 ,山西农业大学分子农业与生物能源研究所,山西太谷030801
,山西农业大学分子农业与生物能源研究所,山西太谷030801Phenotypic Analysis of Epoxygenase-Transgenic Soybeans
HAO QingTing, ZHANG Fei, JI XiaJie, XUE JinAi, LI RunZhi,Institute of Molecular Agriculture and Bioenergy, Shanxi Agricultural University, Taigu 030801, Shanxi通讯作者:
责任编辑: 李莉
收稿日期:2018-07-13接受日期:2018-10-11网络出版日期:2019-01-16
| 基金资助: |
Received:2018-07-13Accepted:2018-10-11Online:2019-01-16
作者简介 About authors
郝青婷,E-mail:
张飞,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (1237KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
郝青婷, 张飞, 吉夏洁, 薛金爱, 李润植. 脂肪酸环氧化酶SlEPX转基因大豆的表型分析[J]. 中国农业科学, 2019, 52(2): 191-200 doi:10.3864/j.issn.0578-1752.2019.02.001
HAO QingTing, ZHANG Fei, JI XiaJie, XUE JinAi, LI RunZhi.
0 引言
【研究意义】油料作物的种子不仅是食用油的主要来源,亦是日益重要的可再生优质原料,用于生物柴油和各种油脂化工品的商业化生产[1,2]。普通油料作物的种子油包含5种常见的脂肪酸﹕棕榈酸(16﹕0)、硬脂酸(18﹕0)、油酸(18﹕1Δ9)、亚油酸(18﹕2Δ9,12)和亚麻酸(18﹕3Δ9,12,15),这些脂肪酸已广泛应用于食品和其他工业。然而,很多未商业化的野生植物种子能合成和高水平积累一些稀有脂肪酸(unusual fatty acids,UFAs),如羟化脂肪酸、环氧化脂肪酸、乙炔酸脂肪酸和共轭脂肪酸等[3]。它们是油漆、涂料、黏合剂、增塑剂等化工产品的可再生优质原材料,具有独特的商业价值[4]。例如,蓖麻(Ricinus communis L.)种子中积累的蓖麻油酸高达总脂的90%[5],斑鸠菊(Vernonia galamensis)和琉璃菊(Stokesia laevis)种子中的环氧化脂肪酸的含量约为总脂的80%[6,7]。但是,由于这些植物存在种子小、产量低、可种植地理区域有限等不良农艺性状,使得这些稀有脂肪酸难以商业化生产以满足工业化应用的需求。因此,越来越多的研究聚焦于使用基因工程手段在普通油料作物种子中组装这些UFA的合成途径,以期实现规模化生产。大豆是中国及世界主要栽培油料作物之一,全球种植广泛,面积大,产量高[8]。大豆油的产量是世界植物油总产量的30%[9]。近年来由于生物柴油的兴起,大豆作为一种优质的生物燃油工业原料也成为研究热点之一。自2010年栽培大豆和野生型大豆基因组序列相继问世以来[10],在基因水平改造大豆的技术越来越成熟,研究周期也大幅缩短,有助于人类选育更符合要求的大豆品种。【前人研究进展】已有一些研究聚焦于在大豆种子中组装相关油脂合成途径,建立利用普通油料作物种子为平台,以期商业化生产各种高值珍稀脂肪酸。例如,棕榈油酸(16﹕1Δ9)是一种高值健康型珍稀脂肪酸,大豆种子不能合成这种脂肪酸,但含有它的前体物质棕榈酸(16﹕0)。表达源于哺乳动物的CoA-Δ9脱氢酶基因可使大豆种子棕榈酸的一半转化成棕榈油酸,含量达10%,而棕榈酸含量则从25% 降低到5%[11]。山西农业大学李润植课题组与美国肯塔基大学Hildebrand实验室先前合作研究从一种平菇(Pleurotus ostreatus)中分离到CoA-Δ9脱氢酶基因,用种子特异启动子驱动该基因在大豆体细胞胚中表达,获得棕榈油酸含量高达14%的转化体[12]。长链ω-3多不饱和脂肪酸EPA (20﹕5)和DHA(22﹕6)是人类必须从食物中获取的健康型脂肪酸,普通油料作物种子不能合成。KINNEY等[13]在大豆种子中成功组装了EPA和DHA合成途径,共表达7个相关基因(Δ6脱氢酶、Δ6延长酶、Δ15脱氢酶和Δ17脱氢酶等),转基因种子含EPA和DHA分别达20%和2.3%。环氧化脂肪酸(epoxy fatty acids,EFAs)是在酰基链上的单个或多个位置处含有跨越相邻碳原子的氧桥,使得它们具有高反应性并易于交联,用于多种高值化工品的生产[14,15,16]。工业上,植物油脂可经一系列化学反应生成各种环氧化脂肪[17,18]。相关研究表明,EFAs的生物合成是由2种类型的脂肪酸环氧化酶(P450型-环氧化酶和FAD2型-环氧化酶)催化的。脂肪酸环氧化酶以亚油酸(18﹕2Δ9,12)为底物,催化其碳链上Δ12位置处的双键加入一个氧原子,生成具有1个氧桥和一个双键的环氧化脂肪酸(12-epoxy-18﹕1Δ9),即斑鸠菊酸(Vernolic acid,Va)[19,20]。将来自于大戟属植物Euphobia lagascae的P450型-环氧化酶基因在大豆体细胞胚中表达,体细胞胚可合成8% Va[21]。将来自还阳参属植物Crepis palaestina的FAD2型-环氧化酶基因(Cpal2)在拟南芥种子中表达,Va合成积累达6.2%[22]。Cpal2表达也可使棉花种子中Va积累达8.3%[23]。【本研究切入点】前期研究已从高积累Va的琉璃菊(S. laevis)发育种子中分离到编码FAD2型-环氧化酶的SlEPX[24],并通过转基因拟南芥(Arabidopsis thaliana)验证了SlEPX催化合成Va的酶活性。进一步构建了SlEPX种子特异表达载体,采用基于体细胞胚的粒子轰击法对大豆进行遗传转化,获得种子特异表达SlEPX大豆转化体材料[25],但转基因遗传稳定性、表型及相关生理生化特征还未详尽鉴定。【拟解决的关键问题】本研究经连续选择获得表型一致、遗传稳定的高代SlEPX转基因大豆株系,重点鉴定和解析该基因种子特异表达所导致的大豆籽粒不利表型材料以及产生不利表型的分子机理。探讨SlEPX转基因大豆种子出现负效应的机制,以及进一步提高转基因种子合成积累环氧化脂肪酸和减少负效应的基因工程策略。1 材料与方法
1.1 试验材料
试验所用目的基因来自高积累环氧化脂肪酸的琉璃菊(Stokesia laevis)的环氧化酶基因SlEPX(GenBank accession,AY462108,1 137 bp),种子特异表达载体pCAMBIA1301含有种子特异性菜豆蛋白(Phaseolin)启动子驱动目的基因表达盒及潮霉素(Hyg)抗性基因选择标记。转基因受体种质材料为大豆Jack品种。经过基于体细胞胚发生的粒子轰击法进行大豆遗传转化。经连续跟踪选择鉴定获得第六代遗传稳定、表型一致的纯合转基因株系。试验于2012年至2018年在山西农业大学进行,所有转化载体和转基因大豆材料保存于山西农业大学分子农业与生物能源研究所。
1.3 叶片DNA提取及PCR
田间种植非转基因Jack品种大豆、空载体转化大豆对照及环氧化酶转基因的第六代大豆株系。在植株生长处于幼嫩时期,采用传统的CTAB法分别提取Jack及转基因大豆的顶端嫩叶的DNA,以大豆Jack品种的非转基因植株基因组为阴性对照、SlEPX表达载体DNA为阳性对照,用SlEPX特异性引物进行PCR扩增(表1),PCR产物经琼脂糖凝胶电泳检测,确定为转基因的植株挂牌标记,便于后期取样。Table 1
