 ,1
,1Characteristics, Developmental Expression and RNAi Effect Analysis of a Novel Trehalose-6-Phosphate Synthase Gene in Nilaparvata lugens
TANG Bin1,2, SHEN QiDa2, ZENG BoPing1, XIAO ZhongJiu1, QIU LingYu2, PAN BiYing2, LI Kun1, ZHANG DaoWei,1通讯作者:
收稿日期:2018-09-11接受日期:2018-10-23网络出版日期:2019-02-13
| 基金资助: |
Received:2018-09-11Accepted:2018-10-23Online:2019-02-13
作者简介 About authors
唐斌,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (7281KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
唐斌, 沈祺达, 曾伯平, 肖仲久, 邱玲玉, 潘碧莹, 李昆, 张道伟. 褐飞虱一个新的海藻糖合成酶基因的特性、 发育表达及RNAi效果分析[J]. 中国农业科学, 2019, 52(3): 466-477 doi:10.3864/j.issn.0578-1752.2019.03.007
TANG Bin, SHEN QiDa, ZENG BoPing, XIAO ZhongJiu, QIU LingYu, PAN BiYing, LI Kun, ZHANG DaoWei.
0 引言
【研究意义】海藻糖是昆虫体内的血糖,其含量高达体内糖类物质的80%以上,在昆虫的发育及生理活动中起着非常重要的作用[1,2,3,4]。研究褐飞虱(Nilaparvata lugens)不同海藻糖合成酶基因的具体功能和功能差异,可促进昆虫海藻糖合成途径进化及多样性研究的发展。明确新的海藻糖合成酶基因的特性和功能,可为褐飞虱防治提供新思路、新方向。【前人研究进展】昆虫中能够合成海藻糖最早在1961年时被发现,当时STEELE将心侧体的组织液提取物注射到美洲大蠊(Periplaneta americana)体内,结果直接导致了血淋巴中的海藻糖含量上升[5]。1963年,BOWERS等在对另一种蟑螂(Blaberus discoidalis)的研究中也发现上述现象[6]。此后的一系列研究表明,主要由于昆虫心侧体释放出神经肽(Hypertre- halosaemic)到血淋巴中,对脂肪体产生刺激,使其开始合成海藻糖,并且将合成的海藻糖输送到昆虫血淋巴中[7,8,9]。昆虫中的TPS和TPP,其蛋白序列都具有较高的保守性,与酿酒酵母(Saccharomyces cerevisiae)和大肠杆菌(Escherichia coli)海藻糖合成酶途径中的OtsA和OtsB相对应[8,10-11]。目前,已在多种昆虫体内发现并克隆出海藻糖合成酶基因,如甜菜夜蛾(Spodoptera exigua)、埃及伊蚊(Aedes aegypti)、家蚕(Bombyx mori)、黑腹果蝇(Drosophila melanogaster)、马铃薯甲虫(Leptinotarsa decemlineata)、东亚飞蝗(Locusta migratoria manilensis)、白背飞虱(Sogatella furcifera)、柑橘大实蝇(Bactrocera minax)、赤拟谷盗(Tribolium castaneum)等,其中德国小蠊(Blattella germanica)、金小蜂(Nasonia vitripennis)和褐飞虱中均发现和报道了两个不同的TPS [3,12-18]。【本研究切入点】褐飞虱为水稻最具破坏力的害虫之一[19],在20世纪中叶,亚洲褐飞虱仅在部分国家暴发,但自70年代至今,褐飞虱持续危害我国长江流域大面积的水稻,西南地区以及华南地区也多次暴发,并且有越来越严重的趋势;2012年在浙江金华、2014年在四川南充和浙江台州、2015年在云南多地均发生了大规模的褐飞虱危害[7,20-24]。褐飞虱为单食性害虫,只以水稻(Oryza sativa)或普通野生稻(Oryza rufipogon)为食,且具有繁殖速度快、生命周期短以及环境适应性强的特点[25]。褐飞虱通过其口器穿刺直接吸食水稻下部茎秆的汁液,攫取水稻养分,其取食及产卵行为会在水稻茎秆表皮上产生大量伤口,破坏输导组织,导致水稻茎秆内的养分和水分输送受阻。受创的水稻极易受到水稻纹枯病以及小球菌核病等水稻疾病的感染,对水稻的生长造成极大的危害,造成减产乃至绝收[7,26-27]。近年来,随着褐飞虱基因组测序的完成[28],褐飞虱迁飞相关的长翅型和短翅型调控机理等越来越受到关注[29,30]。但是,由于褐飞虱具有迁飞性,长期有效防治褐飞虱的方法仍然匮乏。【拟解决的关键问题】通常昆虫中仅有一个单独的TPS,TPS及TPP的功能研究却少有报道[17,31],本研究从褐飞虱新发现的第3个TPS入手,分析TPS3的序列结构特征,并研究褐飞虱不同TPS在各发育阶段的表达特征及TPS3在褐飞虱蜕皮和翅形成中潜在功能,为褐飞虱防治提供理论依据。1 材料与方法
试验于2016年9月至2017年7月在杭州师范大学完成。1.1 供试材料
供试褐飞虱采集自浙江省农业科学院,在笔者实验室饲养。供试褐飞虱和水稻均置于人工气候箱或人工气候室中饲养和培育。褐飞虱饲养条件:温度(26±1)℃,光周期16 h/8 h,相对湿度为70%。用于饲养褐飞虱的水稻品种为TN1(Taichung Native 1)。水稻种植步骤:首先浸种,将水稻种子浸入约70℃的温水中浸泡10 min左右,再将种子浸泡到自来水中放至30℃人工气候培养箱浸种24 h;随后倒去浸种用水,用自来水冲洗数次后用湿纱布包裹种子,置于30℃人工气候培养箱催芽24—48 h,种子发芽后播种于塑料盆;适当施肥促进小苗生长,待长至三叶一心,转移至大田插秧;待水稻生长至分蘖中期后将其移至养虫笼,每隔2—3 d更换一次水稻。大肠杆菌为笔者实验室保存,基因型为DH5α,质粒pMD18-T载体购自TaKaRa公司。
1.2 试验方法
1.2.1 褐飞虱总RNA的抽提及反转录成cDNA 取新鲜或存于-80℃冰箱的褐飞虱7头放入1.5 mL塑料离心管中,加Trizol 200 μL,用电动组织研磨器充分研磨,再加Trizol 800 μL补足至1 mL,用力振荡3 min,室温静置8 min;吸取200 μL氯仿加入塑料离心管中,上下颠倒直至混匀呈现均匀的乳粉色,室温静置10 min;4℃,12 000 r/min离心15 min;将上清液转移至1.5 mL塑料离心管,加入异丙醇500 μL,上下颠倒直至混匀,室温静置20 min; 4℃,12 000 r/min离心10 min;轻轻倒去上清,加入1 mL的75%乙醇,并吹打洗涤沉淀;4℃,7 500 r/min离心5 min;轻轻倒去上清,用移液器吸出多余液体后将离心 管倒置于超净台中3—5 min,根据沉淀量加入20—50 μL DEPC水,轻轻吹打至沉淀溶解;利用电泳及分光光度计评价总RNA的完整性,剩余总RNA存放于-80℃冰箱。使用TaKaRa PrimeScriptTM RT reagent Kit With gDNA Eraser试剂盒。先去除基因组DNA,方法为将RNase-free的500 μL塑料离心管置于冰上,加入5×gDNA Eraser Buffer 2 μL、gDNA Eraser 1 μL及Total RNA 1 μg,并用RNase Free dH2O补足至10 μL,水浴42℃反应2 min。随后在冰上向前述塑料离心管中加入5×PrimeScript Buffer 4 μL,PrimeScript RT Enzyme Mix I 1 μL及RT Primer Mix 1 μL,并用RNase Free dH2O补足至20 μL,水浴37℃反应15 min,水浴85℃反应5 s,cDNA保存于-20℃冰箱。
