 ,, 朱雅婧, 赵飞云, 王爽, 刘骕骦, 郭凌凯, 赵海红, 麻浩,南京农业大学作物遗传与种质创新国家重点实验室,南京 210095
,, 朱雅婧, 赵飞云, 王爽, 刘骕骦, 郭凌凯, 赵海红, 麻浩,南京农业大学作物遗传与种质创新国家重点实验室,南京 210095Isolation and Functional Analysis of Soybean GmLEA in Seed Vigor
ZHOU YaLi,, ZHU YaJing, ZHAO FeiYun, WANG Shuang, LIU SuShuang, GUO LingKai, ZHAO HaiHong, MA Hao,National Key Laboratory of Crop Genetics and Germplasm Enhancement, Nanjing Agricultural University, Nanjing 210095通讯作者:
第一联系人:
责任编辑: 李莉
收稿日期:2018-06-22接受日期:2018-07-29网络出版日期:2018-12-01
| 基金资助: |
Received:2018-06-22Accepted:2018-07-29Online:2018-12-01

摘要
关键词:
Abstract
Keywords:
PDF (3927KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
周亚丽, 朱雅婧, 赵飞云, 王爽, 刘骕骦, 郭凌凯, 赵海红, 麻浩. 大豆GmLEA的分离及其在种子活力中的功能分析[J]. 中国农业科学, 2018, 51(23): 4397-4408 doi:10.3864/j.issn.0578-1752.2018.23.001
ZHOU YaLi, ZHU YaJing, ZHAO FeiYun, WANG Shuang, LIU SuShuang, GUO LingKai, ZHAO HaiHong, MA Hao.
0 引言
【研究意义】大豆是世界上最重要的饲粮兼用作物,可作为植物蛋白最主要来源之一。中国南方是春大豆生产的重要区域,然而在春大豆种子发育成熟时期(R6或R7期),该地区常处于高温、多雨、高湿的季节(6月底—8月初),同时大豆籽粒大,蛋白、脂肪和水分含量高,导致种子田间劣变严重,种子活力下降[1]。因此,耐高温高湿的大豆新品种的选育尤为重要。【前人研究进展】种子活力(seed vigor)是一种以植物生理学、生物化学和遗传学为根本的种子生理学方面的一个新领域[2]。通常来说,种子活力的高低主要是由遗传因子和种子发育过程中的条件一起决定的[3]。种子成熟后,会开始走向衰老和劣变即老化,这一过程是不可逆的。种子老化与其贮藏条件密切相关,其中高温高湿的环境是引起种子老化的重要因素。一般来说,种子老化后,会造成活力降低,细胞质膜完整性破坏,内部保护酶系统活性降低,脂质过氧化加剧、有毒有害物质积累等[4]。目前,对于种子收获和储藏时种子发生劣变从而导致大豆种子活力降低的各种因素已成为研究的热点[5,6,7,8,9]。大豆在种子发育成熟期遭遇高温、多雨的天气会导致种子活力降低、营养品质和外观品质下降[3,5,10-11],高温高湿胁迫已成为造成大豆种子劣变的重要原因之一[12]。关于高温高湿胁迫后对种子活力影响生理方面的研究已取得重大进展,但关于分子机制的研究较少。近几年,关于分子机制的研究取得了一定的进展。在不抗种子田间劣变春大豆种质宁镇1号和抗种子田间劣变春大豆种质湘豆3号生理成熟期,宋利茹等对其进行模拟田间高温高湿胁迫处理,通过2-DE电泳和MALDI-TOF/TOF鉴定了33个差异蛋白,这些蛋白在贮藏蛋白、转运过程、氧化还原平衡、能量代谢、蛋白合成等代谢途径与细胞过程中发挥作用[13]。王爽[1]研究发现GmCDPKSK5在响应高温高湿胁迫中发挥重要作用,并以GmCDPKSK5为诱饵,构建高温高湿胁迫下抗种子田间劣变春大豆种质种子膜蛋白酵母双杂交文库,筛选到6个与之互作的蛋白,GmLEA是筛选到的互作蛋白之一。LEA蛋白的积累与抵抗逆境胁迫之间存在正相关的关系,普遍认为其在应对外界胁迫中发挥着重要作用[14,15]。研究表明,LEA蛋白具有广泛的功能,对ABA、低温、盐害与干旱等非生物胁迫都具有一定的抵抗作用,在保护酶活性与膜结构、结合金属离子、清除活性氧自由基等方面均发挥作用[16,17]。【本研究切入点】据报道,多种蛋白质已被证明与CDPK蛋白相互作用,并在植物生长和发育中发挥重要作用,但关于LEA蛋白与CDPK的相互作用鲜有报道,王爽[1]通过酵母双杂筛选,初步筛选到GmLEA是GmCDPKSK5的互作蛋白之一。【拟解决的关键问题】本研究克隆大豆GmLEA,并对其编码的蛋白进行亚细胞定位;分析GmLEA与GmCDPKSK5的互作情况;研究其在高温高湿胁迫下的表达水平及功能。以期为进一步深入研究GmLEA在参与大豆种子活力形成和响应非生物胁迫等方面奠定基础,从而为探明田间高温高湿胁迫下春大豆种子活力形成的机制奠定基础。1 材料与方法
1.1 试验材料
供试大豆材料为南京农业大学作物遗传与种植创新国家重点实验室麻浩教授课题组保存的抗种子田间劣变春大豆品种湘豆3号和不抗品种宁镇1号。试验所需大肠杆菌DH5α感受态细胞购自南京博尔迪生物技术公司;GV3101拟南芥遗传转化菌株购自南京百思禾生物科技有限公司;pMD19-T载体购自TaKaRa公司;植物瞬时表达载体pA7-GFP、双分子荧光互补(BiFC)载体pSAT1-nEYFP-C1与pSAT1-cEYFP- C1-B、酵母双杂交试验载体pPR3N-NubG-X、pOST1- NubG与pBT3-SUC以及植物表达载体pBI121-GUS均由麻浩教授课题组保存。1.2 总DNA和RNA的提取及cDNA的合成
以2周苗龄的大豆幼叶为材料,按照CTAB法提取DNA,参照杭州宝赛新型植物总RNA提取试剂盒说明书提取大豆各种材料的RNA,采用TaKaRa公司的RTase M-MLV试剂盒合成第一链cDNA,-20℃保存备用。1.3 大豆GmLEA的分离与进化树分析
研究所用的引物以NCBI(https://www.ncbi.nlm. nih.gov/)公布的GmLEA(GenBank:NM_001251272.1)的CDS序列及DNA序列作为模板,使用Primer Premier5.0软件设计引物(电子附表1),对湘豆3号和宁镇1号的DNA与cDNA模板进行PCR扩增。PCR体系与反应程序参照王爽[1]的研究。PCR产物经检测后回收、连接,并转化。挑取阳性克隆送上海铂尚生物技术有限公司测序,将测序正确的阳性克隆命名为T-GmLEA。通过NCBI的BLASTN分别对nr数据库和EST数据库同源性搜索,BLASTX对nr数据库进行相似性搜索;使用MEGA 6.0和DNAMAN多重比对蛋白质序列;采用MEGA 6.0的N-J算法构建进化树,BootStrap值为1 000。1.4 酵母双杂交试验
pBT3-SUC-GmCDPKSK5与pPR3N-GmLEA载体由麻浩教授课题组保存。将pBT3-SUC-GmCDPKSK5与pPR3N-GmLEA共转入酵母菌株NMY32中,四缺(SD/-Trp-Leu-His-Ade)培养基30℃培养筛选阳性克隆。pNubG-Fe65与pTSU2-APP质粒组合为阳性对照,pPR3N与pTSU2-APP质粒组合为阴性对照,pPR3N与pBT3-SUC-GmCDPKSK5质粒组合为自激活对照,pPR3N-GmLEA与pBT3-SUC为自激活对照。使用HTX高通量β-半乳糖苷酶试剂盒(Dualsystems Biotech,Swiss)测定β-半乳糖苷酶活性。1.5 GmLEA蛋白的亚细胞定位
通过设计GmLEA的ORF引物,利用限制性内切酶XhoⅠ和Sal Ⅰ对pA7空载和目的片段进行酶切,并通过酶切重组的方法将目的基因构建到pA7-GFP表达载体上,获得pA7-GmLEA-GFP重组载体。取叶脉较少的烟草叶片并剪去主脉,叶片背面朝上平贴于MS培养基,25℃暗培养。使用PDS-1000/He基因枪(Bio-Rad)将重组质粒转化到烟草叶片细胞中,25℃暗培养16 h,激光共聚焦显微镜下观察GFP蛋白表达信号。1.6 双分子荧光互补试验
