 ,, ������,, ���, ���, ���«h, ������,����ũҵ��ѧ�����뻷��ѧԺ/����������Դ��Ч���ù��ҹ���ʵ����,���� 110866
,, ������,, ���, ���, ���«h, ������,����ũҵ��ѧ�����뻷��ѧԺ/����������Դ��Ч���ù��ҹ���ʵ����,���� 110866Influence of Long-term Fertilization on Community Structures and Colonization of Arbuscular mycorrhizal Fungi in a Brown Soil
GUO Jing,, LUO PeiYu,, YANG JinFeng, LI DongDong, HUANG YueYue, HAN XiaoRi,College of Land and Environment, Shenyang Agricultural University/National Engineering Laboratory for Efficient Utilization of Soil and Fertilizer Resources, Shenyang 110866ͨѶ����:
��һ��ϵ��:
�ո�����:2018-04-16��������:2018-07-10�����������:2018-12-26
| ��������: |
Received:2018-04-16Accepted:2018-07-10Online:2018-12-26

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (1063KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
����, ������, ���, ���, ���«h, ������. ����ʩ�ʶ�������֦�������Ⱥ��ṹ������Ⱦ��Ӱ��[J]. �й�ũҵ��ѧ, 2018, 51(24): 4677-4689 doi:10.3864/j.issn.0578-1752.2018.24.008
GUO Jing, LUO PeiYu, YANG JinFeng, LI DongDong, HUANG YueYue, HAN XiaoRi.
0 ����
���о����塿��֦���� ��Arbuscular mycorrhiza,AM�������һ��������������ֲ��Ӫ����ϵ���е���Ҫ����,������������ֲ���ϵ�γɹ�����ϵ[1,2],����������ϵ���շ�Χ�����ձ�������ٽ���������������п���Ԫ�غ�ˮ�ֵ�����[3,4,5,6]������ֲ��ˮ�ִ�л����ǿֲ����ԡ�������������������������״[7]�����ȶ���̬ϵͳ��������̬ƽ��Ϳɳ�����չ,�ٽ�ũ���֡���ҵ�������в�������ľ��á�������̬���塣��ǰ���о���չ��Ȼ��,����AM������ֶ����ԡ��Լ��������ѡ���ԺͶ�������������Ӧ�Բ�ͬ[8],���� AM ����ٽ�ֲ���ڲ�ͬ�����¶�ֳ��������Ҫ����������ص�Ӱ�졣���о�֤ʵ��ͬ���������͡�ֲ�����͡������ƶȡ�ʩ�ʴ�ʩ�Ȼ��AM��������ԡ������ܶȡ���Ⱦ��������Ӱ��[9,10,11,12,13];��Ŀǰ��AM���Ӱ�����ص��о��������һ��,���о���������ʩ�л��ʶ�AM����Ķ����ԡ���Ⱦ�ʺ������ܶ��л�������[14,15],��JONER[16]�о�����ʩ�л��ʷ���������AM���������,�������[17]����AM�������Ⱦ����������������������ԡ�MUCHANE��[18]�о�������������ʩ���������������AM����Ķ�ֳ,�����廪��[12]����AM�������Ⱦ�������������ʼ����������,������������������������͡�ֲ�������ʩ�ʴ�ʩ�IJ�ͬ����ġ������о�����㡿����AM������ܵ��������ص�Ӱ��,����Բ�ͬ���������͡�ũҵ��ʩ����Ӧ������ͬ�����,̽����ͬ������AM�����Ӱ�����ء��������Ĺؼ����⡿���о���38��������������ʩ�ʵĻ�����,��������AM���Ⱥ��ṹ����ϵ AM�����Ⱦ����Ӱ������,̽����������������ʺ�AM���֮��������,����Ϊ���������������� AM �����Դ�����ݡ�1 �����뷽��
1.1 ����ſ�
����������ֲ����Ʒ��������ũҵ��ѧ�������ϳ��ڶ�λ����أ���γ40��48��,����123��33�䣩,�������ʼ��1979��,��2016������38����ʷ,��������-����-�������ƶ�,2016����ֲ���ס�������صĻ����ſ�������[19]��������ѡȡ����6��ʩ�ʴ�������1����ʩ�ʴ�����CK��;��2����ʩ��ѧ���ʣ�N��;��3��ʩ�û�ѧ���ʣ�NP��;��4��ʩ�û�ѧ���طʣ�NPK��;��5����ʩ�л��ʣ�M��;��6���л��ʺͻ�ѧ������ʩ��MNP�������з�����Ϊ�����ڲ�ǰһ����ʩ��,�л���Ϊ���Ƿ�,ƽ�����л���119.6 g��kg-1,ȫ�� 5.6 g��kg-1,P2O5 8.3 g��kg-1,K2O 10.9 g��kg-1;��ѧ���طʷֱ�Ϊ���ء�������ƺ������,���Ͼ���ʩ����������[19]��1.2 ��Ʒ�ɼ�������
������ֲ����Ʒ��2016���������ڣ�6��6�գ����вɼ�����������������ѡȡ12����,�������Ϊ0��20 cm,ÿ3�����ֻ���,����ʯ��ֲ���ϵ������,��1 mmɸ,һ���ֲⶨ�������л�̼���̬������̬����������ˮ������ȡ������DNA;��һ���ֱܹ��ɺ�,�ⶨ����pH��������Ч�ء���Ч�������ܶȡ���ȡ4������������ϵ,�ô�������ˮ��ϴ�ɾ�����������ij���ˮ��ϴ,��ϴ�����ʸ�����1 cm���ĸ���,������ȡDNA���ⶨ��Ⱦ�ʡ�1.3 �ⶨ����
�������л�̼��K2SO4���ᷨ,����ȡ10 g��������50 mL 0.5 mol��L-1 K2SO4��Һ��1 h,Ȼ����4 000��g������������10 min,����Һ��0.45 ��m����Ĥ����,��Һ��Vario EL II ��TOC�ǽ��вⶨ[20]���̬������̬���ĺ�������0.01 mol��L-1 CaCl2���ᷨ,�����������������ǣ�AutoAnalyzer3,�¹�Seal��˾���ϻ��ⶨ��pH��AP��AK��AHN��SM��������ũ���������������ⶨ[21]��AM��������ܶȲ���ʪɸ������,��20 g ���������ɸȡ���Ӳ����������ܶ�ͳ��[22],��Ⱦ�ʲ���̨����Ⱦɫ-�����߷��ⶨ[23]������DNA����Powersoil? DNA isolation kit�Լ��У�MO BIO, Laboratories Inc,German��������ȡ,ֲ��DNA����DNeasy? Plant Mini Kit�Լ��У�Qiage,German��������ȡ,��ȡ���DNA����-20�������á�
1.4 Ŀ��Ƭ�ε�������DGGE��Ӿ
AM�����Ŀ�������������ó�ʽPCR[24],��һ�ַ�Ӧ��ϵΪ10��PCR Buffer������Mg2+��5 ��L,dNTP 4 ��L��2.5 mmol��L-1��,MgCl2 3 ��L��2.5 mmol��L-1��,NS1��10 mmol��L-1��1.5 ��L,NS41��10 mmol��L-1��1.5 ��L,Taqø��2 U����L-1��1 ��L,DNAģ��1 ��L,��ddH2O��50 ��L���ڶ��ַ�Ӧ��ϵ��һ����ͬ��Ϊ�����DGGE�ķֱ���,�ڵڶ���PCR��������ǰ���˸�GC�У�CGCCCGCCGCGCCCCGCGCCCGTCCGCC GCCCCCGCCCG��������AM1��NS31��������ԭ��ù�ƣ�Archaeosporaceae����������ù�ƣ�Paraglomaceae����AM���,�����������һ������ARCH1311��NS8������ԭ��ù�ƺ�������ù�ơ����о���DGGE��Ӿ���õľ۱�ϩ����������Ũ��Ϊ6%,���Լ���Χ��30%��50%,��Ӿ����Һ��1��TAE, ��Ӿ��ѹ��60 V,��Ӿʱ�䣺16 h��PCR������������1��ʾ��Table 1
��1
��1AM���Ŀ������PCR��Ӧ����
Table 1
| ��������Primer | �������У�5��-3�䣩Primer sequence��5��-3�䣩 | ��Ӧ����Conditions |
|---|---|---|
| NS1 | GTAGTCATATGCTTGTCTC | 94��Ԥ����3 min;94�� 1 min,50 �� 1 min,72 �� 1 min,30��ѭ��;72 �� 7 min (һ��) An initial denaturation of 94��for 3 min; 94�� 1 min, 50�� 1 min, 72�� 1 min, 30 cycles; 72�� 7 min (The first round) |
| NS41 | CCCGTGTTGAGTCAAATTA | |
| GC-NS31 | TTGGAGGGCAAGTCTGGTGCC | |
| AM1 | GTTTCCCGTAAGGCGCCGAA | 94 ��Ԥ����3 min;94 �� 30 s,58 �� 45 s,72 �� 45 s,28��ѭ��;72 �� 7 min�����֣� An initial denaturation of 94�� for 3 min; 94�� 30 s, 58�� 45 s, 72�� 45 s, 28 cycles; 72�� 7 min (The second round) |
| GC-ARCH1311 | TGCTAAATAGCCAGGCTGY | |
| NS8 | TCCGCAGGTTCACCTACGGA |
�´��ڴ�|����CSV
1.5 DGGE����ղ���
��DGGEͼ������ȡ���������д����Ե�����,���þ۱�ϩ��������DNA�����Լ��У�������ά������Ŀ�Ļ���Ļ��ա����պ��DNA��AM���������������NS31��AM1����PCR������֤,Ȼ���ɱ���������ʢ���\���������ι�˾���п�¡����1.6 ���ݷ���������
ͼ����������Microsoft Office Excel 2010;����SPSS 19.00���з����������С����������,LSD��;���������Redundancy Analysis, RDA�������Ͷ�Ӧ������Canonical Correlation Analysis, CCA������CANOCO 4.5����;DGGEͼ��������Quantity one 4.6����;DNA���бȶ���Gen Bank ���ݿ�� BLAST �����н�������������,������Ŀ��������Ե��ϵ���������,Ȼ�����MEGA 6.0������������������ָ������ Shannon-Wiener ָ����H���;��ȶ�ָ����E����ʾ, Shannonָ����H�����㹫ʽΪH=-��(PilnPi),���ȶ�ָ����E�����㹫ʽΪE=H/lnS��ʽ��,Pi��ʾ��i����ռ����Ӿ�������ܹ��ܶ�ֵ�ı���,S��ʾÿһ��Ӿ��AM���������������2 ���
2.1 ���ڶ�λʩ�ʶ������������ʵ�Ӱ��
����2���Կ���,����ʩ�������ı�����������������,ʩ�ʴ���������AP��AHN��NO3--N��NH4+-N��DOC��SM���Ȳ�ʩ�ʶ�����������,������Ϊ���л��ʴ��������ʴ�����CK;�������������,MNP��������AHN��NO3--N��NH4+-N��AP��AK��DOC������ߡ�M����������pH���,Ϊ6.64;N������pH���,Ϊ4.87;�벻ʩ�ʴ������,ʩ���л��������������pH,�����ʻ�ʹ����pH����,�����ǵ�ʩ��ѧ���ʡ�Table 2
��2
��2������������������
Table 2
| ���� Treatment | pH (H2O) | ��Ч�� AP (mg��kg-1) | ��Ч�� AK (mg��kg-1) | �� AHN (mg��kg-1) | ��̬�� NO3--N (mg��kg-1) | �̬�� NH4+-N (mg��kg-1) | �������л�̼ DOC (mg��kg-1) | ������ˮ�� SM (%) |
|---|---|---|---|---|---|---|---|---|
| CK | 5.75��0.01b | 1.19��0.64d | 96.66��1.21c | 85.04��0.68f | 12.06��0.02e | 3.97��0.18d | 39.42��0.26e | 15.40��0.14b |
| N | 4.87��0.00d | 2.14��0.64d | 81.62��1.86c | 110.43��0.94d | 48.38��1.03c | 4.45��0.12cd | 65.41��0.05d | 15.41��0.22b |
| NP | 5.35��0.01c | 16.15��0.22c | 76.94��2.32c | 96.60��1.17e | 51.22��1.39bc | 4.83��0.16bc | 71.29��0.74c | 15.12��0.38b |
| NPK | 4.99��0.01c | 17.10��0.63c | 107.02��1.16c | 120.73��2.43c | 55.97��0.74b | 5.26��0.046b | 73.10��0.17c | 15.80��0.04b |