1.2.2 全长TPS3的PCR扩增及目的片段回收 使用TaKaRa Ex TaqTM试剂盒。向200 μL PCR管中加入褐飞虱cDNA 1 μL、Ex Taq 0.2 μL、10×Ex Taq Buffer(20 mmol·L-1 Mg2+ plus)2.5 μL、正向引物1 μL、反向引物1 μL、dNTP Mixture(2.5 mmol·L-1)2 μL,加无菌水补足至25 μL。将PCR产物扩增产物中取5 μL样品进行浓度为1%的琼脂糖凝胶电泳分析,采用Andy SafeTM核酸染料作为指示剂,检测到目的条带后将剩余PCR产物进行电泳,在紫外凝胶成像系统下用手术刀切割目的条带并放入1.5 mL塑料离心管。使用生工生物工程(上海)股份有限公司EZ Spin Column DNA Gel Extraction Kit纯化DNA。回收DNA,保存于-20℃冰箱。
1.2.3 回收产物的连接、转化、鉴定及测序 在冰上向200 μL PCR管中加入pMD 18-T Vector 0.5 μL、Solution I 3.5 μL及回收DNA片段3 μL,吹打混匀,水浴16℃反应30 min,若片段较长则延长反应时间。连接反应结束后,将连接片段转化到DH5α感受细胞中。等到平板长出菌落后,挑取生长均匀,菌落边缘光滑且颜色透明的菌落至20 μL无菌水,混匀,用作菌落PCR模板,用于验证连接转化是否成功,片段长度验证为正确的菌液制备成甘油菌送往上海Invitrogen公司测序。
1.2.4 TPS3蛋白序列及二级、三级结构分析 在获得TPS cDNA序列后,将其ORF翻译成蛋白质,使用BLAST序列比对软件(
利用SOPMA 在线分析软件(
1.2.5 不同发育阶段褐飞虱TPS的表达 从4龄褐飞虱开始,每12 h取材一次,每次随机取3组,每组7头,抽提总RNA,反转录成cDNA,用于通过实时荧光定量PCR(qRT-PCR)检测褐飞虱TPS1、TPS2及TPS3在不同发育时期的相对表达量。选用褐飞虱18S作为内参基因[36],使用Primer Premier 5.0软件设计褐飞虱TPS1、TPS2及TPS3的qRT-PCR引物(表1)。对引物进行梯度PCR验证,根据Tm值选择最适退火温度,并分析熔解曲线,进行琼脂糖凝胶电泳,确保PCR产物的特异性。
Table 1
表1
表1荧光实时定量PCR引物和TPS3及GFP的dsRNA引物序列
Table 1
| 基因名称 Primer name | 正向引物 Forward primer (5′-3′) | 反向引物 Reverse primer (5′-3′) |
|---|---|---|
| QNl18S QNlTPS1 QNlTPS2 QNlTPS3 | CGCTACTACCGATTGAA AAGACTGAGGCGAATGGT AGAGTGGACCGCAACAACA GTGATGCGTCGGTGGCTAT | GGAAACCTTGTTACGACTT AAGGTGGAAATGGAATGTG TCAACGCCGAGAATGACTT CCGTTCATCATTGGGCATAGT |
| dsNlTPS3 | GAGTCTGACCTGATAGCCTTTA | ATCGGAGTCCATTTAGTTGT |
| dsNlTPS3-T7 | GGATCCTAATACGACTCACTATAGGGAGTCTGACCTGATAGCCTTTA | GGATCCTAATACGACTCACTATAGGATCGGAGTCCATTTAGTTGT |
| dsNlGFP | AAGGGCGAGGAGCTGTTCACCG | CAGCAGGACCATGTGATCGCGC |
| dsNlGFP-T7 | GGATCCTAATACGACTCACTATAGGAAGGGCGAGGAGCTGTTCACCG | GGATCCTAATACGACTCACTATAGGCAGCAGGACCATGTGATCGCGC |
新窗口打开|下载CSV
qRT-PCR体系:SYBR Premix Ex Taq 10 μL、模板cDNA 1 μL(80 ng·μL-1)、PCR Forward primer(10 pmol)1 μL、PCR Reverse primer(10 pmol)1 μL,并用无菌水补足至20 μL。PCR扩增程序:95℃预变性3 min;95℃变性5 s,55—60℃退火延伸30 s,39个循环;65℃ 5 s,95℃绘制熔解曲线。
1.2.6 褐飞虱dsRNA注射及TPS基因表达量的测定 dsRNA的合成:设计合成GFP和褐飞虱TPS3的dsRNA引物(具体引物序列见表1)进行PCR扩增,扩增产物进行T克隆,然后用带T7启动子的引物进行交叉PCR反应,使用T7 RiboMAXTM Express RNAi System试剂盒合成dsRNA,最后通过NanoDropTM 2000型微量测定分光光度计检测合成的dsRNA的浓度。
dsRNA注射褐飞虱:选取5龄第1天褐飞虱若虫为试验对象,进行dsTPS3注射,同时注射dsGFP作为对照组。将dsRNA从褐飞虱胸部两对后足中间注入体腔,注射量为200 ng/头,设置3个生物学重复。将注射好的褐飞虱转移至装有新鲜水稻的玻璃管中饲养,在注射后48 h收取褐飞虱活体,用液氮速冻后存于-80℃备用。
TPS表达情况的测定:取注射后48 h收取的褐飞虱活体样本抽提总RNA,然后反转录得到3管平行cDNA,参照SYBR? Premix Ex TaqTM试剂盒的方法进
行qRT-PCR检测。
1.2.7 数据统计与分析 通过定量PCR测定出4个基因的CT值,每管样品的3个重复孔取平均值用于计算(数值间变异系数应<5%,标准差应<0.5,3个数值中有2个或3个符合,数值可用,否者重做)。每次样品有3组数值,即最后得到的数据为平均值±9个重复。再通过2-ΔΔCT法进行计算,对照组为褐飞虱dsGFP注射组的CT值。最后将换算出的值再进行具体分析。2-ΔΔCT计算公式[18]:2-ΔΔCT= 2-[(CT对照组-CT对照18s)- (CT待测组-CT待测组18s)]。
采用SPSS数据分析软件One-Way ANOVA进行差异性分析,再使用SigmaPlot 10.0软件的Vertical Bar Chart类型中的Grouped Error Bars图表选项绘制柱状图。
2 结果
2.1 褐飞虱TPS3序列全长及蛋白特性分析
褐飞虱TPS3的开放阅读框长度为2 352 bp,翻译成蛋白质的氨基酸数目为783个,预测的蛋白分子量为88.9 kD(C3978H6237N1063O1163S42),等电点为5.47,具有亲水性(表2、图1)。TPS3蛋白分析发现,含有20种氨基酸,其中亮氨酸(Leu)、异亮氨酸(Ile)、丝氨酸(Ser)和缬氨酸(Val)等含量较丰富,所占比例分别为10.0%、7.5%、7.4%和7.4%。酸性氨基酸(带负电荷)为101个,碱性氨基酸(带正电荷)为83个。与之前克隆的两个TPS相比,本文克隆的第3个TPS与前两个TPS具有相似的氨基酸数量,等电点都偏酸性。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1褐飞虱TPS3的核苷酸和翻译的氨基酸序列
Fig. 1Nucleotide and amino acid sequences of N. lugens TPS3
2.2 昆虫TPS氨基酸序列同源分析
选择德国小蠊的TPS1和TPS2、黑腹果蝇的TPS1、埃及伊蚊的TPS、赤拟谷盗的TPS以及家蚕的TPS与褐飞虱的TPS1、TPS2、TPS3蛋白进行同源比对,发现与其他昆虫的TPS蛋白相比,褐飞虱的3条TPS蛋白具有较高的氨基酸序列相似性。同时,包括褐飞虱在内的昆虫TPS蛋白都具有TPS和TPP这两个保守结构域,分别对应着酵母的OtsA和OtsB基因。此外,昆虫TPS还包含着一些非常保守的特征序列,如“WPLFHSMP”、“CRVDR”、“VDRLD”、“GRING”、“TPLRDGMN”和“YDGTL”(图2)。依据褐飞虱已经发表的基因组序列[28],分析发现TPS1(GQ397450)、TPS2(KU556826)和TPS3(KU556827)分别定位在scaffolds3259、scaffolds2574及scaffolds80上。上结果表明,新发现的TPS3确实为一种新的TPS蛋白。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2褐飞虱的3条TPS与其他昆虫同源蛋白的氨基酸序列比对
Fig. 2Amino acid sequence alignment of three TPSs from N. lugens with homogeneous proteins from other insects
