率最高,结缕草锈病发生较普遍。同时,明确了棉蚜虫害也可以引直民狗牙根白化。
DOI:10.3969/j.issn.1002-2090.2014.01.001URL [本文引用: 1]
为明确黑龙江和吉林省稻瘟病菌种群多样性,采用Pot2- Rep-PCR方法对来自黑龙江省和吉林省部分稻区49个稻瘟病菌菌株的DNA进行指纹分析。研究结果表明,供试的稻瘟病菌种群在DNA水平上变异较大, 存在丰富的多态性;在20个遗传谱中第2个谱系为优势谱系,包含黑龙江省11个菌株,吉林省9个菌株,其他19个系谱的菌株数为1~10个不等。谱系3菌 株既是黑龙江省菌株,又是黑龙江省特有的谱系类型。聚类分析结果表明,参试稻瘟菌株遗传多样性程度(系谱数/菌株数)黑龙江省(7 /23=0.3043)、吉林省(11/26=0.4230),吉林省稻瘟病菌的多样性要强于黑龙江省稻瘟病菌。
DOI:10.3969/j.issn.1002-2090.2014.01.001URL [本文引用: 1]
为明确黑龙江和吉林省稻瘟病菌种群多样性,采用Pot2- Rep-PCR方法对来自黑龙江省和吉林省部分稻区49个稻瘟病菌菌株的DNA进行指纹分析。研究结果表明,供试的稻瘟病菌种群在DNA水平上变异较大, 存在丰富的多态性;在20个遗传谱中第2个谱系为优势谱系,包含黑龙江省11个菌株,吉林省9个菌株,其他19个系谱的菌株数为1~10个不等。谱系3菌 株既是黑龙江省菌株,又是黑龙江省特有的谱系类型。聚类分析结果表明,参试稻瘟菌株遗传多样性程度(系谱数/菌株数)黑龙江省(7 /23=0.3043)、吉林省(11/26=0.4230),吉林省稻瘟病菌的多样性要强于黑龙江省稻瘟病菌。
[J].
[本文引用: 1]
[本文引用: 1]
[D].Yaan: Sichuan Agricultural University,
[本文引用: 1]
.
DOI:10.1094/PHYTO-97-5-0574URLPMID:18943576 [本文引用: 2]
Abstract ABSTRACT Isolates of Puccinia triticina collected from durum wheat from Argentina, Chile, Ethiopia, France, Mexico, Spain, and the United States were analyzed with 11 simple sequence repeat (SSR) markers in order to determine the genetic relationship among isolates. These isolates also were compared with P. triticina isolates from common wheat from North America, and an isolate collected from Aegilops speltoides from Israel, to determine genetic relationships among groups of P. triticina found on different telial hosts. The large majority of isolates from durum wheat were identical for SSR markers or had 70% genetic dissimilarity from isolates from durum wheat, and the isolate from A. speltoides was >90% dissimilar from all isolates tested. Analysis of molecular variance tests showed significant levels (P = 0.001) of genetic differentiation among regions and among isolates within countries. Isolates of P. triticina from durum wheat from South America, North America, and Europe were closely related based on SSR genotypes, suggesting a recent common ancestor, whereas P. triticina from Ethiopia, common wheat, and A. speltoides each had distinct SSR genotypes, which suggested different origins.
DOI:10.1186/1471-2148-12-76URLPMID:3457899 [本文引用: 1]
Background Powdery mildew, caused by the obligate biotrophic fungus Blumeria graminis, is a major problem in cereal production as it can reduce quality and yield. B. graminis has evolved eight distinct formae speciales (f.sp.) which display strict host specialization. In the last decade, powdery mildew has emerged on triticale, the artificial intergeneric hybrid between wheat and rye. This emergence is probably triggered by a host range expansion of the wheat powdery mildew B. graminis f.sp. tritici. To gain more precise information about the evolutionary processes that led to this host range expansion, we pursued a combined pathological and genetic approach. Results B. graminis isolates were sampled from triticale, wheat and rye from different breeding regions in Europe. Pathogenicity tests showed that isolates collected from triticale are highly pathogenic on most of the tested triticale cultivars. Moreover, these isolates were also able to infect several wheat cultivars (their previous hosts), although a lower aggressiveness was observed compared to isolates collected from wheat. Phylogenetic analysis of nuclear gene regions identified two statistically significant clades, which to a certain extent correlated with pathogenicity. No differences in virulence profiles were found among the sampled regions, but the distribution of genetic variation demonstrated to be geography dependent. A multilocus haplotype network showed that haplotypes pathogenic on triticale are distributed at different sites in the network, but always clustered at or near the tips of the network. Conclusions This study reveals a genetic structure in B. graminis with population differentiation according to geography and host specificity. In addition, evidence is brought forward demonstrating that the host range expansion of wheat isolates to the new host triticale occurred recently and multiple times at different locations in Europe.