表1
表1引物序列
Table 1
| 引物名称 Primer name | 引物序列 Primer sequence (5′-3′) | 大小 Size (bp ) |
|---|---|---|
| SlE-PCR | F:AATCTTGCATCCTCACTGGTTTA | 23 |
| R:TGTGGTGCAGATAAGTGATTACG | 23 | |
| Actin-qPCR | F:CAGAAAGGATCTATATGGCAAC | 22 |
| R:ATTTTCTTTCTGGTGGAGCTAC | 22 | |
| SlE-qPCR | F:TCAGGCAAGAAGTACGATAGA | 21 |
| R:AACCCGATTCAGGAGACC | 18 |
新窗口打开|下载CSV
1.4 种子总RNA提取及荧光定量PCR
分别提取转基因植株叶片及3个发育时期(大豆植株开花后7、20和35 d)的种子样品,按照RNA提取试剂盒说明书提取总RNA,并反转录合成cDNA。进行实时荧光定量PCR反应,用Primer 5.0软件设计引物(表1),以Actin为内参基因,参照GenStar说明书配置荧光定量PCR体系,每个样品6个重复,反应程序为95℃ 10 min;95℃ 15 s;60℃ 1 min,40个循环。1.5 SlEPX转基因种子3个发育时期TAG含量及脂肪酸成分的测定
分别将早期(开花后12—15 d)、中期(25—35 d)、晚期(45 d)的Jack及转基因种子研磨成粉末,并称取10 mg样品置于干燥玻璃试管中。参照张玲慧[26]方法进行油脂和甲酯化的分析。甲酯化脂肪酸样品置于GC瓶中,-20℃保存用于气相检测。使用安捷伦7890B气相色谱仪检测TAG中脂肪酸的成分和含量,色谱柱型号为:HP-88,阶梯升温程序为:140℃保持5 min,然后以2℃·min-1的速率上升至210℃,最后保持12.5 min;FID的温度保持250℃。1.6 斑鸠菊酸(Va)在SlEPX转基因大豆种子不同脂类分子中的含量测定
将SlEPX转基因大豆成熟风干的种子充分研磨,称取适量样品,加入氯仿﹕甲醇(2﹕1)混匀,离心后吸出底部的氯仿相,浓缩至50—100 μL后,通过薄层层析(thin-layer chromatography,TLC)将细胞内不同类别的脂类分开,2次的展开剂分别为氯仿﹕甲醇﹕水(65﹕25﹕4)和正己烷﹕乙醚﹕乙酸(100﹕100﹕2)。将薄层板上的样品条带刮下转移到带玻璃纤维的吸管中,用氯仿﹕甲醇洗脱2次,将脂类样品用氮气吹干后加入甲醇钠进行甲酯化反应。甲酯化的样品用于GC分析。1.7 SlEPX转基因大豆种子总脂含量测定
称取50 mg野生型Jack和SlEPX转基因大豆成熟风干的种子粗粉末,3个重复,加入石英砂充分研磨,先后分2次加入7.5 mL甲醇﹕氯仿(2﹕1),置于37℃的摇床分别抽提24 h,离心后收集上层有机相合并。加入5 mL氯仿、9 mL 1%的NaCl溶液,使得氯仿﹕甲醇﹕H2O为2﹕2﹕1.8,混匀后8 000 r/min离心10 min,收集下层有机相,氮气吹干后计算脂肪酸的总重量。计算方法:总脂含量=[(提取后总重)-(提取前管重)]/0.05。1.8 SlEPX转基因大豆发芽率、种子形态、百粒重及蛋白含量的检测
百粒重测定:利用分析天平对非转基因大豆Jack和SlEPX转基因大豆进行百粒重测定,至少6次重复。发芽率测定:分别选取非转基因大豆Jack和SlEPX转基因大豆150粒种子,放入铺有专用发芽滤纸的培养皿内,50粒/皿,各设3个重复。置于20℃—25℃培养箱内,发芽率测定期间,保持滤纸湿润。以胚根突破种皮,长度达到种子长度的1/2作为发芽标准,从培养的第2天开始统计发芽数量,并将每天发芽的种子移出培养皿,直到再无种子萌发为止。计算方法:发芽率=(发芽种子总数)/(供试种子总数)×100%。
形态学观察:利用体式显微镜对非转基因大豆Jack和SlEPX转基因大豆的成熟种子进行形态学观察。
种子籽粒大小统计:随机选取转基因和空载体、非转基因对照种子样品,用一系列不同孔径的筛子将不同大小种子分开。分别统计各孔径大小的种子所占比例,至少6次重复。
蛋白含量检测:采用凯氏定氮法测定蛋白质含量,至少6次重复。
所有设置重复的数据(脂肪酸含量、总脂含量、百粒重、发芽率及蛋白质含量)均使用SPSS软件进行t检验统计分析。
2 结果
2.1 高代转基因大豆材料靶基因SlEPX整合及表达鉴定
以非转基因大豆Jack(负对照)、SlEPX转基因大豆的叶片基因组及SlEPX表达载体质粒DNA(阳性对照)为模板进行PCR扩增,结果(图1)表明,对照Jack植株基因组中未扩增出环氧化酶SlEPX片段,转基因大豆基因组和SlEPX表达载体质粒DNA中分别扩增出大小一致的目的条带(1 137 bp),证明目的基因依然稳定整合在所检测的高代转基因大豆基因组中。图1
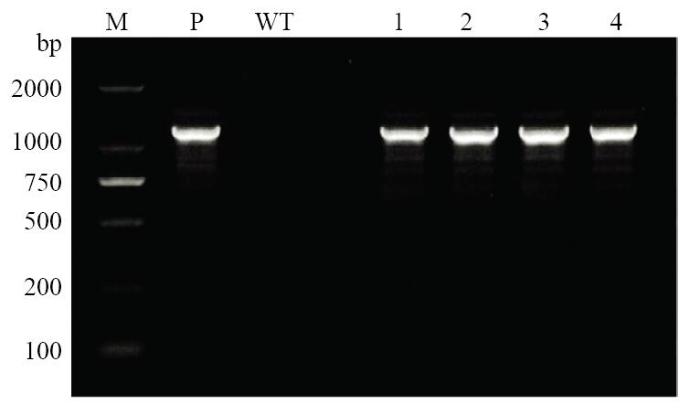 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1转基因大豆植株基因组中靶基因SlEPX的PCR检测
M:Marker;WT:非转基因Jack植株;P:SlEPX表达载体质粒DNA;1—4:SlEPX转基因大豆株系
Fig. 1Detection of the target gene SlEPX in the transgenic soybean genome by PCR analysis
M: DNA marker; WT: Wild-type soybean (negative control); P: Plasmid DNA (positive control); 1-4: SlEPX-transgenic soybean lines
通过对各样品进行实时荧光定量PCR检测(图2),目的基因SlEPX在转基因大豆种子3个不同发育时期均有表达,但在叶片中没有表达,表明目的基因在种子中特异表达。SlEPX在发育种子中的表达水平呈现标准的抛物线型增长,中期表达量最高,早期和晚期表达量低,并且中期的表达量是其他2个时期的2—3倍。这表明在高代转基因大豆中,SlEPX主要在转基因大豆种子发育中期稳定有效表达,而此时期正是大豆种子油脂高速积累期。
图2
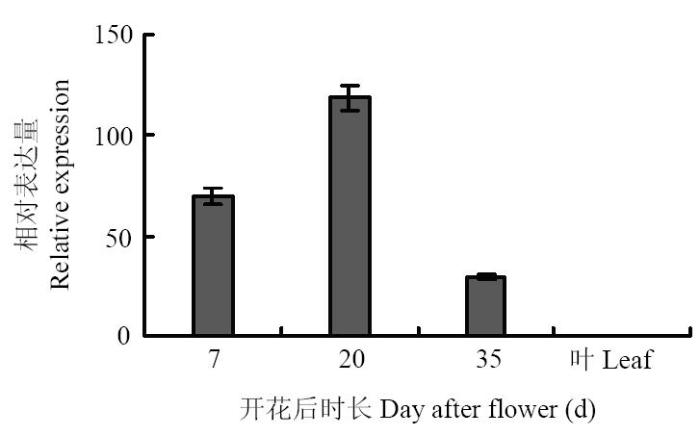 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2大豆种子不同发育时期SlEPX的表达分析
Fig. 2Expression analysis of SlEPX in the transgenic soybean seeds at different development stages
2.2 SlEPX转基因大豆种子油脂肪酸成分分析
为检测转入大豆的SlEPX编码的脂肪酸环氧化酶是否能催化合成斑鸠菊酸(Va)以及是否对其他脂肪酸合成产生影响,运用气相色谱分析转基因种子和非转基因对照种子不同发育时期样品种子油脂肪酸成分,结果(图3)显示,非转基因大豆Jack和SlEPX转基因大豆TAG中主要脂肪酸成分包括棕榈酸(16﹕0)、硬脂酸(18﹕0)、油酸(18﹕1)、亚油酸(18﹕2)和亚麻酸(18﹕3)。转基因大豆TAG中含有极少量的斑鸠菊酸(Va),仅占TAG总量的2.9%。与非转基因大豆(空载体转化体表型和非转基因大豆表型一致,下同)相比,在SlEPX转基因大豆种子油脂中其他5种主要脂肪酸含量发生了明显的变化。在种子发育中期和后期,转基因种子油中亚油酸(18﹕2)含量显著降低,亚麻酸(18﹕3)含量亦相应减少。然而,油酸(18﹕1)含量却增加。亚油酸(18﹕2)含量显著减少,说明转入的SlEPX编码的环氧化酶正确行使功能,即以亚油酸(18﹕2Δ9,12)为底物,在Δ12位置的双键处插入氧原子,生成斑鸠菊酸(Va)(12-epoxy-18﹕1Δ9)。亚油酸是亚麻酸的前体物质,亚油酸减少必然导致亚麻酸含量减低。油酸(18﹕1)含量的升高,可能是导入的FAD-型环氧化酶(SlEPX)部分干扰了大豆本身正常的FAD2催化油酸生成亚油酸的功能。图3