Table 2
表2
表2褐飞虱3个TPS蛋白的部分基本理化性质
Table 2
| 蛋白名称 Protein name | 氨基酸数 Amino acid number | 相对分子量 Relative molecular weight | 等电点 Isoelectric point | 亲水性 Hydrophily |
|---|---|---|---|---|
| TPS1 | 807 | 90536 | 6.14 | -0.180 |
| TPS2 | 820 | 93215 | 6.07 | -0.190 |
| TPS3 | 783 | 88909 | 5.47 | -0.126 |
新窗口打开|下载CSV
2.3 褐飞虱3个TPS二级结构预测及分析
TPS比较保守,在二级结构上TPS1、TPS2和TPS3有明显的相似性(图3),特别是TPS1和TPS2,它们在相似的位点具有α-螺旋或β-折叠结构。具体表现在TPS1、TPS2和TPS3的α-螺旋所占比例分别为40.27%、36.59%和35.50%;β-折叠所占比例分别为12.76%、13.29%和18.01%;而无规则卷曲所占的比例分别为45.35%、49.39%和46.49%(表3)[13]。
图3
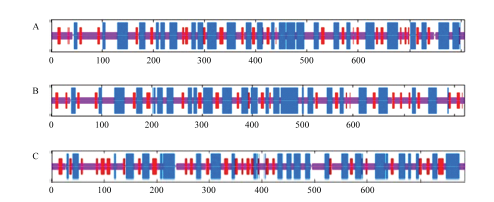 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3TPS1、TPS2和TPS3的二级结构预测
A: TPS1; B: TPS2; C: TPS3。蓝色代表α-螺旋,红色代表β-折叠[31] The blue represents α-helix and the red represents β-sheet
Fig. 3The secondary structure prediction of TPS1, TPS2 and TPS3
Table 3
表3
表3褐飞虱TPS二级结构预测
Table 3
| α-螺旋 α-helix (%) | β-折叠 β-sheet (%) | 无规则卷曲 Random coil (%) | |
|---|---|---|---|
| TPS1 | 40.27 | 12.76 | 45.35 |
| TPS2 | 36.59 | 13.29 | 49.39 |
| TPS3 | 35.50 | 18.01 | 46.49 |
新窗口打开|下载CSV
2.4 蛋白质三维结构预测
由图4可以看出,TPS1的3D结构的寡聚物(oligostate)类型为同型二聚体(homodimer),含有较多的α-螺旋结构,建模残基范围(modelled residue range)为6—416,基于模型5hxa.1.A建立,序列同源性(sequence identity)为37.91%。TPS2的3D结构的寡聚物类型为同型四聚体(homotetramer),由α-螺旋及β-折叠盘绕形成,并通过无规则卷曲相连,建模残基范围为8—492,基于模型5tvg.1.A建立,序列同源性为36.75%。TPS3的3D结构的寡聚物类型为单体(monomer),含有大量α-螺旋及β-折叠,建模残基范围为13—494。图4
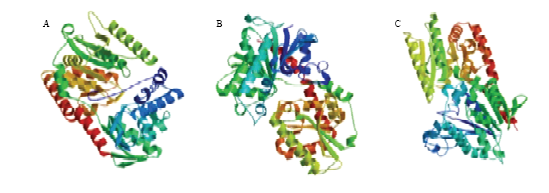 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4褐飞虱3个TPS蛋白质三维结构的预测
Fig. 4The predicted 3D structure of three TPSs from N. lugens
A: TPS1; B: TPS2; C: TPS3
2.5 褐飞虱TPS1、TPS2及TPS3不同发育阶段表达
由图5可以看出,TPS1和TPS2在褐飞虱4龄阶段表达量相对比较低,并且TPS1从4龄及5龄若虫阶段相对表达量呈现逐步上升到趋于平缓,在成虫阶段相对表达量明显上升,在成虫48 h开始相对表达量呈现极大的上升,在成虫60 h达到最大(图5-A)。TPS2在褐飞虱4龄36 h表达量明显上升,整个5龄若虫阶段都有相对较高的表达,在5龄96 h开始逐渐降低,在成虫36 h后开始回升(图5-B)。TPS3在褐飞虱4龄、5龄若虫乃至成虫阶段都有较高的相对表达量,其中5龄72 h开始较此之前的相对表达量有所提高(图5-C)。图5
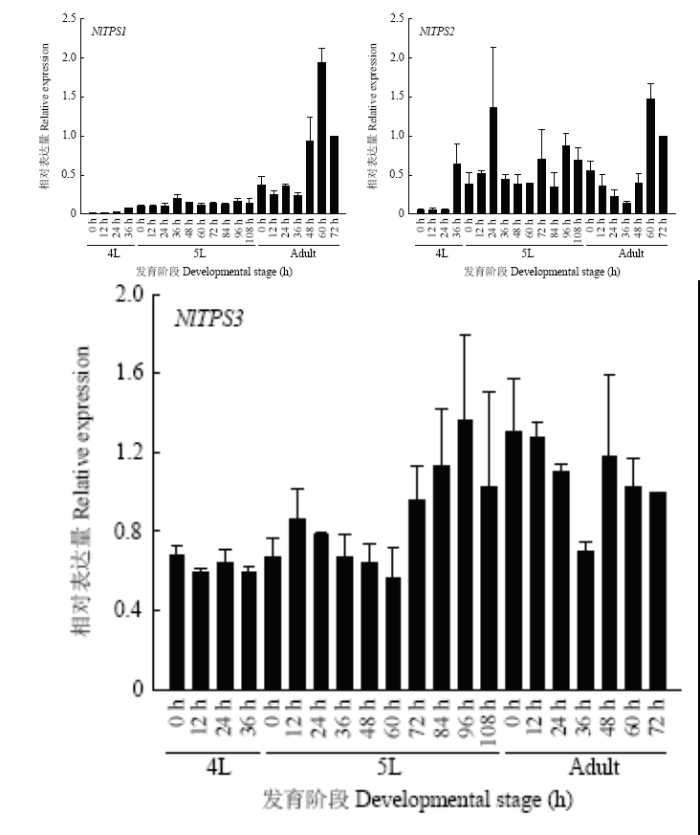 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5褐飞虱3条TPS发育表达情况
羽化后72 h的表达量设为1
Fig. 5Relative expression of three TPS genes in different developmental stages of N. lugens
The expression level at 72 h after emergence is set to 1
2.6 褐飞虱TPS3 dsRNA注射沉默效果
单独干扰褐飞虱TPS3 48 h时,TPS3的表达显著下降(P<0.01),说明单独注射TPS3可以有效抑制本基因的表达(图6-A)。同时,与dsGFP组相比较,dsTPS3注射后,部分褐飞虱的蜕皮过程出现障碍,表现为蜕皮不能顺利完成及翅发育出现畸形(图6-B)。图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6注射48 h后褐飞虱TPS3的表达(A)及褐飞虱蜕皮和翅发育(B)情况
**:差异极显著(P<0.01)Extremely significant difference
Fig. 6Relative expression of TPS3 after injection for 48 h (A) and molting and wing development (B) of N. lugens
3 讨论
昆虫TPS具有很强的保守性,一般包含约800个氨基酸,相对分子量在90 kD左右,等电点在5—8[3,37]。褐飞虱的TPS1、TPS2以及本研究新发现的TPS3分别包含807、820及783个氨基酸,预测出的相对分子量相似,且均为偏酸性等电点。尽管先前的研究表明昆虫海藻糖合成酶应该包含TPS和TPP两个合成酶基因。然而,TPS均有发现和报道,如东亚飞蝗、白背飞虱、猫栉首蚤(Ctenocephalus felis)、黑腹果蝇等[12,38-39];而仅在葱蝇(Delia antiqua)等个别物种中报道含有TPS和TPP[40,41];单独的TPP基因序列自2011年后在NCBI中有登录,如黑腹果蝇(NM_135269)、台湾乳白蚁(Coptotermes formosanus,JN565075)和小菜蛾(Plutella xylostella,XM_011559193),但TPP的功能研究少有报道[17,31]。同样,过去通常认为昆虫体内只有1个TPS,但在2014年赵丽娜[13]在褐飞虱中发现和克隆到了第2个TPS,随后在德国小蠊中也发现2个TPS[14]。在一些无脊椎动物中,多个TPS早有发现,如同属节肢动物门的蓝蟹(Callinectes sapidus)体内具有5个TPS,并且长度各不相同,但有部分序列重叠[42];一种蛔虫(Ascaris suum)中同样发现和报道了3个TPS[31,43]。本研究在褐飞虱中发现并获得了第3个TPS,通过比较发现TPS1、TPS2、TPS3的基本理化性质和二级结构比较相似,但TPS3的三级结构与TPS1、TPS2相比差异较大,这可能与其具体的功能有关。目前比较认可的海藻糖合成途径为海藻糖在昆虫体内首先由UDP-葡萄糖和葡萄糖-6-磷酸通过海藻糖-6-磷酸合成酶(TPS)合成为海藻糖-6-磷酸,随后海藻糖-6-磷酸酯酶(TPP)将其转变为海藻糖[10,11]。唐斌等于2018年提出一种潜在的可能途径,即某些昆虫或无脊椎物种中,海藻糖合成酶可以单独将UDP-葡萄糖和葡萄糖-6-磷酸转化为海藻糖[10,31]。支持这种途径的依据有两个:一是昆虫所有发现的TPS蛋白都包含了TPS和TPP保守结构域,与酵母中OtsA和OtsB相对应,而OtsA和OtsB在酵母中分别为合成海藻糖的两个关键基因;其二,将棉铃虫的TPS体外表达,发现其具有单独合成海藻糖的能力[44],同样在果蝇中也有类似的研究结果[45]。当然,这种潜在海藻糖合成途径还需要更多的科学研究进行验证,而包括褐飞虱在内的多个TPS的具体功能及功能差异性研究将有助于推动昆虫海藻糖合成途径进化及多样性研究的发展。