URL [本文引用: 1]
为了明确甘肃玉米大斑病菌生理小种与交配型的结构组成,采用单基因鉴别寄主测定了甘肃4个典型生态区57株玉米大斑病菌的抗、感反应型,对峙培养显微观测法测定了它们与标准菌株杂交是否产生有性态子囊壳。结果表明,甘肃玉米大斑病菌生理小种结构组成十分复杂,57株病菌中共发现0、1、12、23、123、N、1N、2N、3N、123N共10个生理小种,0号以61.40%分布频率成为优势生理小种,1号以17.54%分布频率成为次优势生理小种,23号生理小种出现频率仅为5.26%、123和N号生理小种出现频率仅为3.51%,12、1N、2N、3N、123N生理小种出现频率仅为1.75%,4个典型生态区中河西灌溉绿洲生态区生理小种结构组成最为复杂,15株病菌中共检出0、1、N、12、1N、2N、3N、123N 8个生理小种,中部干旱雨养、陇东半湿润半干旱和南部湿润生态区生理小种结构组成相对简单,42株病菌共检出0、1、23、123和N号5个生理小种。相对而言,甘肃玉米大斑病菌交配型结构组成简单,57株病菌中共检出A型22株、a型24株, Aa型11株,分布频率分别为38.6%、42.1%和19.3%;其中河西灌溉绿洲、中部干旱雨养和陇东半湿润半干旱生态区45株病菌共检出18株A型、18株a型和9株Aa型,A∶a比例完全符合1∶1,南部湿润生态区12株病菌共检出4株A型、6株a型和2株Aa型,A∶a比例小于1∶1。
.
URL [本文引用: 1]
为了明确甘肃玉米大斑病菌生理小种与交配型的结构组成,采用单基因鉴别寄主测定了甘肃4个典型生态区57株玉米大斑病菌的抗、感反应型,对峙培养显微观测法测定了它们与标准菌株杂交是否产生有性态子囊壳。结果表明,甘肃玉米大斑病菌生理小种结构组成十分复杂,57株病菌中共发现0、1、12、23、123、N、1N、2N、3N、123N共10个生理小种,0号以61.40%分布频率成为优势生理小种,1号以17.54%分布频率成为次优势生理小种,23号生理小种出现频率仅为5.26%、123和N号生理小种出现频率仅为3.51%,12、1N、2N、3N、123N生理小种出现频率仅为1.75%,4个典型生态区中河西灌溉绿洲生态区生理小种结构组成最为复杂,15株病菌中共检出0、1、N、12、1N、2N、3N、123N 8个生理小种,中部干旱雨养、陇东半湿润半干旱和南部湿润生态区生理小种结构组成相对简单,42株病菌共检出0、1、23、123和N号5个生理小种。相对而言,甘肃玉米大斑病菌交配型结构组成简单,57株病菌中共检出A型22株、a型24株, Aa型11株,分布频率分别为38.6%、42.1%和19.3%;其中河西灌溉绿洲、中部干旱雨养和陇东半湿润半干旱生态区45株病菌共检出18株A型、18株a型和9株Aa型,A∶a比例完全符合1∶1,南部湿润生态区12株病菌共检出4株A型、6株a型和2株Aa型,A∶a比例小于1∶1。
[本文引用: 1]
DOI:10.3969/j.issn.0253-2654.2002.02.015URL [本文引用: 1]
玉米大斑病长蠕孢存在生理分化现象,人们在研究该菌的同时发现其生理小种存在着遗传变异.论文主要从三个 方面进行了综述:玉米大斑病长蠕孢生理小种划分标准的演变、我国玉米大斑病长蠕孢生理小种研究的历史与现状和玉米大斑病长蠕孢生理小种变异的原因.指出, 玉米大斑病长蠕孢生理小种变异的原因主要有:基因突变、生殖方式、选择作用和基因漂移等.
DOI:10.3969/j.issn.0253-2654.2002.02.015URL [本文引用: 1]
玉米大斑病长蠕孢存在生理分化现象,人们在研究该菌的同时发现其生理小种存在着遗传变异.论文主要从三个 方面进行了综述:玉米大斑病长蠕孢生理小种划分标准的演变、我国玉米大斑病长蠕孢生理小种研究的历史与现状和玉米大斑病长蠕孢生理小种变异的原因.指出, 玉米大斑病长蠕孢生理小种变异的原因主要有:基因突变、生殖方式、选择作用和基因漂移等.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}