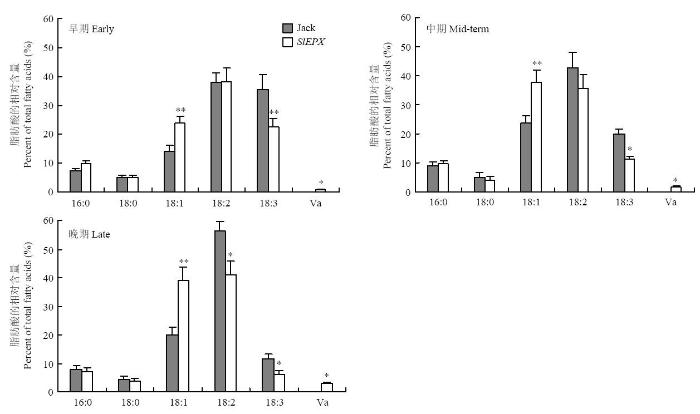 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同发育时期SlEPX转基因大豆种子油中脂肪酸成分
**和*分别表示在P<0.01和P<0.05水平差异极显著。下同
Fig. 3Fatty acid composition in seed oil of the SlEPX- transgenic soybeans at different seed development stages
** and * indicates statistically significant differences at P<0.01 and P<0.05, respectively. The same as below
2.3 SlEPX转基因大豆种子新合成的斑鸠菊酸在不同脂类分子中的分布
为解析SlEPX转基因大豆种子油中斑鸠菊酸(Va)积累量较少的机理,应用TLC对从种子萃取的总脂质样品进行层析分离,分别收集TAG和PC脂质分子样品,甲酯化后进行GC分析。结果(图4)显示,转基因大豆TAG和PC脂质分子均含有16﹕0、18﹕0、18﹕1、18﹕2、18﹕3和Va 6种脂肪酸。Va在TAG中的含量仅为2.3%,比例远远低于在PC分子中的含量(12.6%)。与此不同的是,在Va高积累的斑鸠菊和琉璃菊种子中,绝大部分Va分布在TAG分子中。图4
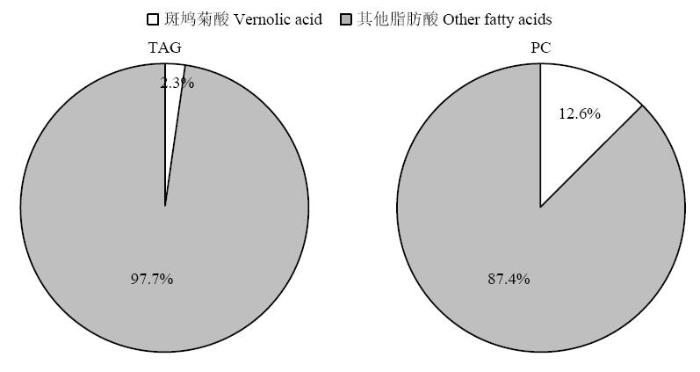 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4SlEPX转基因大豆种子中斑鸠菊酸(Va)在TAG和PC脂质分子中的分布
Fig. 4Distribution of vernolic acid (Va) in TAG and PC lipids in the SlEPX-transgenic soybean seeds
2.4 SlEPX转基因对大豆总脂含量和蛋白含量的影响
为分析种子特异表达SlEPX是否对大豆种子贮藏物积累产生影响,分别测试了非转基因大豆Jack和转基因大豆种子总脂含量和蛋白含量(图5),野生大豆Jack的总脂含量占其种子干重的21.06%,SlEPX转基因大豆的总脂含量比对照降低了5%左右,仅为其种子干重的16.84%。与非转基因大豆相比,SlEPX转基因大豆种子蛋白含量亦降低,且比对照下降了6%左右。显然,SlEPX转基因对大豆种子油脂和蛋白贮藏物积累构成负效应。图5
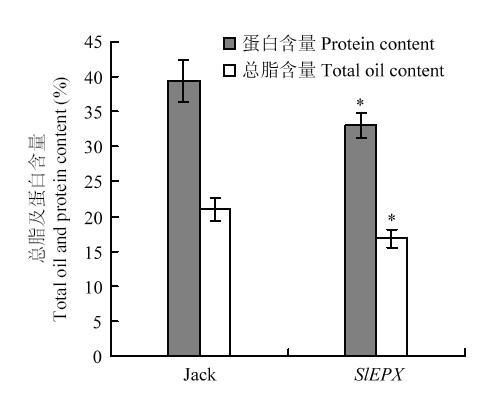 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5SlEPX转基因大豆种子总脂和总蛋白质含量
Fig. 5Total fatty acid and protein percent of the SlEPX- transgenic soybeans
2.5 SlEPX转基因对大豆百粒重、发芽率和种子其他表型的影响
为分析种子特异表达SlEPX是否对大豆种子其他农艺性状产生影响,分别测试了非转基因大豆Jack和转基因大豆种子的百粒重、发芽率、种子形态及植株表型差异(图6),转环氧化酶基因的大豆植株矮小,种子表皮皱缩,种子变长。SlEPX转基因种子百粒重比对照Jack种子减轻8.28%。对照和转基因大豆种子发芽率没有显著差异,均高达93%—96%。与对照相比,转基因大豆种子粒径<4 mm的种子所占比例明显增加。表明仅导入和种子特异表达脂肪酸环氧化酶基因,尽管能新合成一定量的高值脂肪酸Va,但也导致种子表皮皱缩、总脂和蛋白含量及百粒重减低等不良表型。这些不利农艺性状的产生可能与在大豆种子中新合成的Va未能有效从PC分子转移整合到TAG分子有关。因为Va在非原宿主细胞中产生、并主要积累在构成细胞膜系统的PC分子中,可能影响到细胞一些正常功能。图6
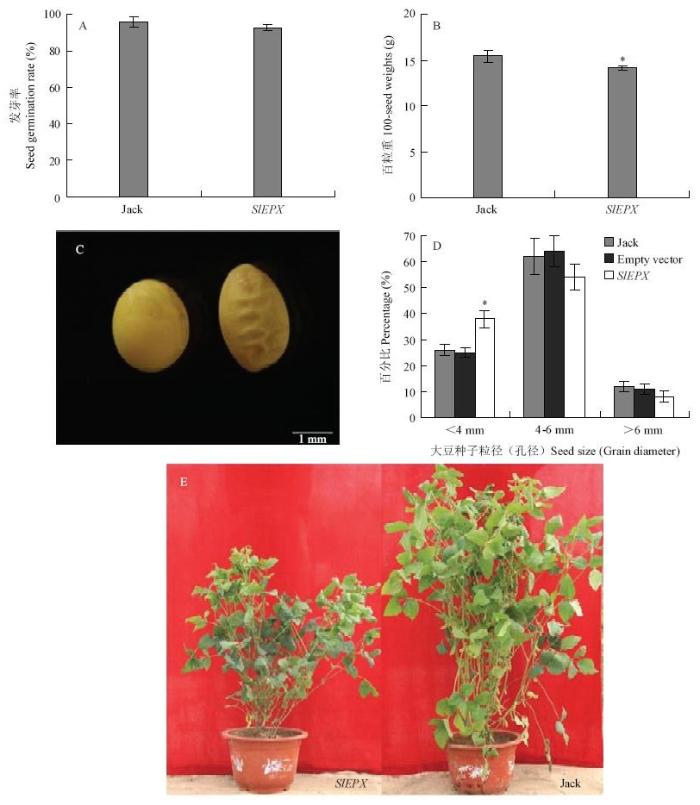 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6SlEPX转基因大豆和对照大豆籽粒性状比较
A:种子的发芽率;B:种子的百粒重;C:转基因(右)和非转基因(左)大豆种子形态;D:种子大小(粒径);E:转基因(左)和非转基因(右)大豆植株
Fig. 6Comparison of seed traits between SlEPX-transgenic and wild-type soybean
A: Seed germination rate; B: 100-seed weights; C: Seed morphology of transgenic (right) and wild-type (left) soybean; D: Seed size (grain diameter); E: Transgenic (left) and wild-type (right) soybean plants
3 讨论
3.1 异源表达催化珍稀脂肪酸合成酶的表型效应
一些野生植物种子合成积累的环氧化脂肪酸、羟化脂肪酸和共轭脂肪酸等珍稀脂肪酸(unsaturated free fatty acids,UFAs)可加工成油漆、黏合剂等多种化工产品,具有极其重要的商业应用价值[4,27],一些环氧化脂肪酸还可应用于临床医学[28,29]。现今,已从能高水平合成此类脂肪酸的非农业植物中分离到多种负责合成UFAs的关键酶基因[30]。相关研究利用这些优异基因,在传统油料作物(品种产量高、种植管理技术成熟)种子中组装特殊脂肪酸的合成途径[31]。这样所培育的工程油料作物及其推广种植,就可以实现低成本生产可再生和环境友好型工业用油原料,进而替代传统的不可再生的石油资源[32]。例如,早在1998年已从斑鸠菊和琉璃菊等植物发育种子中克隆到脂肪酸环氧化酶基因[33,34],并将这些基因转入拟南芥中,拟南芥种子合成并积累了近10%的环氧化脂肪酸[35]。在烟草愈伤中过表达源于大戟属植物Euphobia lagascae的环氧化酶基因,使烟草中积累了15%的Δ12-环氧化脂肪酸[21]。这些研究均展示了利用普通油料作物种子为平台可持续生产高值特异脂肪酸的发展前景。本文对高代SlEPX转基因大豆材料种子籽粒表型分析揭示了异源种子特异表达SlEPX,可使大豆种子新合成积累一定量的环氧化脂肪酸Va,但也导致一些不利表型。例如,与非转基因品种Jack相比,SlEPX转基因大豆油脂总量占种子干重的比例下降了5%,蛋白质含量下降了6%,百粒重也降低了8.28%,种子变得狭长且表皮皱缩。这些性状的变化均说明单独表达SlEPX对大豆的生理生化活动造成了不良的影响。类似的现象曾在转基因拟南芥研究中报道,单独表达脂肪酸环氧化酶基因导致转基因拟南芥种子不能正常发芽和生长发育受损等问题[35]。单独表达催化羟化脂肪酸(蓖麻酸)合成的脂肪酸羟化酶基因的拟南芥[36]及表达α-桐酸合成基因的大豆体细胞胚亦出现负效应[37,38]。这些报道和本文研究结果均表明异源特殊脂肪酸的合成积累会影响宿主种子的代谢和生理。一些研究显示,蓖麻酸在异源内质网上合成而不能有效转运储藏于油体,就会诱导质体中乙酰-CoA羧化酶活性的翻译后抑制,从而抑制脂肪酸合成和总油脂积累[39]。本文研究发现SlEPX转基因大豆新合成的Va大部分结合在PC分子(12.6%)上,而在TAG分子中极少。新合成珍稀脂肪酸Va大量分布在膜脂PC分子中,势必影响到宿主细胞的生命活动,进而导致大豆种子产生一些不利表型。