通过对GmCDPKSK5和GmLEA的ORF进行引物的设计,利用限制性内切酶SalⅠ和BamHⅠ对nE-YFP和cE-YFP 2个空载和目的片段进行酶切,并通过酶切重组的方法将目的基因分别构建到nE-YFP和cE-YFP 2个载体上,共得到nE-GmCDPKSK5和cE-GmLEA 2个重组载体。采用上述1.5相同的方法将重组质粒转化到烟草叶片细胞中,25℃暗培养16 h,激光共聚焦显微镜下观察YFP蛋白表达信号。1.7 GmLEA的表达分析
将大豆种子播种于塑料花盆中,每盆约3—5粒种子;并将其置于正常条件下生长。当大豆生长至R6与R7之间时,置于人工气候培养箱中,模拟田间高温高湿环境条件进行处理,正常环境中生长植株为对照。大豆植株正常生长条件与处理条件见表1。在处理期间的0、24、48、96和168 h收集来自处理和对照植物中部的种子。当大豆生长至一节期(V1)时,采集大豆植株的子叶、根、茎及叶;花取自其盛花期(R2);幼荚和幼嫩的豆粒取自盛荚期(R4);成熟的荚和成熟的豆粒取自完熟期(R8),进行组织特异性表达分析。
当大豆植株处于盛花期(R2)时,选取植株上部的花在花柄处挂牌并标注日期,从花后15 d开始到花后60 d结束,每隔5 d取一次样,收集整个荚,进行种子发育时期分析。
所有样品取样后立即于液氮中冷冻,-80℃保存备用。参照WANG等[18]方进行法qRT-PCR分析,设置3次重复。
1.8 拟南芥转化及转基因纯合植株的获得
将GmLEA的ORF区域构建到pBI121-GUS 植物过表达载体上,该载体主要包括Kan抗性基因、CaMV35S启动子和GUS。通过农杆菌介导法转化野生型拟南芥(WT)。经筛选鉴定得到3个GmLEA过表达阳性株系LEA-1、LEA-2和LEA-3,分别收获纯合的T3代过表达拟南芥种子进行功能分析。1.9 过表达拟南芥种子生活力与活力指标的测定
按表1将WT、pBI121-GUS过表达和GmLEA过表达植株荚黄熟期的植株处理2 d,同时设置对照。Table 1
表1
表1植物生长条件
Table 1
| 类别 Class | 白天 Day | 黑夜 Dark night | 用途 Use | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 温度 Temperature (℃) | 湿度 Humidity (%) | 光照强度 Light intensity (μmol·m-2·s-1) | 时间 Time (h) | 温度 Temperature (℃) | 湿度 Humidity (%) | 光照强度 Light intensity (μmol·m-2·s-1) | 时间 Time (h) | ||
| 对照 Control | 30 | 70 | 22000 | 10 | 25 | 60 | 0 | 14 | GmLEA的表达分析Expression of GmLEA |
| 处理 Treatment | 40 | 100 | 22000 | 10 | 30 | 70 | 0 | 14 | |
| 对照 Control | 22 | 70 | 22000 | 16 | 20 | 60 | 0 | 8 | 种子生活力与活力指标的测定Determination of seed viability and vigor |
| 处理 Treatment | 40 | 100 | 22000 | 16 | 35 | 80 | 0 | 8 | |
新窗口打开|下载CSV
待种子收获后,处理组和对照组各株系拟南芥种子各取100粒,进行TTC染色,种胚全部或大部分被染成红色的为具有生活力的种子,反之为死种子,观察种子生活力。另外,分别取100粒种子点播于MS培养基上,从播种后的第1天至第5天,每半天记录一次种子的发芽数,称取第5天拟南芥幼苗重量,统计种子发芽率、发芽势、发芽指数、活力指数及平均发芽天数。
1.10 数据处理
使用软件DPS进行方差(ANOVA)分析,检测差异显著性。2 结果
2.1 GmLEA的分离与进化树分析
以湘豆3号和宁镇1号的cDNA为模板进行PCR扩增,在1 377 bp处有与预期大小相同的特异性条带,经测序发现该基因的cDNA序列在2个品种间没有差异。以湘豆3号和宁镇1号的DNA为模板,经PCR扩增得到长度为1 879 bp的DNA序列。进化树分析结果显示,按科分为4大类,分别为豆科(Leguminosae)、蝶形花科(Papilionaceae)、葫芦科(Cucurbitaceae)和小杆科(Rhabditidae),GmLEA与豆科的LEA蛋白亲缘关系比较近(图1)。图1
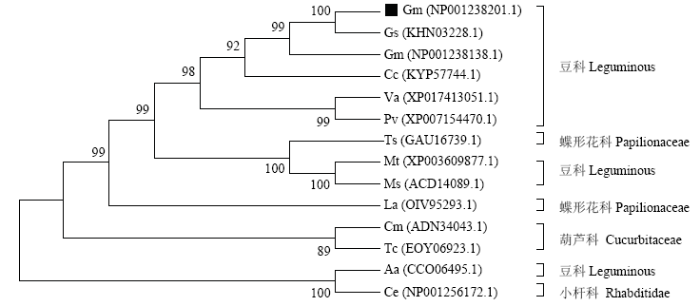 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1多个物种生物LEA蛋白的进化关系图
Gm:大豆Glycine max,NP001238201.1;Gs:野生大豆Glycine max,KHN03228.1;Cc:木豆Cajanus cajan,KYP57744.1;Va:红豆Vigna angularis,XP017413051.1;Pv:菜豆Phaseolus vulgaris,XP007154470.1;Ts:地三叶草Trifolium subterraneum,GAU16739.1;Mt:蒺藜苜蓿Medicago truncatula,XP003609877.1;Ms:紫花苜蓿Medicago sativa,ACD14089.1;La:狭叶羽扇豆Lupinus angustifolius,OIV95293.1;Cm:甜瓜Cucumis melo,ADN34043.1;Aa:小花南芥Arabis alpina,CCO06495.1;Ce:秀丽隐杆线虫Caenorhabditis elegans,NP001256172.1;蝶形花科 Papilionaceae;豆科Leguminous;葫芦科Cucurbitaceae;小杆科Rhabditidae
Fig. 1Phylogenetic tree of organism LEA protein from various organisms
2.2 GmLEA与GmCDPKSK5的酵母双杂交验证
由图2可知,全部质粒组合均在SD-TL(SD/-Trp- Leu)培养基上正常生长,说明全部质粒组合均成功转入酵母细胞,只有pNubG-Fe65与pTSU2- APP和pPR3N-GmLEA与pBT3SUC-GmCDPKSK5 2个质粒组合能够正常生长在SD-TLHA(SD/-Trp-Leu- His-Ade)培养基上且能被X-α-Gal染成蓝色,而其他3个质粒组合的酵母细胞不能在SD-TLHA培养基上生长,且pPR3N-GmLEA与pBT3SUC-GmCDPKSK5质粒组合的β-半乳糖苷酶活性值显著高于3个质粒组合,表明GmLEA与GmCDPKSK5在酵母体内存在特异性互作。图2
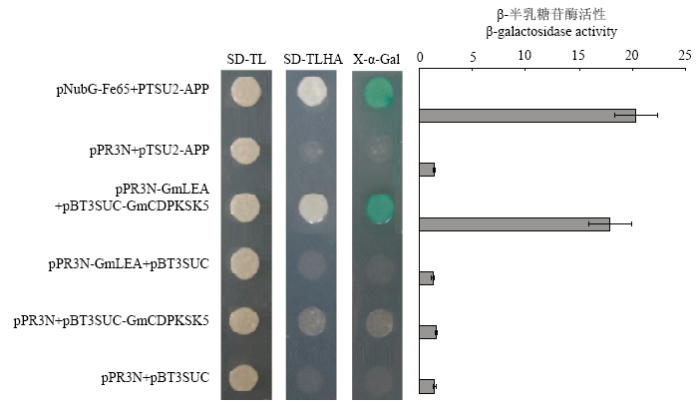 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2GmLEA与GmCDPKSK5蛋白的酵母双杂交验证
Fig. 2Identification of the interaction between GmLEA and GmCDPKSK5 through yeast two hybrid
2.3 GmLEA的亚细胞定位
采用基因枪介导法将融合表达载体共转入烟草叶片细胞中,在激光共聚焦显微镜下观察荧光位置。发现pA7-GmLEA-GFP融合蛋白在烟草叶片细胞细胞膜上表达且与膜Marker蛋白pm-ck CD3-1001信号重合,而pA7-GFP空载分布在烟草叶片细胞的整个细胞中(图3)。结果表明,GmLEA定位于烟草叶片细胞膜上。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3GmLEA在烟草叶肉细胞中的亚细胞定位