基因的表达与其功能行使有着非常密切的关系。前期研究发现蓝蟹血淋巴中的TPS在预蜕皮期及蜕皮前期有较高的相对表达量,并且在蜕皮后1—7 d也有一定的表达[46]。在葱蝇中发现TPS在预蛹期也有较高的相对表达量[41]。褐飞虱为一种不完全变态昆虫,不同阶段发育表达研究表明TPS1在5龄若虫蜕皮为成虫后表达量上升,TPS2在4龄和5龄末的蜕皮前表达量较高,TPS3则在整个4龄若虫到成虫阶段表达量都较高,这些结果表明褐飞虱在蜕皮阶段和成虫期需要大量的海藻糖,而且可能不同的TPS潜在功能有所不同。相似的研究发现果蝇幼虫阶段体内海藻糖含量较高,预蛹阶段的海藻糖含量达到最高,但在蛹期迅速回落[47]。同样的在4龄若虫和5龄若虫蜕皮前褐飞虱体内的海藻糖含量均较高[48],不同发育阶段的糖原含量变化也出现了这种情况,说明褐飞虱蜕皮生长时需要大量消耗储存的海藻糖和糖原。图5的结果表明褐飞虱3个TPS在不同发育阶段相对表达量的变化趋势不尽相同,这从某种意义上说明3个TPS在褐飞虱生长发育过程中可能具有不同的功能。先前的研究发现TPS1和TPS2都能通过调控几丁质代谢使得几丁质含量降低,从而控制褐飞虱的蜕皮[13,17],本研究同样发现TPS3表达被抑制后出现蜕皮困难与翅发育畸形现象,这些结果与TPS1和TPS2的功能类似,但是TPS3是否同样具有调控几丁质合成或其他的功能,则需要更加深入的研究。
4 结论
TPS3属于一个新TPS家族基因,其与褐飞虱中已经报道的TPS1和TPS2具有较高的同源性,包含TPS和TPP两个保守结构域,3个TPS具有相似的氨基酸组成。不同发育阶段表达结果表明,3个TPS在发育过程中行使的功能不同,dsTPS3注射能够有效抑制TPS3的表达,同时导致部分褐飞虱翅畸形和蜕皮出现障碍。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
DOI:10.1093/glycob/cwg047URLPMID:12626396 [本文引用: 1]

ABSTRACT Trehalose is a nonreducing disaccharide in which the two glucose units are linked in an alpha,alpha-1,1-glycosidic linkage. This sugar is present in a wide variety of organisms, including bacteria, yeast, fungi, insects, invertebrates, and lower and higher plants, where it may serve as a source of energy and carbon. In yeast and plants, it may also serve as a signaling molecule to direct or control certain metabolic pathways or even to affect growth. In addition, it has been shown that trehalose can protect proteins and cellular membranes from inactivation or denaturation caused by a variety of stress conditions, including desiccation, dehydration, heat, cold, and oxidation. Finally, in mycobacteria and corynebacteria, trehalose is an integral component of various glycolipids that are important cell wall structures. There are now at least three different pathways described for the biosynthesis of trehalose. The best known and most widely distributed pathway involves the transfer of glucose from UDP-glucose (or GDP-glucose in some cases) to glucose 6-phosphate to form trehalose-6-phosphate and UDP. This reaction is catalyzed by the trehalose-P synthase (TPS here, or OtsA in Escherichia coli ). Organisms that use this pathway usually also have a trehalose-P phosphatase (TPP here, or OtsB in E. coli) that converts the trehalose-P to free trehalose. A second pathway that has been reported in a few unusual bacteria involves the intramolecular rearrangement of maltose (glucosyl-alpha1,4-glucopyranoside) to convert the 1,4-linkage to the 1,1-bond of trehalose. This reaction is catalyzed by the enzyme called trehalose synthase and gives rise to free trehalose as the initial product. A third pathway involves several different enzymes, the first of which rearranges the glucose at the reducing end of a glycogen chain to convert the alpha1,4-linkage to an alpha,alpha1,1-bond. A second enzyme then releases the trehalose disaccharide from the reducing end of the glycogen molecule. Finally, in mushrooms there is a trehalose phosphorylase that catalyzes the phosphorolysis of trehalose to produce glucose-1-phosphate and glucose. This reaction is reversible in vitro and could theoretically give rise to trehalose from glucose-1-P and glucose. Another important enzyme in trehalose metabolism is trehalase (T), which may be involved in energy metabolism and also have a regulatory role in controlling the levels of trehalose in cells. This enzyme may be important in lowering trehalose concentrations once the stress is alleviated. Recent studies in yeast indicate that the enzymes involved in trehalose synthesis (TPS, TPP) exist together in a complex that is highly regulated at the activity level as well as at the genetic level.