3.2 在普通油料作物种子中组装珍稀油脂合成途径的基因工程优化策略
对斑鸠菊和琉璃菊发育种子中斑鸠菊酸合成积累研究发现,结合在TAG分子而储藏于油体的斑鸠菊酸含量可高达50%—80%[6,7],相反,结合于细胞膜PC分子中的斑鸠菊酸极少。这表明,这些原产斑鸠菊酸的植物经长期进化,细胞形成了独特机制,能高效地将脂肪酸环氧化酶催化合成的结合于PC的斑鸠菊酸转移整合到TAG分子而储存于油体中。DGAT(二酰基甘油酰基转移酶)和PDAT(磷脂二酰甘油酰基转移酶)是控制TAG合成的最后一步酰化反应的限速酶,它们能将CoA分子结合的酰基或PC分子上sn-2位置的酰基链转移到DAG(二酰甘油)分子的sn-2位置而合成TAG分子。立体化学分析证实,斑鸠菊酸等特异脂肪酸合成后就结合在PC的sn-2位置。显然,原产这些珍稀脂肪酸的植物细胞存在能特异识别这些珍稀脂肪酸酰基、并将其从PC分子转移整合到TAG分子中的酰基转移酶(DGAT和PDAT等)。这在异源植物种子中组装原产于蓖麻的蓖麻酸合成途径相关研究得到证实。例如,在拟南芥种子中特异性表达蓖麻酸羟化酶(RcFAH)时,种子新合成羟化脂肪酸(蓖麻酸),但是含量低,仅占种子油的17%,且大多数分布在细胞膜PC分子上[40,41]。与野生型拟南芥种子相比,转RcFAH的拟南芥种子油脂含量下降。将来自蓖麻发育种子RcPDAT1A或RcDGAT2与RcFAH共表达,转基因拟南芥的油脂含量接近在蓖麻种子的水平,新合成的羟化脂肪酸含量分别上升至27%和30%[42,43]。山西农业大学李润植课题组与美国肯塔基大学Hildebrand实验室先前合作研究发现高积累环氧化脂肪酸Va的斑鸠菊、琉璃菊发育种子中的DGAT对Va有底物特异性,能有效将Va-酰基转移到TAG分子中[44]。然而,大豆本身的DGAT对环氧化脂肪酸酰基没有底物选择性[45]。本研究检测的SlEPX转基因大豆种子虽然合成了Va,但在TAG中仅有2.3%左右,相反在PC中的Va的高达12.6%。推测转SlEPX大豆种子中Va含量低且在TAG分子积累少的原因是大豆本身缺少像斑鸠菊或琉璃菊能特异高效将环氧化脂肪酸酰基从CoA或PC转入TAG的DGAT或PDAT等相关酶。据此,我们提出在大豆种子中组装环氧化脂肪酸合成途径的优化策略,即在种子特异表达环氧化酶SlEPX大豆种子中,进一步共表达来自斑鸠菊的DGAT或PDAT以及其他相关基因,能够将结合于CoA及PC分子中的Va酰基有效转移到TAG分子中,大幅提高Va在种子油中的积累量。同时,消除新合成积累的Va对细胞伤害,进而培育获得富含环氧化脂肪酸且其他农艺性状优良的大豆新种质,为这些高值珍稀脂肪酸的规模化绿色生产和应用奠定基础。4 结论
种子特异表达异源脂肪酸环氧化酶(SlEPX)可使大豆种子合成环氧化脂肪酸即斑鸠菊酸(Va),但含量低且有负效应。在SlEPX转基因大豆种子中共表达DGAT或PDAT,可实现大幅提高Va在种子油中的积累量,并消除Va积累于细胞膜所导致的不利表型。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/j.pbi.2010.01.008URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/S1369-5266(03)00013-XURLPMID:12667876 [本文引用: 2]

Unusual fatty acids that have useful industrial properties occur widely in the seed oils of many non-agronomic plant species. Researchers are attempting to use biotechnology to produce high levels of these fatty acids in the seeds of existing crop plants. cDNAs for a wide variety of unusual fatty acid biosynthetic enzymes have been identified, particularly through the use of expressed sequence tags. However, it has not yet been possible to use these cDNAs to produce large amounts of unusual fatty acids in seeds of transgenic plants. This difficulty points to the need for a greater understanding of fatty acid metabolism in oilseeds.
URL [本文引用: 1]
As one of the world's ten largest oil crops,castor has broad utilization prospects based on its high economic value.This paper analyzes the current situation of high-yielding castor cultivation technology,summarizes the comprehensive utilization of castor and describes the development of castor biodiesel fuel.Castor salt resistance and the remediation of saline-alkali soil by castor cultivationis are also discussed,as well as the potential of China's cultivation of castor.In this paper, recommendation for castor research and industrial development were also proposed, including the castor surrey and zoning in suitable planting area, the investigation and innovation of germplasm resources, high-yielding cultivation technology of castor in low quality and non-farming soil, preparation process of castor biodiesel fuel, comprehensive utilization research of castor, sustainable development and security system of castor industry.