Fig. 3Subcellular localization of GmLEA protein in tobacco leaf cells
2.4 BiFC验证GmLEA与GmCDPKSK5互作
采用基因枪介导法将融合表达载体共转入烟草叶片细胞中,在激光共聚焦显微镜下观察荧光位置。将nEYFP-GmCDPKSK5与cEYFP-GmLEA质粒共转入烟草叶片细胞中,在烟草叶片细胞的细胞膜上可观察到有黄色荧光分布,并且与膜Marker蛋白pm-ck CD3-1001信号重合,而nEYFP与cEYFP-GmLEA、nEYFP-GmCDPKSK5与cEYFP和nEYFP与cEYFP质粒组合共转入烟草叶片中没有观察到黄色荧光(图4)。说明GmLEA与GmCDPKSK5蛋白主要在烟草叶片细胞的细胞膜上发生特异性互作。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4GmLEA与GmCDPKSK5蛋白在烟草叶片细胞中的BiFC验证
Fig. 4BiFC visualization of interaction between GmLEA and GmCDPKSK5 in tobacco leaf cells
2.5 GmLEA的组织表达分析
为了解GmLEA在大豆不同组织器官的表达特性,通过qRT-PCR分别检测了GmLEA在湘豆3号和宁镇1号不同组织器官中的表达情况。GmLEA主要在2个品种发育中的种子及成熟的种子中表达,在荚中有少量表达,在根、茎、叶、花和子叶中几乎无表达(图5)。图5
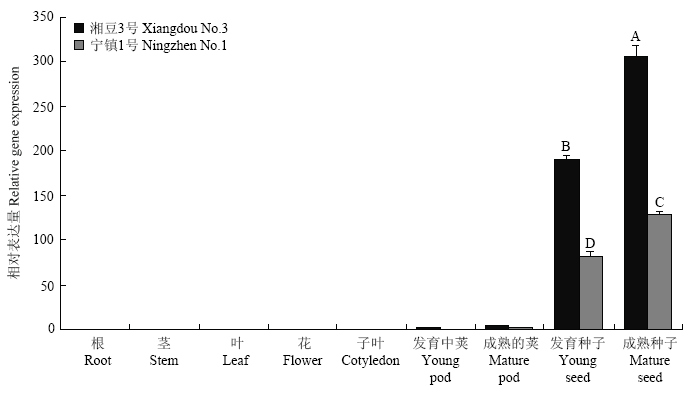 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5大豆不同组织器官中GmLEA的相对表达量
不同大写字母表示在P<0.01水平差异显著
Fig. 5The relative expression of GmLEA gene in soybean different organs
Different uppercase letters indicate significant differences at P<0.01 levels
2.6 大豆种子发育过程中GmLEA的表达特性
通过组织表达特性分析,发现GmLEA在湘 豆2号和宁镇1号种子中的表达量很高,进一步分析了该基因在2个品种种子发育过程中的表达水平。结果表明,在湘豆3号种子发育过程中GmLEA表达量先上升后下降,并且在开花后50 d达到 最大值。在宁镇1号种子发育过程中GmLEA表达量呈上升的趋势,并且在开花后60 d达到最大值(图6)。结果表明,GmLEA可能参与了种子发育过程。图6
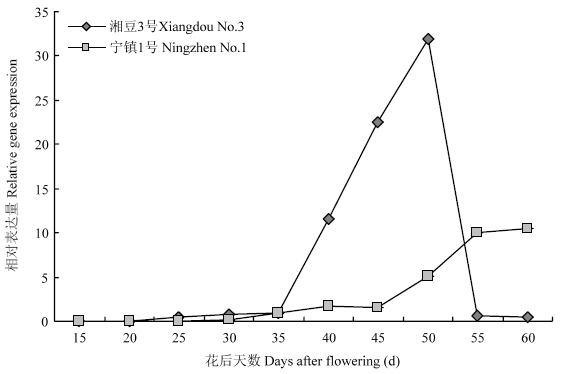 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6大豆种子发育过程中GmLEA的相对表达量
Fig. 6The relative expression of GmLEA gene in developing soybean seed
2.7 高温高湿胁迫下GmLEA的表达特性
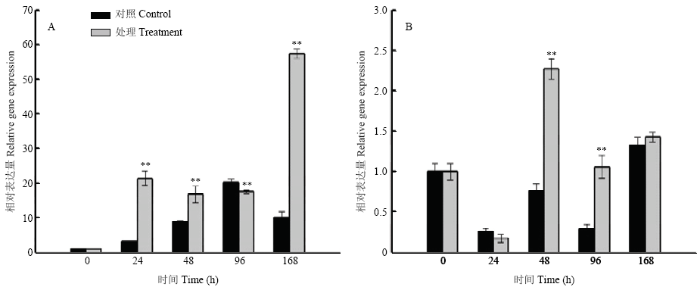
大豆植株在种子生理成熟期经高温高湿胁迫处理后,在湘豆3号种子中,GmLEA仅在96 h下调表达,在其余时间点均上调表达,且在24、48、96和168 h差异达极显著(P<0.01)水平;在宁镇1号种子中,GmLEA的表达量仅在24 h低于对照组,在其余时间点表达量均高于对照组,且在48 h和96 h差异达极显著(P<0.01)水平(图7)。结果表明,GmLEA响应高温高湿胁迫,并推测其可能参与春大豆种子田间劣变抗性过程。图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7高温高湿胁迫下GmLEA在种子中的表达
A:湘豆3号;B:宁镇1号。**表示差异极显著(P<0.01)
Fig. 7The expression patterns of the GmLEA gene in the seeds under HTH
A: Xiangdou No.3; B: Ningzhen No.1. ** indicate that the differences are markly significant at 0.01 level
2.8 高温高湿胁迫下GmLEA过表达拟南芥种子的活力分析
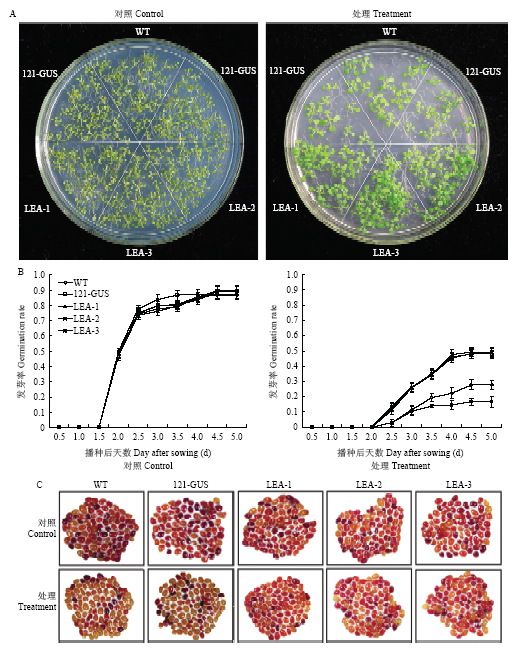
通过对种子进行高温高湿胁迫,在正常生长条件下,所有植株的种子在第5天发芽率均能达到90%左右,且都能正常生长为幼苗;而经高温高湿胁迫后,各个株系种子的发芽率均有所下降,但GmLEA过表达植株下降的幅度最小,状态要好于其他2个株系(图8-A和图8-B)。TTC染色结果与种子萌发的结果基本一致(图8-C)。表2显示高温高湿胁迫后,GmLEA过表达植株、pBI121-GUS过表达植株与WT植株的发芽势和发芽指数均显著(P<0.01)低于对照组,但是GmLEA过表达植株优于(P<0.01)其他2个株系;而WT、pBI121-GUS过表达株系和GmLEA过表达株系种子的平均发芽天数比对照组均有所提高,GmLEA过表达植株种子的平均发芽天数低于WT、pBI121-GUS过表达植株。结果表明,高温高湿胁迫后GmLEA过表达株系的种子活力显著高于野生型。图8
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8高温高湿胁迫对拟南芥种子发芽率与生活力的影响
A:播种7 d后的经高温高湿处理2 d收获的野生型和转基因株系幼苗;B:对照条件下与经高温高湿处理的种子发芽率;C:经高温高湿处理的种子TTC染色,Bar=2 mm
Fig. 8Effect on germination percentage of Arabidopsis seeds under HTH stress
A: Illustration of representative 7-day-old seedlings from the developing seeds harvested from the wild type and transgenic line plants under the control and HTH treatment for 2 d; B: Germination percentage of the developing seeds harvested from the wild type and transgenic line plants under the control and HTH stress; C: Seed viability of the developing seeds harvested from the wild type and transgenic line plants under the control and HTH treatment, Bar=2 mm
Table 2
表2
表2高温高湿处理对拟南芥种子萌发特性的影响
Table 2
| 类别 Class | 株系 Lines | 发芽势 Germination potential (%) | 发芽指数 Germination index | 活力指数 Vitality index | 平均发芽天数 Average germination days (d) |
|---|---|---|---|---|---|
| 处理 Treatment | WT | 10%±0.02D | 10.88±0.30DE | 5.52±0.25H | 4.07±+0.02AB |