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/192680a0URL [本文引用: 1]
ALMOST two decades have elapsed since Abramowitz et al. 1 first reported the occurrence of a ‘diabetogenic’ factor in the sinus gland of a crustacean. To the best of my knowledge the experiments described here are the first to show that a substance with a similar effect occurs in the corpus cardiacum of an insect. The corpus cardiacum was selected for investigation because of its analogy to the sinus glands of Crustacea and its known content of biologically active materials 2–4 . In view of the recent finding that the main blood sugar in insects is the non-reducing disaccharide trehalose 5–7 the effect of corpora cardiaca extracts on both reducing and non-reducing sugars in the blood has been investigated.
DOI:10.1038/198685a0URL [本文引用: 1]
HUMORAL involvement in the carbohydrate metabolism of Periplaneta americana (L.) has been reported by Steele 1 , who demonstrated a 150 per cent increase in blood trehalose and a drop in blood glucose following injection of an aqueous extract of the corpus cardiacum. The work recorded here, conducted as part of an overall investigation of the regulation of sugar utilization in the South American cockroach, Blaberus discoidalis (Serville), confirms the rise in trehalose, but reveals also a significant increase in blood glucose and a concomitant decrease in fat body glycogen.
[D].
[本文引用: 3]
[本文引用: 3]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
.
[本文引用: 2]
[D].
[本文引用: 3]
[D].
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.ibmb.2016.07.012URLPMID:27524277
61Knockdown of trehalose-6-phosphate synthase (LdTPS) decreased but silencing of trehalase genes (LdTRE1aorLdTRE2) increased trehalose contents inLeptinotarsa decemlineatalarvae.61TheLdTPSRNAi survivors consumed more but theLdTRE1aRNAi hypomorphs fed less foliage.61TheLdTPSRNAi larvae were fat in contrast to theLdTRE1aRNAi hypomorphs.61LdTPSandLdTRE1aRNAi reduced chitin contents.61LdTPS,LdTRE1aandLdTRE2RNAi caused lethality.
DOI:10.1002/ps.4287URLPMID:27060284 [本文引用: 3]
Abstract BACKGROUND: RNA interference combined with digital gene expression (DGE) analysis can be used to study gene function. Trehalose-6-phosphate synthase (TPS) plays a key role in the synthesis of trehalose and insect development. RESULTS: DGE analysis revealed that the expression of nine or four chitinase genes was reduced significantly 48 h after NlTPS1 and NlTPS2 knockdown by RNAi, respectively. Additionally, abnormal phenotypes were noted, and approximately 30% of insects died. HK and G6PI2 expression decreased significantly whereas GFAT, GNPNA and UAP expression increased significantly 72 h after NlTPS1 and NlTPS2 knockdown. PGM1 expression decreased significantly after TPS2 knockdown, whereas PGM2 expression increased significantly and the expression of three CHS genes decreased 48 h after TPS1 knockdown. The mRNA expression of all 12 chitin degradation genes decreased 48 h after NlTPS1 and NlTPS2 treatment, and Cht2, Cht3, Cht6, Cht7, Cht10 and ENGase levels remained significantly decreased up to 72 h after NlTPS1 and NlTPS2 knockdown. CONCLUSIONS: These results demonstrate that silencing of TPS genes can lead to increased moulting deformities and mortality rates owing to the misregulation of genes involved in chitin metabolism, and TPS genes are potential pest control targets in the future. 2016 Society of Chemical Industry. 2016 Society of Chemical Industry.