URL [本文引用: 1]
As one of the world's ten largest oil crops,castor has broad utilization prospects based on its high economic value.This paper analyzes the current situation of high-yielding castor cultivation technology,summarizes the comprehensive utilization of castor and describes the development of castor biodiesel fuel.Castor salt resistance and the remediation of saline-alkali soil by castor cultivationis are also discussed,as well as the potential of China's cultivation of castor.In this paper, recommendation for castor research and industrial development were also proposed, including the castor surrey and zoning in suitable planting area, the investigation and innovation of germplasm resources, high-yielding cultivation technology of castor in low quality and non-farming soil, preparation process of castor biodiesel fuel, comprehensive utilization research of castor, sustainable development and security system of castor industry.
[本文引用: 2]
DOI:10.1016/0926-6690(94)90065-5URL [本文引用: 2]
Considerable variation in plant and flower characteristics was observed among seventeen accessions of the Vernonia galamensis (Cass.) Less. complex, including representatives of all but one subspecies. With the exception of ssp. nairobensis , a high percentage of plants of all accessions had flowered by 121 days after planting (DAP) in Arizona on the 9th of January. Mean days to flower for various accessions varied from 37 to 121 days, and those flowering by 121 DAP had mean numbers of capitula ranging from 0.1 to 86 per plant. Fully mature capitula on accessions ranged from 0 to 55%, and plant height varied from 23 to 88 cm at 121 DAP. Seeds of five accessions from three subspecies harvested at full maturity had significantly higher oil and vernolic acid contents, and seed weights than comparable samples harvested at a less mature stage when involucres surrounding maturing seeds were still green in color. Seeds of uniform lots of the day-neutral accession of V. galamensis ssp. galamensis var. petitiana [A 20295 (V 029)], grown at six locations throughout the United States, had highly significant differences in oil percentage (34.5–44.3%), vernolic acid content (61.0–80.0%), and seed weight (1.87–2.91 g/1000). Although environmental factors play a significant role in seed oil and vernolic acid contents, and seed weight, there appears to be no serious constraints to the ultimate domestication and commercialization of vernonia for production in temperate areas. However, considerable genetic improvement is required, and further evaluation, selection, and development of germplasm within various climatic and geographical locations will be necessary to maximize adaptation and yield.
[本文引用: 1]
[本文引用: 1]
DOI:10.3969/j.issn.1000-9841.2013.01.029URL [本文引用: 1]
植物种子是油脂的主要来源之一,种子中的油脂主要在其发育过程中积累。油脂的积累除受脂肪酸合成途径中多种酶的活性、特异性和稳定性的影响,更受到转录前、转录和转录后水平的调控。众多相关基因参与此过程,形成一系列基因网络调控了下游油脂合成基因的表达。文章综述了转录因子对油脂积累的调控、DNA甲基化和组蛋白修饰相关基因在种子发育和油脂积累中的作用以及小RNA调控种子发育的分子机理,并介绍了大豆油脂积累调控机制的研究进展。
DOI:10.3969/j.issn.1000-9841.2013.01.029URL [本文引用: 1]
植物种子是油脂的主要来源之一,种子中的油脂主要在其发育过程中积累。油脂的积累除受脂肪酸合成途径中多种酶的活性、特异性和稳定性的影响,更受到转录前、转录和转录后水平的调控。众多相关基因参与此过程,形成一系列基因网络调控了下游油脂合成基因的表达。文章综述了转录因子对油脂积累的调控、DNA甲基化和组蛋白修饰相关基因在种子发育和油脂积累中的作用以及小RNA调控种子发育的分子机理,并介绍了大豆油脂积累调控机制的研究进展。
DOI:10.1038/nature08670URLPMID:20075913 [本文引用: 1]
Soybean (Glycine max) is one of the most important crop plants for seed protein and oil content, and for its capacity to fix atmospheric nitrogen through symbioses with soil-borne microorganisms. We sequenced the 1.1-gigabase genome by a whole-genome shotgun approach and integrated it with physical and high-density genetic maps to create a chromosome-scale draft sequence assembly. We predict 46,430 protein-coding genes, 70% more than Arabidopsis and similar to the poplar genome which, like soybean, is an ancient polyploid (palaeopolyploid). About 78% of the predicted genes occur in chromosome ends, which comprise less than one-half of the genome but account for nearly all of the genetic recombination. Genome duplications occurred at approximately 59 and 13 million years ago, resulting in a highly duplicated genome with nearly 75% of the genes present in multiple copies. The two duplication events were followed by gene diversification and loss, and numerous chromosome rearrangements. An accurate soybean genome sequence will facilitate the identification of the genetic basis of many soybean traits, and accelerate the creation of improved soybean varieties. 2010 Macmillan Publishers Limited. All rights reserved.
DOI:10.1007/BF02318963URL [本文引用: 1]
Somatic embryo cycling, a modification of soyabean somatic embryogenic suspension culture, was developed as an efficient and rapid method of producing tissue suitable for stable transformation of soyabean germplasm by biolistic particle bombardment.Instead of using immature seed explants, cotyledon-stage somatic embryo hypocotyls were placed on auxin-containing medium, where they initiated new somatic embryos primarily from single epidermal cells. By bombarding hypocotyls prior to initiation of subsequent embryo formation, transformed soyabean somatic embryos were effectively transformed with the reporter genes neomycin phosphotransferase, beta-glucuronidase and a mammalian stearyl CoA delta-9 desaturase, controlled by a seed-specific promoter. These embryos contained significantly reduced levels of saturated palmitic and stearic fatty acids, and significant amounts of monounsaturated palmitoleic acid, which is not normally abundant in soyabean seeds. This study demonstrates the effectiveness ofsomatic embryo cycling for soyabean transformation and for testing expression of genes for seed-specific proteins. Abnormal flower development in recovered plants is a limitation for application of the technique to produce transgenic seed at present.
URL [本文引用: 1]
植物种子油可提供人类营养所需的多种脂肪酸,也是工业用油的原料之一。文章结合我们对植物种子发育、脂肪酸生物合成途径和大豆油脂遗传改良的研究,重点论述参与脂肪酸合成及其调控的一些关键酶的基因、代谢工程改良植物油脂营养价值的技术策略及其研究进展,分析目前应用油料作物种子作为“生物反应器”规模化生产有重要营养价值和特殊用途的脂肪酸的问题及技术“瓶颈”,讨论未来植物脂肪酸代谢工程主攻方向以及在培育可再生资源和推动人类社会及经济可持续发展中的应用前景。
URL [本文引用: 1]
植物种子油可提供人类营养所需的多种脂肪酸,也是工业用油的原料之一。文章结合我们对植物种子发育、脂肪酸生物合成途径和大豆油脂遗传改良的研究,重点论述参与脂肪酸合成及其调控的一些关键酶的基因、代谢工程改良植物油脂营养价值的技术策略及其研究进展,分析目前应用油料作物种子作为“生物反应器”规模化生产有重要营养价值和特殊用途的脂肪酸的问题及技术“瓶颈”,讨论未来植物脂肪酸代谢工程主攻方向以及在培育可再生资源和推动人类社会及经济可持续发展中的应用前景。
[本文引用: 1]
DOI:10.1104/pp.104.3.827URLPMID:12232129 [本文引用: 1]
Abstract We have previously demonstrated that the double bond of petroselinic acid (18:1[delta]6cis) in coriander (Coriandrum sativum L.) seed results from the activity of a 36-kD desaturase that is structurally related to the [delta]9-stearoyl-acyl carrier protein (ACP) desaturase (E.B. Cahoon, J. Shanklin, J.B. Ohlrogge [1992] Proc Natl Acad Sci USA 89: 11184-11188). To further characterize the biosynthetic pathway of this unusual fatty acid, 14C-labeling experiments were conducted using developing endosperm of coriander. Studies were also performed using suspension cultures of transgenic tobacco (Nicotiana tabacum L.) that express the coriander 36-kD desaturase, and as a result produce petroselinic acid and [delta]4-hexadecenoic acid. When supplied exogenously to coriander endosperm slices, [1-14C]palmitic acid and stearic acid were incorporated into glycerolipids but were not converted to petroselinic acid. This suggested that petroselinic acid is not formed by the desaturation of a fatty acid bound to a glycerolipid or by reactions involving acyl-coenzyme As (CoA). Instead, evidence was most consistent with an acyl-ACP route of petroselinic acid synthesis. For example, the exogenous feeding of [1-14C]lauric acid and myristic acid to coriander endosperm slices resulted in the incorporation of the radiolabels into long-chain fatty acids, including primarily petroselinic acid, presumably through acyl-ACP-associated reactions. In addition, using an in vitro fatty acid biosynthetic system, homogenates of coriander endosperm incorporated [2-14C]malonyl-CoA into petroselinic acid, of which a portion was detected in a putative acyl-ACP fraction. Furthermore, analysis of transgenic tobacco suspension cultures expressing the coriander 36-kD desaturase revealed significant amounts of petroselinic acid and [delta]4-hexadecenoic acid in the acyl-ACP pool of these cells. Also presented is evidence derived from [U-14C]nonanoic acid labeling of coriander endosperm, which demonstrates that the coriander 36-kD desaturase positions double bonds relative to the carboxyl end of acyl-ACP substrates. The data obtained in these studies are rationalized in terms of a biosynthetic pathway of petroselinic acid involving the [delta]4 desaturation of palmitoyl-ACP by the 36-kD desaturase followed by two-carbon elongation of the resulting [delta]4-hexadecenoyl-ACP.