| 121-GUS | 9%±0.03D | 7.51±0.39E | 4.80±0.21I | 4.02±0.03A | |
| LEA-1 | 31%±0.07C | 21.53±0.40C | 9.88±0.14FG | 3.92±0.04BC | |
| LEA-2 | 32%±0.05C | 22.62±0.41C | 10.02±0.45F | 3.91±0.06CD | |
| LEA-3 | 34%±0.05C | 20.08±0.27CD | 9.93±0.23G | 3.89±0.07BC | |
| 对照 Control | WT | 69%±0.08B | 53.37±0.32AB | 40.33±0.15E | 3.59±0.02E |
| 121-GUS | 85%±0.01A | 61.3±0.30A | 41.74±0.22D | 3.61±0.05E | |
| LEA-1 | 72%±0.05B | 51.74±0.45B | 45.18±0.13B | 3.69±0.04DE | |
| LEA-2 | 73%±0.04AB | 53.06±0.33AB | 44.97±0.21C | 3.67±+0.08E | |
| LEA-3 | 75%±0.04AB | 52.33±+0.29B | 45.77±0.22A | 3.70±0.03DE |
新窗口打开|下载CSV
3 讨论
3.1 GmLEA的表达分析
LEA蛋白是一类在生物体内广泛存在的蛋白质,它参与植物抵抗胁迫和渗透调节等过程,在植物的不同器官中都有LEA蛋白的广泛分布,不仅在高等植物种子中分布,花粉管和营养组织中也有分布,主要在种子胚胎发育后期积累[19,20,21]。有研究表明LEA在种子发育过程中逐渐积累[22]。本研究中组织特异性表达结果显示GmLEA在种子中大量表达,由此推测GmLEA可能参与种子的发育;但值得注意的是在宁镇1号种子发育该基因表达低于湘豆3号种子,通过2个品种中该基因的序列比对,发现其在2个品种间没有差异,因此,推测该基因在2个品种中差异表达可能是由于该基因在不同大豆品种的启动子序列或者上游调控基因的差异造成的,关于造成该基因转录模式差异的具体原因尚不明确,还需进一步研究中。3.2 GmLEA参与高温高湿胁迫反应
有研究表明,在干旱胁迫下,植物体内的LEA蛋白的构象会发生变化,形成α螺旋结构,可以和膜脂结合从而阻止水分的进一步丧失[23]。LIU等[24]研究发现蜡梅(Chimonanthus praecox (Linn.) Link)中过表达CpLEA5能够在一定程度上加强过表达拟南芥对低温胁迫的抵抗能力。大豆中的2个LEA蛋白,即GmPM1和GmPM9都可以通过C端的组氨酸与Cu2+和Fe3+结合,以清除羟基自由基,减少金属离子的毒害[25]。柑橘中的脱水素Cucor19能够清除羟基自由基和过氧化氢,一定程度上减轻活性氧自由基对植物造成的伤害[26]。目前关于LEA蛋白的功能已经有许多研究,但是该类蛋白在高温高湿方面的研究鲜有报道。本研究发现GmLEA在转录水平上响应高温高湿胁迫。此外,高温高湿胁迫处理后收获的GmLEA过表达植株种子的发芽率、发芽势、发芽指数、活力指数均优于WT和pBI121-GUS过表达植株,且GmLEA过表达植株种子的平均发芽天数明显缩短。因此可以推测高温高湿胁迫后过表达GmLEA可以提高拟南芥种子的活力。3.3 GmLEA与GmCDPKSK5相互作用
据报道,许多蛋白质已被证明与CDPK蛋白相互作用,并在植物生长和发育中发挥重要作用。AtCPK10与HSP1相互作用,在调节由ABA和钙离子介导的气孔运动中发挥重要作用[27]。麻浩老师课题组前期研究发现GmCDPKSK5与高温高湿胁迫下春大豆种子活力的形成相关,以GmCDPKSK5为诱饵构建高温高湿胁迫下抗种子田间劣变春大豆种子膜蛋白酵母双杂交文库,在细胞膜上初步筛选到6个与之互作的蛋白,分别为1个LEA蛋白、1个翻译控制肿瘤蛋白、1个种子成熟蛋白、1个微粒体油酸脱氢酶以及2个未知功能蛋白[28]。进一步研究发现,GmCDPKSK5与GmTCTP相互作用并在植物响应高温高湿胁迫中发挥重要作用[18]。在本研究中,通过酵母双杂交试验验证了GmLEA与GmCDPKSK5在酵母体内存在特异性互作;又通过BiFC试验对GmLEA与GmCDPKSK5的互作进行了进一步的验证,结果发现二者在烟草叶片细胞的细胞膜上发生特异性互作。有趣的是,结合实验室前期对GmCDPKSK5的研究结果[1],发现高温高湿胁迫后无论是GmLEA还是GmCDPKSK5都能够响应高温高湿的胁迫,而且表达趋势基本上一致。这种现象一种可能的解释就是GmLEA与GmCDPKSK5相互作用共同响应大豆种子发育过程中的高温高湿胁迫。
基于以上结果,推测GmLEA和GmCDPKSK5相互作用共同对高温高湿胁迫应激反应,但是GmLEA和GmCDPKSK5如何参与响应高温高湿胁迫的机制需要进一步研究。
4 结论
从大豆中扩增获得GmLEA,其cDNA全长1 377 bp,DNA序列全长1 879 bp,由2个外显子和1个内含子组成。GmLEA蛋白定位于烟草叶片细胞膜上,与GmCDPKSK5在细胞膜上特异性互作。GmLEA具有组织特异性表达,在大豆种子中表达量最高;过表达GmLEA能够提高拟南芥种子的活力。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 5]
[本文引用: 5]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
Magsci [本文引用: 2]

中国南方春大豆种子易劣变,已成为该地区发展粒用型和菜用型大豆生产的重大障碍。【目的】对中国南方春大豆种质种子劣变抗性的鉴定筛选可获得抗性种质,从而为劣变抗性机理研究、遗传育种改良和新品种选育奠定种质基础。【方法】本研究采用甲醇胁迫法和温箱蚀化法对92份中国南方春大豆地方品种和推广品种进行收获前和收获后种子劣变的鉴定。【结果】经筛选鉴定获得收获前种子劣变(田间劣变)相对较抗种质(发芽率≥80.00%、活力指数≥2.64和简易活力指数≥8.61)3个,收获后种子劣变较抗(耐贮藏)种质(发芽率≥80.00%、活力指数≥1.00和简易活力指数≥7.03)3个,双抗种质1个以及双不抗种质1个。【结论】中国南方春大豆种质收获前种子劣变抗性大于收获后种子劣变抗性,而且资源中收获前种子劣变抗性种质更为丰富;与种子活力和劣变性相关的发芽率、发芽势、发芽指数、活力指数和简易活力指数等指标间皆呈极显著正相关;宜联合采用活力指数、简易活力指数和发芽率作为种质种子劣变评价鉴定指标。
Magsci [本文引用: 2]
中国南方春大豆种子易劣变,已成为该地区发展粒用型和菜用型大豆生产的重大障碍。【目的】对中国南方春大豆种质种子劣变抗性的鉴定筛选可获得抗性种质,从而为劣变抗性机理研究、遗传育种改良和新品种选育奠定种质基础。【方法】本研究采用甲醇胁迫法和温箱蚀化法对92份中国南方春大豆地方品种和推广品种进行收获前和收获后种子劣变的鉴定。【结果】经筛选鉴定获得收获前种子劣变(田间劣变)相对较抗种质(发芽率≥80.00%、活力指数≥2.64和简易活力指数≥8.61)3个,收获后种子劣变较抗(耐贮藏)种质(发芽率≥80.00%、活力指数≥1.00和简易活力指数≥7.03)3个,双抗种质1个以及双不抗种质1个。【结论】中国南方春大豆种质收获前种子劣变抗性大于收获后种子劣变抗性,而且资源中收获前种子劣变抗性种质更为丰富;与种子活力和劣变性相关的发芽率、发芽势、发芽指数、活力指数和简易活力指数等指标间皆呈极显著正相关;宜联合采用活力指数、简易活力指数和发芽率作为种质种子劣变评价鉴定指标。
URL [本文引用: 1]
用甲醇胁迫,高温高湿等加速老化法,鉴定了10个栽培大豆和野生大豆品种的抗种子劣变性,结果表明,人工老化后,各野生大豆品种仍保持较高的种子活力,而栽培大豆的种子活力下降较快,电镜观察发现,野生大豆的种皮栅状细胞排列紧密,细胞层厚度大大高于栽培大豆,种皮发芽孔小于栽培大豆,这正是野生大豆高抗种子劣变的原因所在。