DOI:10.1017/S000748531700089XURLPMID:28920565 [本文引用: 2]
Abstract RNA interference (RNAi) is a very effective technique for studying gene function and may be an efficient method for controlling pests. Trehalose-6-phosphate synthase (TPS), which plays a key role in the synthesis of trehalose and insect development, was cloned in Tribolium castaneum (Herbst) (TcTPS) and the putative functions were studied using RNAi via the injection of double-stranded RNA (dsRNA) corresponding to conserved TPS and trehalose-6-phosphate phosphatase domains. Expression analyses show that TcTPS is expressed higher in the fat body, while quantitative real-time polymerase chain reaction results show that the expression of four trehalase isoforms was significantly suppressed by dsTPS injection. Additionally, the expression of six chitin synthesis-related genes, such as hexokinase 2 and glutamine-fructose-6-phosphate aminotransferase, was suppressed at 48 and 72 h post-dsTPS-1 and dsTPS-2 RNA injection, which were two dsTPS fragments that had been designed for two different locations in TcTPS open reading frame, and that trehalose content and trehalase 1 activity decreased significantly at 72 h post-dsRNA injection. Furthermore, T. castaneum injected with dsTPS-1 and dsTPS-2 RNA displayed significantly lower levels of chitin and could not complete the molting process from larvae to pupae, revealing abnormal molting phenotypes. These results demonstrate that silencing TPS gene leads to molting deformities and high mortality rates via regulation of gene expression in the chitin biosynthetic pathway, and may be a promising approach for pest control in the future.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
DOI:10.1186/s13059-014-0521-0URLPMID:4269174 [本文引用: 2]
Abstract BACKGROUND: The brown planthopper, Nilaparvata lugens, the most destructive pest of rice, is a typical monophagous herbivore that feeds exclusively on rice sap, which migrates over long distances. Outbreaks of it have re-occurred approximately every three years in Asia. It has also been used as a model system for ecological studies and for developing effective pest management. To better understand how a monophagous sap-sucking arthropod herbivore has adapted to its exclusive host selection and to provide insights to improve pest control, we analyzed the genomes of the brown planthopper and its two endosymbionts. RESULTS: We describe the 1.14 gigabase planthopper draft genome and the genomes of two microbial endosymbionts that permit the planthopper to forage exclusively on rice fields. Only 40.8% of the 27,571 identified Nilaparvata protein coding genes have detectable shared homology with the proteomes of the other 14 arthropods included in this study, reflecting large-scale gene losses including in evolutionarily conserved gene families and biochemical pathways. These unique genomic features are functionally associated with the animal's exclusive plant host selection. Genes missing from the insect in conserved biochemical pathways that are essential for its survival on the nutritionally imbalanced sap diet are present in the genomes of its microbial endosymbionts, which have evolved to complement the mutualistic nutritional needs of the host. CONCLUSIONS: Our study reveals a series of complex adaptations of the brown planthopper involving a variety of biological processes, that result in its highly destructive impact on the exclusive host rice. All these findings highlight potential directions for effective pest control of the planthopper.
DOI:10.1038/nature14286URLPMID:25799997 [本文引用: 1]
Abstract Wing polyphenism is an evolutionarily successful feature found in a wide range of insects. Long-winged morphs can fly, which allows them to escape adverse habitats and track changing resources, whereas short-winged morphs are flightless, but usually possess higher fecundity than the winged morphs. Studies on aphids, crickets and planthoppers have revealed that alternative wing morphs develop in response to various environmental cues, and that the response to these cues may be mediated by developmental hormones, although research in this area has yielded equivocal and conflicting results about exactly which hormones are involved. As it stands, the molecular mechanism underlying wing morph determination in insects has remained elusive. Here we show that two insulin receptors in the migratory brown planthopper Nilaparvata lugens, InR1 and InR2, have opposing roles in controlling long wing versus short wing development by regulating the activity of the forkhead transcription factor Foxo. InR1, acting via the phosphatidylinositol-3-OH kinase (PI(3)K)-protein kinase B (Akt) signalling cascade, leads to the long-winged morph if active and the short-winged morph if inactive. InR2, by contrast, functions as a negative regulator of the InR1-PI(3)K-Akt pathway: suppression of InR2 results in development of the long-winged morph. The brain-secreted ligand Ilp3 triggers development of long-winged morphs. Our findings provide the first evidence of a molecular basis for the regulation of wing polyphenism in insects, and they are also the first demonstration--to our knowledge--of binary control over alternative developmental outcomes, and thus deepen our understanding of the development and evolution of phenotypic plasticity.
DOI:10.1073/pnas.1721473115URL [本文引用: 1]
Variation in food nutrient content and density are key ecological factors linked to the expression of condition-dependent, adaptive phenotypes such as wing polyphenisms. There is very little known about exactly what the ecological cue is that induces the appropriate insect phenotype in wing polyphenic insects. Our study reveals that glucose concentration of the host plant and insect density directly influence the development of brown planthoppers into either the long-winged migratory morph or the short-winged reproductive morph. This study is a step in linking host quality signals and other factors such as density to the induction of adaptive phenotypes in insects. Food quality is a critical environmental condition that impacts an animal growth and development. Many insects facing this challenge have evolved a phenotypically plastic, adaptive response. For example, many species of insect exhibit facultative wing growth, which reflects a physiological and evolutionary trade-off between dispersal and reproduction, triggered by environmental conditions. What the environmental cues are and how they are transduced to produce these alternative forms, and their associated ecological shift from dispersal to reproduction, remains an important unsolved problem in evolutionary ecology. In this study, we investigated the role that host quality has on the induction of wing development in a wing polyphenic insect exhibiting strong tradeoffs in investment between dispersal and reproduction, the brown planthopper, a serious rice pest in Asia. As rice plants grow, the short-winged brown planthopper dominates the population, but a shift occurs as the plants mature and senesce in the field such that long-winged brown planthoppers emerge and migrate. It remains unknown how changes in the rice plant induce development of the long-winged morph, despite recent discoveries on the role of the insulin and JNK signaling pathways in wing development. We found that by mimicking the glucose concentration of senescing rice plants, we significantly increased the proportion of long-winged female planthoppers. The effects of glucose on wing morph are additive with previously described effects of density. Our results show that host quality both directly regulates phenotypic plasticity and interacts with other factors such as density to produce the appropriate phenotype for specific environmental conditions.