DOI:10.1016/j.biochi.2009.03.021URLPMID:19375482 [本文引用: 1]
Our society is highly depending on petroleum for its activities. About 90% is used as an energy source for transportation and for generation of heat and electricity and the remaining as feedstocks in the chemical industry. However, petroleum is a finite source as well as causing several environmental problems such as rising carbon dioxide levels in the atmosphere. Petroleum therefore needs to be replaced by alternative and sustainable sources. Plant oils and oleochemicals derived from them represent such alternative sources, which can deliver a substantial part of what is needed to replace the petroleum used as feedstocks. Plant derived feedstock oils can be provided by two types of oil qualities, multi-purpose and technical oils. Multi-purpose oils represent oil qualities that contain common fatty acids and that can be used for both food and feedstock applications. Technical oil qualities contain unusual fatty acids with special properties gained from their unique molecular structure and these types of oils should only be used for feedstock applications. As a risk mitigation strategy in the selection of crops, technical oil qualities should therefore preferably be produced by oil crop platforms dedicated for industrial usage. This review presents a short survey of oil crop platforms to be considered for either multi-purpose or technical oils production. Included among the former platforms are some of the major oil crops in cultivation such as oil palm, soybean and rapeseed. Among the later are those that could be developed into dedicated industrial platforms such as crambe, flax, cotton and Brassica carinata. The survey finishes off by highlighting the potential of substantial increase in plant oil production by developing metabolic flux platforms, which are starch crops converted into oil crops.
DOI:10.1074/jbc.M304862200URLPMID:12855676 [本文引用: 1]
Abstract RF2a is a bZIP transcription factor that regulates expression of the promoter of rice tungro bacilliform badnavirus. RF2a is predicted to include three domains that contribute to its function. The results of transient assays with mutants of RF2a from which one or more domains were removed demonstrated that the acidic domain was essential for the activation of gene expression, although the proline-rich and glutamine-rich domains each played a role in this function. Studies using fusion proteins of different functional domains of RF2a with the 2C7 synthetic zinc finger DNA-binding domain showed that the acidic region is a relatively strong activation domain, the function of which is dependent on the context in which the domain is placed. Data from transgenic plants further supported the conclusion that the acidic domain was important for maintaining the biological function of RF2a. RF2a and TBP (TATA-binding protein) synergistically activate transcription in vitro (Zhu, Q., Ordiz, M. I., Dabi, T., Beachy, R. N., and Lamb, C. (2002) Plant Cell 14, 795-803). In vitro and in vivo assays showed that RF2a interacts with TBP through the glutamine-rich domain but not the acidic domain. Functional analysis of such interactions indicates that the acidic domain activates transcription through mechanisms other than via the direct recruitment of TBP.
DOI:10.1016/j.chroma.2017.12.066URLPMID:29370919 [本文引用: 1]
Abstract Epoxy and hydroxy fatty acids are important intermediates during lipid oxidation; quantification of both structures may help evaluate the extent of competition among various lipid oxidation pathways. This article describes a method to simultaneously determine saturated- and unsaturated- epoxy and hydroxy fatty acids derived from oxidation of vegetable oils. The experimental procedures employed transesterification with sodium methoxide, separation of epoxy and hydroxy fatty acid methyl esters (FAME) using solid-phase extraction (SPE), and trimethylsilyl (TMS) derivatization of hydroxy groups. GC-MS was used to identify the epoxy and hydroxy FAME in two different SPE fractions, while GC-flame ionization detection (GC-FID) was used to determine their quantities. Epoxy-octadecanoate/octadecenoate and hydroxy-octadecanoate/octadecenoate/octadecadienoate were determined as lipid oxidation products generated from oxidation of sunflower and canola oils. An isomer of methyl 13-hydroxyoctadeca-9,11-dienoate (13-HODE) TMS ether co-eluted with methyl 15-hydroxyoctadeca-9,12-dienoate TMS ether, which was only present in canola oil; thus, GC-MS-selected ion monitoring (GC-MS-SIM) was used to determine the concentration of 13-HODE. The proposed method has been successfully applied to monitor epoxy and hydroxy fatty acids in sunflower oil and canola oil oxidized at 40鈥癈.
[本文引用: 1]
DOI:10.1006/abbi.1993.1265URLPMID:8489257 [本文引用: 1]
Epoxidated fatty acids are the major constituents of the triacylglycerols in a few plant species. We have investigated the biosynthesis of vernolic acid (cis-12-epoxyoctadeca-cis-9-enoic acid) in the seed oil of Euphorbia lagascae. Microsomes were isolated from developing endosperm. The membrane lipids were labeled in situ with [14C]oleate or [14C]linoleate, which mainly were recovered in phosphatidylcholine (PC), and the metabolization of the radioactive fatty acids was followed in incubations with or without NADPH. In the presence of NADPH, [14C]vernoleate was formed. After short incubations, most of the vernolic acid was found in PC, but with increasing incubation times, the free acid dominated. The synthesis of vernoleate was inhibited by carbon monoxide, but not by cyanide. The presence of anticytochrome b5 antibodies inhibited both the desaturation of [14C]oleate to [14C]linoleate and the epoxidation of [14C]linoleate to [14C]vernoleate. Free linoleic acid did not serve as substrate for epoxidation. The results indicate that, in the endosperm of E. lagascae, vernoleate is synthesized on PC from linoleate, and that the epoxidation is catalyzed by a cytochrome P450 and involves cytochrome b5.
[本文引用: 1]
DOI:10.2307/4280328URL [本文引用: 2]
http://www.jstor.org/stable/4280328
DOI:10.1126/science.280.5365.915URLPMID:9572738 [本文引用: 1]
Acetylenic bonds are present in more than 600 naturally occurring compounds. Plant enzymes that catalyze the formation of the 螖12 acetylenic bond in 9-octadecen-12-ynoic acid and the 12 epoxy group in 12,13-epoxy-9-octadecenoic acid were characterized, and two genes, similar in sequence, were cloned. When these complementary DNAs were expressed in Arabidopsis thaliana, the content of acetylenic or epoxidated fatty acids in the seeds increased from 0 to 25 or 15 percent, respectively. Both enzymes have characteristics similar to the membrane proteins containing non-heme iron that have histidine-rich motifs.
DOI:10.1071/FP05297URL [本文引用: 1]
The transgenic production of unusual fatty acids in oil seed crops offers an alternative, renewable resource for industry. However, transgenic expression of genes catalysing the synthesis of unusual fatty acids has generally resulted in these fatty acids accumulating at levels significantly below the levels in the wild species from which the genes were sourced. This study reports expression of additional copies of any of three 12-desaturase genes (FAD2) from Crepis palaestina Bornm., cotton (Gossypium hirsutum L.) or Arabidopsis thaliana (L.) Heynh. with C. palaestina 12-epoxygenase gene (Cpal2), in an Arabidopsis mutant having a significantly higher level of linoleic acid substrate. This resulted in the highest levels of vernolic acid accumulation, 21% of total fatty acids, reported so far in any transgenic plant expressing the 12-epoxygenase. Similarly, the co-expression of C. palaestina Cpal2 and a transgenic copy of FAD2 in cotton seed that contains large amounts of linoleic acid substrate also resulted in greater accumulation of vernolic acid in seed than did expression of C. palaestina Cpal2 alone.
[本文引用: 1]
DOI:10.1016/j.ymben.2011.11.004URLPMID:22107928 [本文引用: 1]
78 Expression of a Stokesia laevis epoxygenase (SlEPX) in soybeans achieves 3–7% epoxide levels. 78 SlEPX-transgenic seeds showed a 2–5% reduction in oil and protein levels of 30.9–51.4%. 78 These side effects were overcome with Vernonia galamensis DGAT1 and 2 coexpression. 78 DGAT1 and 2 restored oil and protein contents to normal levels. 78 DGAT1 and 2, especially 2 greatly increase epoxide accumulation in the oil.