URL [本文引用: 1]
用甲醇胁迫,高温高湿等加速老化法,鉴定了10个栽培大豆和野生大豆品种的抗种子劣变性,结果表明,人工老化后,各野生大豆品种仍保持较高的种子活力,而栽培大豆的种子活力下降较快,电镜观察发现,野生大豆的种皮栅状细胞排列紧密,细胞层厚度大大高于栽培大豆,种皮发芽孔小于栽培大豆,这正是野生大豆高抗种子劣变的原因所在。
URL [本文引用: 1]
以抗种子劣变性不同的5个栽培 大豆和5个野生大豆品种为材料,比较了高温高湿(相对湿度100%,温度40℃条件下贮藏3天、7天、10天和14天及20%的甲醇浸泡2小时等5种种子 加速老化方法鉴定种手抗劣变性的效果。结果表明.高温高湿老化7~10天和20%的甲醇浸泡2小时,可鉴定出大豆种子抗劣变性。
URL [本文引用: 1]
以抗种子劣变性不同的5个栽培 大豆和5个野生大豆品种为材料,比较了高温高湿(相对湿度100%,温度40℃条件下贮藏3天、7天、10天和14天及20%的甲醇浸泡2小时等5种种子 加速老化方法鉴定种手抗劣变性的效果。结果表明.高温高湿老化7~10天和20%的甲醇浸泡2小时,可鉴定出大豆种子抗劣变性。
URL [本文引用: 1]
URL [本文引用: 1]
DOI:10.2135/cropsci2004.0029URL [本文引用: 1]
Two soybean [Glycine max (L.) Merrill] cultivars were grown in a phytotron at temperatures of 27/22, 33/28, and 38/33°C (day/night, 10 h day) during seed filling to investigate the effect oftemperature on seed germination and vigour. Some of the seeds produced at 33/28 and 38/33°C were smaller, wrinkled, and shriveled, and had poor quality. Most of these seeds would be removed during conditioning and would not affect the final quality of the seed lot. Standard germination of seeds with no visual abnormalities (i.e., normal seeds that were not wrinkled or shriveled) developing at 33/28°C was 98% or higher, but seed vigour was reduced as shown by an average accelerated ageing germination of 71% (97% at 27/22°C) and an average electrical conductivity of 8.2 mS m
[本文引用: 1]
DOI:10.3321/j.issn:1007-9084.2003.02.007URL [本文引用: 1]
利用恒温(30℃)抽真空密封贮藏研究贮藏时间对大豆种子活力和若干性状的影响。结果表明:随大豆种子贮存时间的延长,其发芽率、出苗率、株高、底荚高、主茎节数、有效荚数、单株粒重有降低的趋势,出苗期和开花期推迟,无效荚有增多的趋势,分枝数变化无明显规律性。相关分析显示,大豆种子活力与株高达极显著或显著正相关,而与出苗期和开花期达极显著或显著负相关。株高的降低主要是由于底荚高和主茎节数的减少所致。种子活力与其它性状间的相关系数,不同大豆基因型间差异显著。
DOI:10.3321/j.issn:1007-9084.2003.02.007URL [本文引用: 1]
利用恒温(30℃)抽真空密封贮藏研究贮藏时间对大豆种子活力和若干性状的影响。结果表明:随大豆种子贮存时间的延长,其发芽率、出苗率、株高、底荚高、主茎节数、有效荚数、单株粒重有降低的趋势,出苗期和开花期推迟,无效荚有增多的趋势,分枝数变化无明显规律性。相关分析显示,大豆种子活力与株高达极显著或显著正相关,而与出苗期和开花期达极显著或显著负相关。株高的降低主要是由于底荚高和主茎节数的减少所致。种子活力与其它性状间的相关系数,不同大豆基因型间差异显著。
URL [本文引用: 1]
URL [本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2015.01.03URL [本文引用: 1]
【目的】南方春大豆种子在生理成熟(R6或R7期)过程中易受高温高湿胁迫影响常会发生种子田间劣变,已经严重制约了中国南方春大豆生产和应用的发展。应用比较蛋白质组学技术,在蛋白表达水平上揭示高温高湿下南方春大豆种子田间劣变性和劣变抗性的机制,为遗传育种改良和新品种选育奠定种质基础。【方法】利用抗种子田间劣变种质湘豆3号和不抗种质宁镇1号为材料,在种子发育到生理成熟期时模拟田间高温高湿胁迫处理,运用双向电泳(2-DE)和MALDI-TOF/TOF鉴定技术研究春大豆种子蛋白质表达谱的变化。【结果】高温高湿胁迫处理和对照条件下(1、5、10、16和24 h),湘豆3号和宁镇1号大豆种子可溶性蛋白的每张2-DE重复胶上都可以检测到700多个可重复蛋白点,50个蛋白质点在处理与对照之间表达量上存在显著差异。其中有33个差异蛋白点经质谱分析成功鉴定;功能分类表明,这些成功鉴定的差异蛋白分别涉及细胞修复及防御(9%)、氧化还原平衡(12%)、蛋白合成(3%)、能量代谢(15%)、转运过程(15%)以及贮藏蛋白(31%)等代谢途径和细胞过程。此外,还有5个差异蛋白为未知功能蛋白。【结论】高温高湿胁迫下,抗性种质湘豆3号较不抗种质宁镇1号具有较强的抗氧化和细胞修复及防御能力,可能是其具有较强的抗种子田间劣变性的关键原因。
DOI:10.3864/j.issn.0578-1752.2015.01.03URL [本文引用: 1]
【目的】南方春大豆种子在生理成熟(R6或R7期)过程中易受高温高湿胁迫影响常会发生种子田间劣变,已经严重制约了中国南方春大豆生产和应用的发展。应用比较蛋白质组学技术,在蛋白表达水平上揭示高温高湿下南方春大豆种子田间劣变性和劣变抗性的机制,为遗传育种改良和新品种选育奠定种质基础。【方法】利用抗种子田间劣变种质湘豆3号和不抗种质宁镇1号为材料,在种子发育到生理成熟期时模拟田间高温高湿胁迫处理,运用双向电泳(2-DE)和MALDI-TOF/TOF鉴定技术研究春大豆种子蛋白质表达谱的变化。【结果】高温高湿胁迫处理和对照条件下(1、5、10、16和24 h),湘豆3号和宁镇1号大豆种子可溶性蛋白的每张2-DE重复胶上都可以检测到700多个可重复蛋白点,50个蛋白质点在处理与对照之间表达量上存在显著差异。其中有33个差异蛋白点经质谱分析成功鉴定;功能分类表明,这些成功鉴定的差异蛋白分别涉及细胞修复及防御(9%)、氧化还原平衡(12%)、蛋白合成(3%)、能量代谢(15%)、转运过程(15%)以及贮藏蛋白(31%)等代谢途径和细胞过程。此外,还有5个差异蛋白为未知功能蛋白。【结论】高温高湿胁迫下,抗性种质湘豆3号较不抗种质宁镇1号具有较强的抗氧化和细胞修复及防御能力,可能是其具有较强的抗种子田间劣变性的关键原因。
DOI:10.3389/fpls.2013.00190URLPMID:23805145 [本文引用: 1]