| M | 6.64��0.06a | 137.72��2.29b | 209.96��9.98b | 132.54��1.74b | 41.81��0.75d | 5.33��0.09b | 108.44��2.51b | 18.11��0.04a |
| MNP | 5.78��0.02b | 182.15��7.01a | 268.12��9.75a | 165.56��2.24a | 73.63��0.43a | 7.75��0.12a | 126.45��1.30a | 18.20��0.73a |
�´��ڴ�|����CSV
2.2 ����ʩ�ʶ�AM���Ⱥ��ṹ��Ӱ��
��ͼ1��ʾ,NP����������AM�����������ࣨ9����,CK��N������M������������֮��7��8����,��NPK������MNP�������������٣�6����,˵��ʩ�û�ѧ�طʼ�ʩ�õĺ����ϲ�����������������AM��������ࡣband 16��band 17��band18��band 19��band 22���л��ʴ������е�����,˵��ʩ���л��ʴٽ����⼸��AM���������;band 20������ʩ�ʴ�����������,˵��ʩ��û�жԴ���AM������Ӱ�졣ͼ1
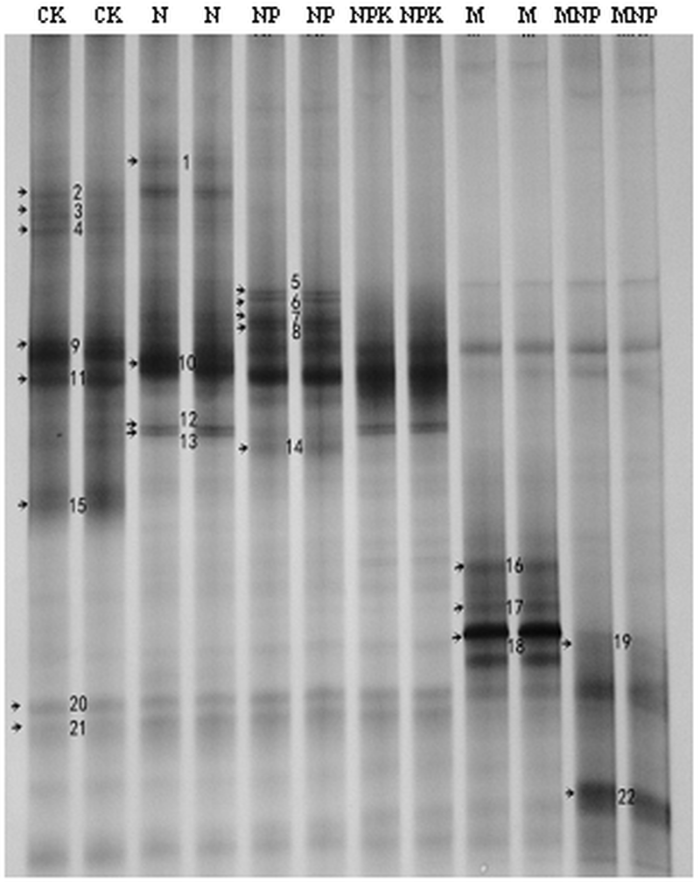 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ1��ͬʩ�ʴ�������AM�����DGGEͼ��
Fig. 1DGGE banding patterns of soil AM fungi under different fertilizer treatments
��ͼ2��ʾ,NP��NPK��M�����ĸ�ϵAM������������϶�,��CK��N��MNP����������������;��ϵband 8�����д������е�����,û����ʩ�ʵ�Ӱ��;��ϵband 5��MNP������������,˵���л����뻯ѧ������ʩ�ٽ��˸�AM�������Ⱦ����ͼ1��ͼ2���Կ���,��������һ���������22������,�������з������������ֻ��9��,˵��ֻ�в���AM �������Ⱦ����ϵ��
ͼ2
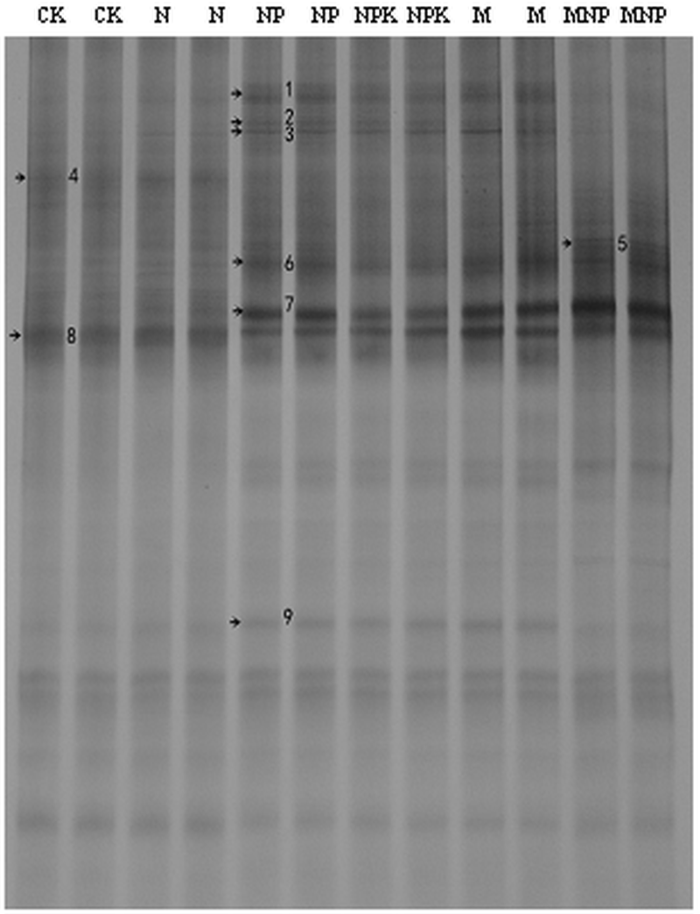 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ2��ͬʩ�ʴ�����ϵAM�����DGGEͼ��
Fig. 2DGGE banding patterns of infected AM fungi under different fertilizer treatments
��������ϵAM�����DGGEͼ���������������и����,����¡������31������,��������AM�����22������,��ϵAM�����9�����С�ͨ��GenBank���ݿ��BLAST ����������бȶ�,Ѱ��������Ե��ϵ��Ϊ�ӽ������ֹ���������,�����ͼ3��ʾ,sband 11��sband 15��sband 18��sband 19���ھ�����ù�ƣ�Uncultured Gigasporaceae��,��������������������ù,��ϵAM�������������ù,˵�������е�AM���ֻ������ù��Ⱦ������ϵ,��������ù��δ��Ⱦ����ϵ��
����3��֪,NP����������AM����Ķ�����ָ������������������,��NPK�����Ķ�����ָ�����,˵����ѧ�ʵ�ʩ���������������AM����Ķ�����,����ѧ�طʵ�ʩ����������AM����Ķ�����;����MNP������Ȼδʩ�뻯ѧ�ط�,����AM���������ָ����NPK��������������,����������NP������M����,�������MNP������ˮƽ�����йء�������������AM������ȶ�����������,˵����ͬʩ�ʶ�����AM����ľ��ȶ�û��Ӱ�졣
Table 3
��3
��3��ͬʩ�ʴ�������AM���������ָ��
Table 3
| ���� Treatment | ��ũָ�� Shannon-Wiener(H) | ���ȶ� Evenness (E) | ������ Bands |
|---|---|---|---|
| CK | 2.04��0.01c | 0.982��0.00a | 8 |
| N | 1.93��0.01d | 0.988��0.01a | 7 |
| NP | 2.18��0.02a | 0.991��0.01a | 9 |
| NPK | 1.78��0.03e | 0.997��0.02a | 6 |
| M | 2.09��0.05b | 0.999��0.01a | 8 |
| MNP | 1.78��0.02e | 0.995��0.01a | 6 |
�´��ڴ�|����CSV
����4��֪,NP��NPK��M������ϵAM���������ָ������������������,˵����ʩ�û�ѧ�ʻ��л����뻯ѧ����ʩ���ɽ�����Ⱦ����ϵ��AM���������;��CK��N������ϵAM����ľ��ȶȾ�����������������,˵������ȱ����Ⱦ����ϵ�ľ��ȶ��и���Ӱ�졣
Table 4
��4
��4��ͬʩ�ʴ�����ϵAM���������ָ��
Table 4
| ���� Treatment | ��ũָ�� Shannon-Wiener(H) | ���ȶ� Evenness(E) | ������ Bands |
|---|---|---|---|
| CK | 0.88��0.06c | 0.799��0.02b | 3 |
| N | 1.03��0.09c | 0.740��0.01b | 4 |
| NP | 1.91��0.03a | 0.981��0.01a | 7 |
| NPK | 1.92��0.02a | 0.985��0.01a | 7 |
| M | 1.91��0.01a | 0.984��0.01a | 7 |
| MNP | 1.36��0.01b | 0.988��0.01a | 4 |
�´��ڴ�|����CSV
ͼ3
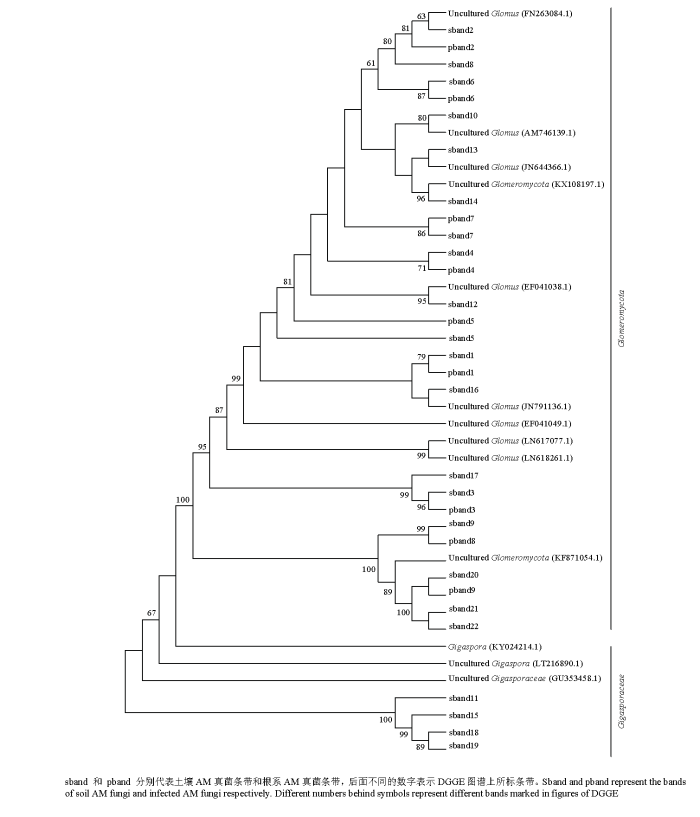 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ3��ͬʩ�ʴ���AM���ϵͳ������
Fig. 3Phylogenetic tree of AM fungi with different fertilizer treatments
2.3 ����ʩ�ʶ�AM��������ܶȼ���Ⱦ�ʵ�Ӱ��
��ͼ4���Կ���,ʩ���л��ʴ����������ܶ��������ڵ�ʩ���ʴ����Ͳ�ʩ�ʴ���,������Ϊ���л��ʴ���>���ʴ���>CK,����MNP�����������ܶ����,Ϊ37.8��/g,��CK�����������ܶ�����,Ϊ19.8��/g;ʩ�û��ʴ����������ܶ��������������֮��,Ϊ26.2��35.6��/g�������ʩ��������AM������ӵIJ���,�����л��ʵ�ʩ�ö�����AM������߾��л���Ӱ�졣��ͼ5���Կ���,ʩ������������AM�������