DOI:10.3389/fphys.2018.00030URLPMID:5797772 [本文引用: 6]
The non-reducing disaccharide trehalose is widely distributed among various organisms. It plays a crucial role as an instant source of energy, being the major blood sugar in insects. In addition, it helps countering abiotic stresses. Trehalose synthesis in insects and other invertebrates is thought to occur via the trehalose-6-phosphate synthase (TPS) and trehalose-6-phosphate phosphatase (TPP) pathways. In many insects, theTPPgene has not been identified, whereas multipleTPSgenes that encode proteins harboring TPS/OtsA and TPP/OtsB conserved domains have been found and cloned in the same species. The function of theTPSgene in insects and other invertebrates has not been reviewed in depth, and the available information is quite fragmented. The present review discusses the current understanding of the trehalose synthesis pathway, TPS genetic architecture, biochemistry, physiological function, and potential sensitivity to insecticides. We note the variability in the number ofTPSgenes in different invertebrate species, consider whether trehalose synthesis may rely only on theTPSgene, and discuss the results ofin vitro TPSoverexpression experiment. Tissue expression profile and developmental characteristics of theTPSgene indicate that it is important in energy production, growth and development, metamorphosis, stress recovery, chitin synthesis, insect flight, and other biological processes. We highlight the molecular and biochemical properties of insect TPS that make it a suitable target of potential pest control inhibitors. The application of trehalose synthesis inhibitors is a promising direction in insect pest control because vertebrates do not synthesize trehalose; therefore, TPS inhibitors would be relatively safe for humans and higher animals, making them ideal insecticidal agents without off-target effects.
DOI:10.1093/bioinformatics/bti770URLPMID:16301204 [本文引用: 1]
Homology models of proteins are of great interest for planning and analysing biological experiments when no experimental three-dimensional structures are available. Building homology models requires specialized programs and up-to-date sequence and structural databases. Integrating all required tools, programs and databases into a single web-based workspace facilitates access to homology modelling from a computer with web connection without the need of downloading and installing large program packages and databases. SWISS-MODEL workspace is a web-based integrated service dedicated to protein structure homology modelling. It assists and guides the user in building protein homology models at different levels of complexity. A personal working environment is provided for each user where several modelling projects can be carried out in parallel. Protein sequence and structure databases necessary for modelling are accessible from the workspace and are updated in regular intervals. Tools for template selection, model building and structure quality evaluation can be invoked from within the workspace. Workflow and usage of the workspace are illustrated by modelling human Cyclin A1 and human Transmembrane Protease 3. The SWISS-MODEL workspace can be accessed freely at http://swissmodel.expasy.org/workspace/
[本文引用: 1]
DOI:10.1093/bioinformatics/btq662URLPMID:21134891Magsci [本文引用: 1]
Abstract MOTIVATION: Quality assessment of protein structures is an important part of experimental structure validation and plays a crucial role in protein structure prediction, where the predicted models may contain substantial errors. Most current scoring functions are primarily designed to rank alternative models of the same sequence supporting model selection, whereas the prediction of the absolute quality of an individual protein model has received little attention in the field. However, reliable absolute quality estimates are crucial to assess the suitability of a model for specific biomedical applications. RESULTS: In this work, we present a new absolute measure for the quality of protein models, which provides an estimate of the 'degree of nativeness' of the structural features observed in a model and describes the likelihood that a given model is of comparable quality to experimental structures. Model quality estimates based on the QMEAN scoring function were normalized with respect to the number of interactions. The resulting scoring function is independent of the size of the protein and may therefore be used to assess both monomers and entire oligomeric assemblies. Model quality scores for individual models are then expressed as 'Z-scores' in comparison to scores obtained for high-resolution crystal structures. We demonstrate the ability of the newly introduced QMEAN Z-score to detect experimentally solved protein structures containing significant errors, as well as to evaluate theoretical protein models. In a comprehensive QMEAN Z-score analysis of all experimental structures in the PDB, membrane proteins accumulate on one side of the score spectrum and thermostable proteins on the other. Proteins from the thermophilic organism Thermatoga maritima received significantly higher QMEAN Z-scores in a pairwise comparison with their homologous mesophilic counterparts, underlining the significance of the QMEAN Z-score as an estimate of protein stability. AVAILABILITY: The Z-score calculation has been integrated in the QMEAN server available at: http://swissmodel.expasy.org/qmean.
DOI:10.1093/nar/gku340URLPMID:24782522 [本文引用: 1]
Protein structure homology modelling has become a routine technique to generate 3D models for proteins when experimental structures are not available. Fully automated servers such as SWISS-MODEL with user-friendly web interfaces generate reliable models without the need for complex software packages or downloading large databases. Here, we describe the latest version of the SWISS-MODEL expert system for protein structure modelling. The SWISS-MODEL template library provides annotation of quaternary structure and essential ligands and co-factors to allow for building of complete structural models, including their oligomeric structure. The improved SWISS-MODEL pipeline makes extensive use of model quality estimation for selection of the most suitable templates and provides estimates of the expected accuracy of the resulting models. The accuracy of the models generated by SWISS-MODEL is continuously evaluated by the CAMEO system. The new web site allows users to interactively search for templates, cluster them by sequence similarity, structurally compare alternative templates and select the ones to be used for model building. In cases where multiple alternative template structures are available for a protein of interest, a user-guided template selection step allows building models in different functional states. SWISS-MODEL is available at http://swissmodel.expasy.org/.
DOI:10.1038/srep27841URLPMID:27328657 [本文引用: 1]
RNA interference (RNAi) is an effective gene-silencing tool, and double stranded RNA (dsRNA) is considered a powerful strategy for gene function studies in insects. In the present study, we aimed to investigate the function of trehalase (TRE) genes (TRE 1-1,TRE 1-2, andTRE-2) isolated from the brown planthopperNilaparvata lugens, a typical piercing-sucking insect in rice, and investigate their regulating roles in chitin synthesis by injecting larvae with dsRNA. The results showed thatTRE1andTRE2had compensatory function, and the expression of each increased when the other was silenced. The total rate of insects with phenotypic deformities ranged from 19.83 to 24.36% after dsTRE injection, whereas the mortality rate ranged from 14.16 to 31.78%. The mRNA levels of genes involved in the chitin metabolism pathway in RNA-Seq and DGEP, namely hexokinase (HK), glucose-6-phosphate isomerase (G6PI) and chitinase (Cht), decreased significantly at 72 after single dsTREs injection, whereas two transcripts of chitin synthase (CHS) genes decreased at 72 after dsTRE1-1 and dsTREs injection. These results demonstrated thatTREsilencing could affect the regulation of chitin biosynthesis and degradation, causing moulting deformities. Therefore, expression inhibitors ofTREs might be effective tools for the control of planthoppers in rice.
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.gene.2015.03.070URLPMID:25841989 [本文引用: 2]
61TPS, TPP and TRE involving in trehalose metabolism were cloned and characterized.61Their enzymatic activities were determined in diapausing Dalia antiqua pupae.61They share a similar expression pattern in summer and winter diapausing pupae.61Glucose and trehalose contents are different between both diapausing pupae.61Trehalose's function was suggested in cold hardiness and desiccation resistance.