Plants are exposed to different external conditions that affect growth, development, and productivity. Water deficit is one of these adverse conditions caused by drought, salinity, and extreme temperatures. Plants have developed different responses to prevent, ameliorate or repair the damage inflicted by these stressful environments. One of these responses is the activation of a set of genes encoding a group of hydrophilic proteins that typically accumulate to high levels during seed dehydration, at the last stage of embryogenesis, hence named Late Embryogenesis Abundant (LEA) proteins. LEA proteins also accumulate in response to water limitation in vegetative tissues, and have been classified in seven groups based on their amino acid sequence similarity and on the presence of distinctive conserved motifs. These proteins are widely distributed in the plant kingdom, from ferns to angiosperms, suggesting a relevant role in the plant response to this unfavorable environmental condition. In this review, we analyzed the LEA proteins from those legumes whose complete genomes have been sequenced such asPhaseolus vulgaris, Glycine max, Medicago truncatula, Lotus japonicus, Cajanus cajan, andCicer arietinum.Considering their distinctive motifs, LEA proteins from the different groups were identified, and their sequence analysis allowed the recognition of novel legume specific motifs. Moreover, we compile their transcript accumulation patterns based on publicly available data. In spite of the limited information on these proteins in legumes, the analysis and data compiled here confirm the high correlation between their accumulation and water deficit, reinforcing their functional relevance under this detrimental conditions.
DOI:10.4161/psb.6.4.15042URL [本文引用: 1]
DOI:10.13560/j.cnki.biotech.bull.1985.2015.04.001URL [本文引用: 1]
Adverse conditions, including drought, salinity and extreme temperatures, often restrict the growth and development of plants. In plants, there exist a group of highly hydrophilic proteins, known as LEA(late embryogenesis abundant)proteins, which generally accumulate at the last stage of embryogenesis under natural conditions. It has a strong resistance to various abiotic stresses, and can respond to drought, cold, high salt and ABA signals. LEA proteins can maintain normal metabolic reactions of plants by maintaining cellular osmotic pressure, protecting the cell membrane structure and functioning as a molecular chaperone to protect other proteins. In this paper, the classification, structure, stress tolerance mechanisms of LEA protein and its application in improvement of stress tolerance in plant were reviewed.
DOI:10.13560/j.cnki.biotech.bull.1985.2015.04.001URL [本文引用: 1]
Adverse conditions, including drought, salinity and extreme temperatures, often restrict the growth and development of plants. In plants, there exist a group of highly hydrophilic proteins, known as LEA(late embryogenesis abundant)proteins, which generally accumulate at the last stage of embryogenesis under natural conditions. It has a strong resistance to various abiotic stresses, and can respond to drought, cold, high salt and ABA signals. LEA proteins can maintain normal metabolic reactions of plants by maintaining cellular osmotic pressure, protecting the cell membrane structure and functioning as a molecular chaperone to protect other proteins. In this paper, the classification, structure, stress tolerance mechanisms of LEA protein and its application in improvement of stress tolerance in plant were reviewed.