[D].
[本文引用: 1]
[D].
[本文引用: 1]
DOI:10.1016/j.pbi.2007.04.005URLPMID:17434788 [本文引用: 1]
Oilseeds provide a unique platform for the production of high-value fatty acids that can replace non-sustainable petroleum and oceanic sources of specialty chemicals and aquaculture feed. However, recent efforts to engineer the seeds of crop and model plant species to produce new types of fatty acids, including hydroxy and conjugated fatty acids for industrial uses and long-chain omega-3 polyunsaturated fatty acids for farmed fish feed, have met with only modest success. The collective results from these studies point to metabolic ‘bottlenecks’ in the engineered plant seeds that substantially limit the efficient or selective flux of unusual fatty acids between different substrate pools and ultimately into storage triacylglycerol. Evidence is emerging that diacylglycerol acyltransferase 2, which catalyzes the final step in triacylglycerol assembly, is an important contributor to the synthesis of unusual fatty acid-containing oils, and is likely to be a key target for future oilseed metabolic engineering efforts.
DOI:10.5650/jos.ess17222URL [本文引用: 1]
DOI:10.1021/acs.jmedchem.8b00243URL [本文引用: 1]
Accumulating studies have linked inflammation to tumor progression. Dietary omega-3 fatty acids including docosahexaenoic acid (DHA) have been shown to suppress tumor growth through their conversion to epoxide metabolites. Alternatively, DHA is converted enzymatically into docosahexaenoylethanolamide (DHEA), an endocannabinoid with anti-proliferative activity. Recently, we reported a novel class of anti-inflammatory DHEA-epoxides (EDP-EAs) that contain both ethanolamide and epoxide moieties. Herein we evaluate the anti-tumorigenic properties of EDP-EAs in an osteosarcoma model. First, we show ~80% increase in EDP-EAs in metastatic lungs versus normal mouse lungs. We found significant differences in the apoptotic and anti-migratory potency of the different EDP-EA regioisomers, which are partly mediated through cannabinoid receptor 1 (CB1). Furthermore, we synthesized derivatives of the most pro-apoptotic regioisomer. These derivatives had reduced hydrolytic susceptibility to fatty acid-amide hydrolase and ...
DOI:10.1016/j.bbalip.2014.07.020URLPMID:25093613 [本文引用: 1]
61PUFA are oxidized to PUFA epoxides by cytochrome P450 epoxygenases.61PUFA epoxides function as mediators in the cardiovascular, renal and nervous system.61PUFA epoxides activate receptor-mediated signaling pathways and ion channels.61Soluble epoxide hydrolase inactivates PUFA epoxides by hydrolysis to PUFA diols.61Inhibition of soluble epoxide hydrolase potentiates the action of PUFA epoxides.
DOI:10.1111/tpj.13172URL [本文引用: 1]
Plant seed lipid metabolism is an area of intensive research, including many examples of transgenic events in which oil composition has been modified. In the selected examples described in this review, progress towards the predictive manipulation of metabolism and the reconstitution of desired traits in a non‐native host is considered. The advantages of a particular oilseed crop,Camelina sativa, as a flexible and utilitarian chassis for advanced metabolic engineering and applied synthetic biology are considered, as are the issues that still represent gaps in our ability to predictably alter plant lipid biosynthesis. Opportunities to deliver useful bio‐based products via transgenic plants are described, some of which represent the most complex genetic engineering in plants to date. Future prospects are considered, with a focus on the desire to transition to more (computationally) directed manipulations of metabolism. In this review we advance the oilseed plantCamelina sativaas a useful platform for metabolic engineering of lipids, and propose that a new paradigm of predictive manipulation will exceed progress made thus far with iterative approaches.
DOI:10.1002/biot.201400197URLPMID:27440175 [本文引用: 1]
Abstract Fatty acid methyl ester analysis (FAME) by gas chromatography coupled to mass spectrometry (GC-MS) is a widely used technique in biodiesel/bioproduct (e.g. poly-unsaturated fatty acids, PUFA) research but typically does not allow distinguishing between bound and free fatty acids. To understand and optimize biosynthetic pathways, however, the origin of the fatty acid is an important information. Furthermore the annotation of PUFAs is compromised in classical GC-EI-MS because the precursor molecular ion is missing. In the present protocol an alkaline methyl esterification step with TMS derivatization enabling the simultaneous analysis of bound and free fatty acids but also further lipids such as sterols in one GC-MS chromatogram is combined. This protocol is applied to different lipid extracts from single cell algae to higher plants: Chlorella vulgaris, Chlamydomonas reinhardtii, Coffea arabica, Pisum sativum and Cuscuta japonica . Further, field ionization (GC-FI-MS) is introduced for a better annotation of fatty acids and exact determination of the number of double bonds in PUFAs. The proposed workflow provides a convenient strategy to analyze algae and other plant crop systems with respect to their capacity for third generation biodiesel and high-quality bioproducts for nutrition such as PUFAs.
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s004250000456URLPMID:11346964 [本文引用: 2]
The Crepis palaestina cDNA Cpal2 encodes a Δ12-epoxygenase that can catalyse the synthesis of 12,13-epoxy-cis-9-octadecenoic acid (18:1E) from linoleic acid (18:2). When the Cpal2 gene was expressed under the control of the napin seed-specific promoter in Arabidopsis thaliana (L.) Heynh., the seed lipids accumulated only low levels of 18:1E and also 12,13-epoxy-cis-9,15-octadec-2-enoic acid (18:2E). Despite the fact that the levels of these epoxy fatty acids comprised only up to 6.2% of the total fatty acids, there was a very marked increase in oleic acid (18:1) and decrease in linoleic (18:2) and α-linolenic (18:3) acids in these plants, indicating that endogenous Δ12-desaturation was greatly reduced in these plants. Significant between-line differences in the levels of Cpal2 mRNA were observed during seed development, but were not associated with any major variation in mRNA levels for the endogenous Arabidopsis Δ12-desaturase (Fad2). This suggests that if an unfavourable interaction occurs between the transgenic Δ12-epoxygenase and the endogenous Δ12-desaturase, which decreases the level of desaturation, it occurs at either the translational or post-translational level. We further show that the co-expression of a Δ12-desaturase gene from C. palaestina in Cpal2 transgenic Arabidopsis returns the relative proportions of the C18 seed fatty acids to normal levels and results in an almost twofold increase in total epoxy fatty acids.
[本文引用: 1]
DOI:10.1016/j.phytochem.2006.04.013URLPMID:16762380 [本文引用: 1]
Expression of 12-oleic acid desaturase-related fatty acid conjugases from Calendula officinalis, Momordica charantia, and Vernicia fordii in seeds of soybean ( Glycine max) or an Arabidopsis thaliana fad3/fae1 mutant was accompanied by the accumulation of the conjugated fatty acids calendic acid or 伪-eleostearic acid to amounts as high as 20% of the total fatty acids. Conjugated fatty acids, which are synthesized from phosphatidylcholine (PC)-linked substrates, accumulated in PC and phosphatidylethanolamine, and relative amounts of these fatty acids were higher in PC than in triacylglycerol (TAG) in the transgenic seeds. The highest relative amounts of conjugated fatty acids were detected in PC from seeds of soybean and A. thaliana that expressed the C. officinalis and M. charantia conjugases, where they accounted for nearly 25% of the fatty acids of this lipid class. In these seeds, >85% of the conjugated fatty acids in PC were detected in the sn-2 position, and these fatty acids were also enriched in the sn-2 position of TAG. In marked contrast to the transgenic seeds, conjugated fatty acids composed <1.5% of the fatty acids in PC from seeds of five unrelated species that naturally synthesize a variety of conjugated fatty acid isomers, including seeds that accumulate conjugated fatty acids to >80% of the total fatty acids. These results suggest that soybean and A. thaliana seeds are deficient in their metabolic capacity to selectively catalyze the flux of conjugated fatty acids from their site of synthesis on PC to storage in TAG.