[本文引用: 1]
DOI:10.1155/2014/936560URLPMID:4227463 [本文引用: 1]
The in vitro effect of ivermectin lethal dose on the activity of trehalose-6-phosphate synthase (TPS) and phosphatase (TPP) and the expression of their mRNA (tps1, tps2, and tpp genes) in the muscle of adult female Ascaris suum was investigated. The presence of ivermectin in the medium caused a decrease in TPS and TPP activities during the experiment compared with the start and control groups. The exception was the group of worms grown for 8 hours in a IVM solution, in which there was a little higher TPS activity than in the control. Real-time qPCR analysis showed reduced expression of tps1 and tps2, and unchanged tpp expression after 20 hours of incubation relative to the expression at time zero. Relative to the appropriate control groups, the expression of tps2 gene was slight increased but the other two genes were reduced after 8-hours of IVM-treatment. Then the expression of all three genes was lower at the end of cultivation. The level of gene expression was positively correlated with the activity of specific enzymes. In the case of tpp gene there was only a weak correlation. Prolonged exposure to ivermectin was effective in lowering TPS and TPP activity and their mRNA expression. However, the drug did not block the pathway.
DOI:10.1093/glycob/cwn127URLPMID:19004876 [本文引用: 1]
Abstract Trehalose and trehalose metabolism are crucial for insect development. We measured the content of polyhydric compounds in the hemolymph of diapause- and nondiapause-destined individuals of Helicoverpa armigera. We found that the trehalose content is much higher in diapause-destined individuals than that in nondiapause individuals. The activity of trehalose-6-phosphate synthase (TPS) during H. armigera larval-pupal development is significantly higher in diapause-type individuals and is closely correlated with the changes in the trehalose content. The cDNA encoding TPS, which converts uridine-5'-diphosphoglucose and glucose-6-phosphate to trehalose-6-phosphate, was cloned from the fat body of H. armigera using rapid amplification of cDNA ends (RACE). The molecular characterization of the cDNA revealed that the mRNA encodes a precursor polypeptide of 826-amino-acid residues, containing Har-TPS at residues 6-507 and a putative trehalose-6-phosphate phosphatase, which converts trehalose-6-phosphate into free trehalose, at residues 512-783. The Har-TPS precursor polypeptide shows 73% identity with that of Drosophila melanogaster. The presence of a 2.8 kb transcript in the fat body and ovary was detected with a northern blot. The Har-TPS mRNA was detected at high levels in the late stage of sixth larval instar and the early middle stage of diapause-destined pupae, which are most likely to respond the changes in TPS activity and trehalose in the hemolymph. The Har-TPS protein was successfully overexpressed in the Bombyx mori baculovirus expression system, and the catalytic activity of Har-TPS was found to be approximately 5-fold higher in B. mori blood infected by the recombined-baculovirus than the control. When diapause is broken, the trehalose content drops significantly and glucose increases rapidly. These results suggest that trehalose is involved in regulating H. armigera pupal diapause.
DOI:10.1038/srep30582URLPMID:27469628 [本文引用: 1]
In insects, trehalose serves as the main sugar component of haemolymph. Trehalose is also recognized as a mediator of desiccation survival due to its proposed ability to stabilize membranes and proteins. Although the physiological role of trehalose in insects has been documented for decades, genetic evidence to support the importance of trehalose metabolism remains incomplete. We here show on the basis of genetic and biochemical evidence that the trehalose synthesis enzyme Tps1 is solely responsible for thede novosynthesis of trehalose inDrosophila. Conversely, a lack of the gene for the trehalose hydrolyzing enzymeTrehcauses an accumulation of trehalose that is lethal during the pupal period, as is observed withTps1mutants. Lack of eitherTps1orTrehresults in a significant reduction in circulating glucose, suggesting that the maintenance of glucose levels requires a continuous turnover of trehalose. Furthermore, changes in trehalose levels are positively correlated with the haemolymph water volume. In addition, bothTps1andTrehmutant larvae exhibit a high lethality after desiccation stress. These results demonstrate that the regulation of trehalose metabolism is essential for normal development, body water homeostasis, and desiccation tolerance inDrosophila.
DOI:10.1186/1746-1448-4-18URLPMID:19077285 [本文引用: 1]
Trehalose in ectoderms functions in energy metabolism and protection in extreme environmental conditions. We structurally characterized trehalose 6-phosphate synthase (TPS) from hemocytes of the blue crab, Callinectes sapidus. C. sapidus Hemo TPS (CasHemoTPS), like insect TPS, encodes both TPS and trehalose phosphate phosphatase domains. Trehalose seems to be a major sugar, as it shows higher levels than does glucose in hemocytes and hemolymph. Increases in HemoTPS expression, TPS enzyme activity in hemocytes, and hemolymph trehalose levels were determined 24 h after lipopolysaccharide challenge, suggesting that both TPS and TPP domains of CasHemoTPS are active and functional. The TPS gene has a wide tissue distribution in C. sapidus, suggesting multiple biosynthetic sites. A correlation between TPS activity in hemocytes and hemolymph trehalose levels was found during the molt cycle. The current study provides the first evidence of presence of trehalose in hemocytes and TPS in tissues of C. sapidus and implicates its functional role in energy metabolism and physiological adaptation.
[本文引用: 1]
DOI:10.3389/fphys.2017.00750URLPMID:5627005 [本文引用: 1]
The brown planthopper,Nilaparvata lugensis one of the most serious pests of rice, and there is so far no effective way to manage this pest. However, RNA interference not only can be used to study gene function, but also provide potential opportunities for novel pest management. The development of wing plays a key role in insect physiological activities and mainly involves chitin. Hence, the regulating role of trehalase (TRE) genes on wing bud formation has been studied by RNAi. In this paper, the activity levels of TRE and the contents of the two sugars trehalose and glucose were negatively correlated indicating the potential role of TRE in the molting process. In addition,NlTRE1-1andNlTRE2were expressed at higher levels in wing bud tissue than in other tissues, and abnormal molting and wing deformity or curling were noted 48 h after the insect was injected with any double-stranded TRE (dsTRE), even though different TREs have compensatory functions. The expression levels ofNlCHS1b, NlCht1, NlCht2, NlCht6, NlCht7, NlCht8, NlCht10, NlIDGF, andNlENGasedecreased significantly 48 h after the insect was injected with a mixture of three kinds ofdsTREs. Similarly, the TRE inhibitor validamycin can inhibitNlCHS1andNlChtgene expression. However, the wing deformity was the result of theNlIDGF, NlENGase, NlAP, andNlTSHgenes being inhibited when a singledsTREwas injected. These results demonstrate that silencing ofTREgene expression can lead to wing deformities due to the down-regulation of theAPandTSHgenes involved in wing development and that the TRE inhibitor validamycin can co-regulate chitin metabolism and the expression of wing development-related genes in wing bud tissue. The results provide a new approach for the prevention and management ofN. lugens.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}