DOI:10.3864/j.issn.0578-1752.2014.19.002URLMagsci [本文引用: 1]
【目的】分析小麦LEA基因<em>TaLEA4</em>和<em>TaLEA5</em>及其编码蛋白的特征,比较它们在干旱、高盐、热和冷胁迫过程中的表达模式,探讨这两个LEA基因在小麦抗逆调控过程中的生物学功能,为其在小麦抗逆分子育种中的应用提供理论依据。<span>【方法】利用</span>RT-PCR技术克隆小麦的LEA基因,通过生物信息学方法分析克隆基因及其编码蛋白的结构特性,采用qRT-PCR技术检测克隆基因对ABA及非生物胁迫的响应模式。<span>【结果】克隆了</span>2个包含完整编码框的小麦LEA基因<em>TaLEA4</em>和<em>TaLEA5</em>,分别编码180和163个氨基酸,推断其分子量分别为18.8和16.9 kD,理论等电点分别为5.6和7.2。基因组序列分析发现,2个LEA基因中均包含1个100 bp的内含子。氨基酸序列分析发现,这两个LEA基因编码蛋白均富含极性氨基酸(约占整个氨基酸序列的71%),具有较强的亲水性。结构域分析显示,TaLEA4和TaLEA5蛋白中均包含1个典型的LEA_4(pfam:02987)保守域,属于LEA_4类蛋白。蛋白质高级结构分析显示,α-螺旋分别占TaLEA4和TaLEA5蛋白的96.7%和96.3%,并可形成弓形的空间结构;在TaLEA4中,检测到1个配体PEV(C<sub>39</sub>H<sub>78</sub>NO<sub>8</sub>P)的结合位点,而在TaLEA5中存在2个这样的配体结合位点。表达特性分析显示,2个LEA基因均可被植物激素ABA诱导而上调表达,其中,<em>TaLEA4</em>的表达水平显著高于<em>TaLEA5</em>;<em>TaLEA4</em>在干旱、高盐和高温胁迫过程中均受胁迫诱导而迅速上调表达,但<em>TaLEA5</em>却只受干旱胁迫的诱导,且其表达水平显著低于<em>TaLEA4</em>;2个LEA基因对冷胁迫均无响应;干旱和高盐胁迫过程中,<em>TaLEA4</em>在根系中的诱导表达水平显著高于叶片,而热胁迫过程中该基因在叶片中的表达水平要显著高于根系,这可能与根系直接感受渗透胁迫而叶片直接感受热胁迫有关。<span>【结论】小麦</span><em>TaLEA4</em>和<em>TaLEA5</em>均属于LEA基因家族的LEA_4亚类,具有较强的亲水性,它们属于依赖于ABA胁迫响应基因调控网络;<em>TaLEA4</em>可能在干旱、高盐和热胁迫过程中均发挥重要功能,<em>TaLEA5</em>仅参与小麦对干旱胁迫的响应,其作用要弱于<em>TaLEA4</em>。
DOI:10.3864/j.issn.0578-1752.2014.19.002URLMagsci [本文引用: 1]
【目的】分析小麦LEA基因<em>TaLEA4</em>和<em>TaLEA5</em>及其编码蛋白的特征,比较它们在干旱、高盐、热和冷胁迫过程中的表达模式,探讨这两个LEA基因在小麦抗逆调控过程中的生物学功能,为其在小麦抗逆分子育种中的应用提供理论依据。<span>【方法】利用</span>RT-PCR技术克隆小麦的LEA基因,通过生物信息学方法分析克隆基因及其编码蛋白的结构特性,采用qRT-PCR技术检测克隆基因对ABA及非生物胁迫的响应模式。<span>【结果】克隆了</span>2个包含完整编码框的小麦LEA基因<em>TaLEA4</em>和<em>TaLEA5</em>,分别编码180和163个氨基酸,推断其分子量分别为18.8和16.9 kD,理论等电点分别为5.6和7.2。基因组序列分析发现,2个LEA基因中均包含1个100 bp的内含子。氨基酸序列分析发现,这两个LEA基因编码蛋白均富含极性氨基酸(约占整个氨基酸序列的71%),具有较强的亲水性。结构域分析显示,TaLEA4和TaLEA5蛋白中均包含1个典型的LEA_4(pfam:02987)保守域,属于LEA_4类蛋白。蛋白质高级结构分析显示,α-螺旋分别占TaLEA4和TaLEA5蛋白的96.7%和96.3%,并可形成弓形的空间结构;在TaLEA4中,检测到1个配体PEV(C<sub>39</sub>H<sub>78</sub>NO<sub>8</sub>P)的结合位点,而在TaLEA5中存在2个这样的配体结合位点。表达特性分析显示,2个LEA基因均可被植物激素ABA诱导而上调表达,其中,<em>TaLEA4</em>的表达水平显著高于<em>TaLEA5</em>;<em>TaLEA4</em>在干旱、高盐和高温胁迫过程中均受胁迫诱导而迅速上调表达,但<em>TaLEA5</em>却只受干旱胁迫的诱导,且其表达水平显著低于<em>TaLEA4</em>;2个LEA基因对冷胁迫均无响应;干旱和高盐胁迫过程中,<em>TaLEA4</em>在根系中的诱导表达水平显著高于叶片,而热胁迫过程中该基因在叶片中的表达水平要显著高于根系,这可能与根系直接感受渗透胁迫而叶片直接感受热胁迫有关。<span>【结论】小麦</span><em>TaLEA4</em>和<em>TaLEA5</em>均属于LEA基因家族的LEA_4亚类,具有较强的亲水性,它们属于依赖于ABA胁迫响应基因调控网络;<em>TaLEA4</em>可能在干旱、高盐和热胁迫过程中均发挥重要功能,<em>TaLEA5</em>仅参与小麦对干旱胁迫的响应,其作用要弱于<em>TaLEA4</em>。
DOI:10.1007/s10725-017-0250-yURL [本文引用: 2]
The developing seed of soybean is susceptible to high temperature and humidity (HTH) stress, resulting in pre-harvest seed deterioration in the field. Many genes are found to respond to the stress. Based on our previous proteomics study, an HTH-responsive gene, GmCDPKSK5 , was isolated from soybean seed. GmCDPKSK5 encodes a cytoplasm- and membrane-associated protein, which belongs to Group I of the CDPK family. By yeast two-hybrid (Y2H) from soybean seed cDNA library, GmTCTP was screened as a GmCDPKSK5-interacting protein. The interaction between GmCDPKSK5 and GmTCTP was further verified using bimolecular fluorescence complementation and GST pull down assays. Expression levels of both GmCDPKSK5 and GmTCTP were induced by HTH stress in soybean seed. Our results indicated that GmCDPKSK5 and GmTCTP interact with each other and may function in responses to HTH stress in soybean developing seed.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2011.19.004URLMagsci [本文引用: 1]
【目的】鉴定大豆全基因组中的LEA家族基因,对其进行定位、分类以及组织表达分析,为植物LEA基因的功能研究与利用提供基础。【方法】利用大豆基因组数据库,通过生物信息学手段,鉴定并获得大豆LEA家族基因的全序列、定位和拷贝数信息;通过序列比对进行进化和分类分析;利用大豆发育表达芯片数据、NCBI中UniGene的EST表达数据进行组织表达谱分析。【结果】系统地分析鉴定了36个大豆的LEA家族基因,根据结构域的差异和系统发育分析将这些LEA基因分成8个亚家族。基因定位分析结果表明,这些基因分布于大豆的16条染色体上,启动子分析表明,几乎全部LEA基因的启动子区含有逆境反应顺式作用元件。各个发育阶段表达谱的分析结果表明,多数成员至少在一个组织中表达,10个差异表达的基因中有5个在种子发育时期优势表达,另外5个在其它部位优势表达。【结论】通过全基因组扫描,获得大豆基因组的36个LEA家族基因,它们分属于8个不同的亚家族,分布于16条大豆染色体上,启动子区含有逆境相关顺式作用元件,基因表达具有一定特异性。
DOI:10.3864/j.issn.0578-1752.2011.19.004URLMagsci [本文引用: 1]
【目的】鉴定大豆全基因组中的LEA家族基因,对其进行定位、分类以及组织表达分析,为植物LEA基因的功能研究与利用提供基础。【方法】利用大豆基因组数据库,通过生物信息学手段,鉴定并获得大豆LEA家族基因的全序列、定位和拷贝数信息;通过序列比对进行进化和分类分析;利用大豆发育表达芯片数据、NCBI中UniGene的EST表达数据进行组织表达谱分析。【结果】系统地分析鉴定了36个大豆的LEA家族基因,根据结构域的差异和系统发育分析将这些LEA基因分成8个亚家族。基因定位分析结果表明,这些基因分布于大豆的16条染色体上,启动子分析表明,几乎全部LEA基因的启动子区含有逆境反应顺式作用元件。各个发育阶段表达谱的分析结果表明,多数成员至少在一个组织中表达,10个差异表达的基因中有5个在种子发育时期优势表达,另外5个在其它部位优势表达。【结论】通过全基因组扫描,获得大豆基因组的36个LEA家族基因,它们分属于8个不同的亚家族,分布于16条大豆染色体上,启动子区含有逆境相关顺式作用元件,基因表达具有一定特异性。
DOI:10.1104/pp.108.120725URLPMID:18772351 [本文引用: 1]
The complete nucleotide sequences of a Korean isolate of Potato virus X (PVX-Kr) has been determined. Full-length cDNA of PVX-Kr has been directly amplified by long template reverse transcription and polymerase chain reaction (RT-PCR) using virus specific F-end primer and 3'-end primer, and then constructed in a plasmid vector. Consecutive subclones of a full-length cDNA clone were constructed to identify whole genome sequence of the virus. Total nucleotide sequences of genome of PVX-Kr were 6,435 excluding one adenine at poly A tail, and genome organization was identical with that of typical PVX species. Comparison of whole genome sequence of PVX-Kr with those of European and South American isolates showed 95.4-96.8% and 77.4-77.9%, in nucleotide similarity, respectively. Sequenced PVX-Kr in this study and twelve isolates already reported could be divided into two subgroups in phylogeny based on their complete nucleotide sequences. Phylogenetic tree analysis demonstrated that PVX-Kr was clustered with European and Asian isolates (Taiwan, os, bs, Kr, S, X3, UK3, ROTH1, Tula) in the same subgroup and South American isolates (CP, CP2, CP4, HB) were clustered