DOI:10.1074/jbc.M009188200URL [本文引用: 1]
DOI:10.1073/pnas.1318511111URLPMID:24398521 [本文引用: 1]
Degradation of unusual fatty acids through -oxidation within transgenic plants has long been hypothesized as a major factor limiting the production of industrially useful unusual fatty acids in seed oils. Arabidopsis seeds expressing the castor fatty acid hydroxylase accumulate hydroxylated fatty acids up to 17% of total fatty acids in seed triacylglycerols; however, total seed oil is also reduced up to 50%. Investigations into the cause of the reduced oil phenotype through in vivo [(14)C]acetate and [(3)H]2O metabolic labeling of developing seeds surprisingly revealed that the rate of de novo fatty acid synthesis within the transgenic seeds was approximately half that of control seeds. RNAseq analysis indicated no changes in expression of fatty acid synthesis genes in hydroxylase-expressing plants. However, differential [(14)C]acetate and [(14)C]malonate metabolic labeling of hydroxylase-expressing seeds indicated the in vivo acetyl-CoA carboxylase activity was reduced to approximately half that of control seeds. Therefore, the reduction of oil content in the transgenic seeds is consistent with reduced de novo fatty acid synthesis in the plastid rather than fatty acid degradation. Intriguingly, the coexpression of triacylglycerol synthesis isozymes from castor along with the fatty acid hydroxylase alleviated the reduced acetyl-CoA carboxylase activity, restored the rate of fatty acid synthesis, and the accumulation of seed oil was substantially recovered. Together these results suggest a previously unidentified mechanism that detects inefficient utilization of unusual fatty acids within the endoplasmic reticulum and activates an endogenous pathway for posttranslational reduction of fatty acid synthesis within the plastid.
[本文引用: 1]
DOI:10.1111/j.1365-313X.2005.02636.xURLPMID:16460516 [本文引用: 1]
It is desirable to produce high homogeneity of novel fatty acids in oilseeds through genetic engineering to meet the increasing demands of the oleo-chemical industry. However, expression of key enzymes for biosynthesis of industrial fatty acids usually results in low levels of desired fatty acids in transgenic oilseeds. The abundance of derivatized fatty acids in their natural species suggests that additional genes are needed for high production in transgenic plants. We used the model oilseed plant Arabidopsis thaliana expressing a castor fatty acid hydroxylase (FAH12) to identify genes that can boost hydroxy fatty acid accumulation in transgenic seeds. Here we describe a high-throughput approach that, in principle, can allow testing of the entire transcriptome of developing castor seed endosperm by shotgun transforming a full-length cDNA library into an FAH12-expressing Arabidopsis line. The resulting transgenic seeds were screened by high-throughput gas chromatography. We obtained several lines transformed with castor cDNAs that contained increased amounts of hydroxy fatty acids in transgenic Arabidopsis. These cDNAs were then isolated by PCR and retransformed into the FAH12-expressing line, thus confirming their beneficial contributions to hydroxy fatty acid accumulation in transgenic Arabidopsis seeds. Although we describe an approach that is targeted to oilseed engineering, the methods we developed can be applied in many areas of plant biotechnology and functional genomic research.
DOI:10.1104/pp.110.167239URLPMID:21173026 [本文引用: 1]
Producing unusual fatty acids (FAs) in crop plants has been a long-standing goal of green chemistry. However, expression of the enzymes that catalyze the primary synthesis of these unusual FAs in transgenic plants typically results in low levels of the desired FA. For example, seed-specific expression of castor (Ricinus communis) fatty acid hydroxylase (RcFAH) in Arabidopsis (Arabidopsis thaliana) resulted in only 17% hydroxy fatty acids (HFAs) in the seed oil. In order to increase HFA levels, we investigated castor phospholipidrdiacylglycerol acyltransferase (PDAT). We cloned cDNAs encoding three putative PDAT enzymes from a castor seed cDNA library and coexpressed them with RcFAH12. One isoform, RcPDATIA, increased HFA levels to 27%. Analysis of HFA-triacylglycerol molecular species and regiochemistry, along with analysis of the HFA content of phosphatidylcholine, indicates that RcPDATIA functions as a PDAT in vivo. Expression of RcFAH12 alone leads to a significant decrease in FA content of seeds. Coexpression of RcPDATIA and RcDGAT2 (for diacylglycerol acyltransferase 2) with RcFAH12 restored FA levels to nearly wild-type levels, and this was accompanied by a major increase in the mass of HFAs accumulating in the seeds. We show the usefulness of RcPDATIA for engineering plants with high levels of HFAs and alleviating bottlenecks due to the production of unusual FAs in transgenic oilseeds.
DOI:10.1111/j.1467-7652.2008.00361.xURLPMID:18643899 [本文引用: 1]
A central goal of green chemistry is to produce industrially useful fatty acids in oilseed crops. Although genes encoding suitable fatty acid-modifying enzymes are available from many wild species, progress has been limited because the expression of these genes in transgenic plants produces low yields of the desired products. For example, Ricinus communis fatty acid hydroxylase 12 (FAH12) produces a maximum of only 17% hydroxy fatty acids (HFAs) when expressed in Arabidopsis . cDNA clones encoding R. communis enzymes for additional steps in the seed oil biosynthetic pathway were identified. Expression of these cDNAs in FAH12 transgenic plants revealed that the R. communis type-2 acyl-coenzyme A:diacylglycerol acyltransferase (RcDGAT2) could increase HFAs from 17% to nearly 30%. Detailed comparisons of seed neutral lipids from the single- and double-transgenic lines indicated that RcDGAT2 substantially modified the triacylglycerol (TAG) pool, with significant increases in most of the major TAG species observed in native castor bean oil. These data suggest that RcDGAT2 prefers acyl-coenzyme A and diacylglycerol substrates containing HFAs, and biochemical analyses of RcDGAT2 expressed in yeast cells confirmed a strong preference for HFA-containing diacylglycerol substrates. Our results demonstrate that pathway engineering approaches can be used successfully to increase the yields of industrial feedstocks in plants, and that members of the DGAT2 gene family probably play a key role in this process.
DOI:10.1007/s11745-010-3385-4URLPMID:20101470 [本文引用: 1]
Triacylglycerol (TAG) is the main storage lipid in plants. Acyl-CoA: diacylglycerol acyltransferase (DGAT1 and DGAT2) and phospholipid: diacylglycerol acyltransferase (PDAT) can catalyze TAG synthesis. It is unclear how these three independent genes are regulated in developing seeds, and particularly if they have specific functions in the high accumulation of unusual fatty acids in seed oil. The expression patterns of DGAT1, DGAT2 and a PDAT in relation to the accumulation of oil and epoxy and hydroxy fatty acids in developing seeds of the plant species Vernonia galamensis, Euphorbia lagascae, Stokesia laevis and castor that accumulate high levels of these fatty acids in comparison with soybean and Arabidopsis were investigated. The expression patterns of DGAT1, DGAT2 and the PDAT are consistent with all three enzymes playing a role in the high epoxy or hydroxy fatty acid accumulation in developing seeds of these plants. PDAT and DGAT2 transcript levels are present at much higher levels in developing seeds of epoxy and hydroxy fatty acid accumulating plants than in soybeans or Arabidopsis. Moreover, PDAT, DGAT1 and DGAT2 are found to be expressed in many different plant tissues, suggesting that these enzymes may have other roles in addition to seed oil accumulation. DGAT1 appears to be a major enzyme for seed oil accumulation at least in Arabidopsis and soybeans. For the epoxy and hydroxy fatty acid accumulating plants, DGAT2 and PDAT also show expression patterns consistent with a role in the selective accumulation of these unusual fatty acids in seed oil.
DOI:10.1007/s10142-012-0306-zURLPMID:23322364 [本文引用: 1]
Diacylglycerol acyltransferase (DGAT) catalyzes the acyl-CoA-dependent acylation of sn-1,2-diacylglycerol to form seed oil triacylglycerol (TAG). To understand the features of genes encoding soybean (Glycine max) DGATs and possible roles in soybean seed oil synthesis and accumulation, two full-length cDNAs encoding type 1 diacylglycerol acyltransferases (GmDGAT1A and GmDGAT1B) were cloned from developing soybean seeds. These coding sequences share identities of 94 % and 95 % in protein and DNA sequences. The genomic architectures of GmDGAT1A and GmDGAT1B both contain 15 introns and 16 exons. Differences in the lengths of the first exon and most of the introns were found between GmDGAT1A and GmDGAT1B genomic sequences. Furthermore, detailed in silico analysis revealed a third predicted DGAT1, GmDGAT1C. GmDGAT1A and GmDGAT1B were found to have similar activity levels and substrate specificities. Oleoyl-CoA and sn-1,2-diacylglycerol were preferred substrates over vernoloyl-CoA and sn-1,2-divernoloylglycerol. Both transcripts are much more abundant in developing seeds than in other tissues including leaves, stem, roots, and flowers. Both soybean DGAT1A and DGAT1B are highly expressed at developing seed stages of maximal TAG accumulation with DGAT1B showing highest expression at somewhat later stages than DGAT1A. DGAT1A and DGAT1B show expression profiles consistent with important roles in soybean seed oil biosynthesis and accumulation.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}