[本文引用: 1]
[本文引用: 1]
DOI:10.3390/ijms161126006URLPMID:4661864 [本文引用: 1]
Plants synthesize and accumulate a series of stress-resistance proteins to protect normal physiological activities under adverse conditions.Chimonanthus praecoxwhich blooms in freezing weather accumulates late embryogenesis abundant proteins (LEAs) in flowers, butC. praecoxLEAs are little reported. Here, we report a group of five LEA genes ofC. praecox(CpLEA5, KT727031). Prokaryotic-expressedCpLEA5was employed inEscherichia colito investigate bioactivities and membrane permeability at low-temperature. In comparison with the vacant strains,CpLEA5-containing strains survived in a 20% higher rate; and the degree of cell membrane damage inCpLEA5-containing strains was 55% of that of the vacant strains according to a conductivity test, revealing the low-temperature resistance ofCpLEA5in bacteria.CpLEA5was also expressed inPichia pastoris.Interestingly, besides low-temperature resistance,CpLEA5conferred high resistance to salt and alkali inCpLEA5overexpressing yeast. TheCpLEA5gene was transferred intoArabidopsis thalianato also demonstrateCpLEA5actions in plants. As expected, the transgenic lines were more resistant against low-temperature and drought while compared with the wild type. Taken together,CpLEA5-conferred resistances to several conditions in prokaryote and eukaryotes could have great value as a genetic technology to enhance osmotic stress and low-temperature tolerance.
DOI:10.1093/pcp/pcr052URLPMID:21531760 [本文引用: 1]
Late embryogenesis abundant (LEA) group 4 (LEA4) proteins play an important role in the water stress tolerance of plants. Although they have been hypothesized to stabilize macromolecules in stressed cells, the protective functions and mechanisms of LEA4 proteins are still not clear. In this study, the metal binding properties of two related soybean LEA4 proteins, GmPM1 and GmPM9, were tested using immobilized metal ion affinity chromatography (IMAC). The metal ions Fe(3+), Ni(2+), Cu(2+) and Zn(2+) were observed to bind these two proteins, while Ca(2+), Mg(2+) or Mn(2+) did not. Results from isothermal titration calorimetry (ITC) indicated that the binding affinity of GmPM1 for Fe(3+) was stronger than that of GmPM9. Hydroxyl radicals generated by the Fe(3+)/H(2)O(2) system were scavenged by both GmPM1 and GmPM9 in the absence or the presence of high ionic conditions (100 mM NaCl), although the scavenging activity of GmPM1 was significantly greater than that of GmPM9. These results suggest that GmPM1 and GmPM9 are metal-binding proteins which may function in reducing oxidative damage induced by abiotic stress in plants.
DOI:10.1016/j.plaphy.2004.06.004URLPMID:15331095 [本文引用: 1]
Dehydrins are ubiquitous proteins produced by plants in response to water stress. Their functions, however, are not fully understood. The overexpression of Citrus unshiu Marcov. dehydrin (CuCOR19) enhanced cold tolerance in transgenic plants by reducing lipid peroxidation promoted by cold stress, suggesting that the CuCOR19 protein directly scavenges radicals. In this paper, we report the radical scavenging activity and oxidative modification of CuCOR19. The hydroxyl radical generated by the Fe 2+/H 2O 2 system and peroxyl radical generated from 2, 2 -azobis (2-amidinopropane) (AAPH) were scavenged by CuCOR19, but hydrogen peroxide and superoxide were not. The scavenging activity for the hydroxyl radical and peroxyl radical of CuCOR19 was more potent than that of mannitol, and approximately equal to that of serum albumin, which is known as an antioxidative protein in mammals. CuCOR19 was degraded by the hydroxyl radical and peroxyl radical in a time- and dose-dependent manner. Mannitol and thiourea inhibited the degradation. Analysis of the amino acid composition of CuCOR19 indicated that glycine, histidine, and lysine, which are major residues in many dehydrins, were targeted by the hydroxyl radical. These results suggest that CuCOR19 is a radical scavenging protein, and may reduce oxidative damage induced by water stress in plants.
[本文引用: 1]
DOI:10.7685/jnau.201701036URL [本文引用: 1]
钙是植物必需的大量元素之一,同时钙离子(Ca2+)是细胞信号转导的第二信使.钙依赖蛋白激酶(calcium-dependent protein kinase,CDPK)作为Ca2+的感受器普遍存在于植物和部分原生动物中,是植物特有的一类丝氨酸/苏氨酸型蛋白激酶,参与了多种Ca2+介导的信号通路,在植物发育信号和逆境信号转导中具有重要作用.本文概述了钙依赖蛋白激酶在植物体内的分布、结构、底物、功能以及大豆CDPK的研究进展,旨在为今后培育抗逆植物品种提供参考依据.
DOI:10.7685/jnau.201701036URL [本文引用: 1]
钙是植物必需的大量元素之一,同时钙离子(Ca2+)是细胞信号转导的第二信使.钙依赖蛋白激酶(calcium-dependent protein kinase,CDPK)作为Ca2+的感受器普遍存在于植物和部分原生动物中,是植物特有的一类丝氨酸/苏氨酸型蛋白激酶,参与了多种Ca2+介导的信号通路,在植物发育信号和逆境信号转导中具有重要作用.本文概述了钙依赖蛋白激酶在植物体内的分布、结构、底物、功能以及大豆CDPK的研究进展,旨在为今后培育抗逆植物品种提供参考依据.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}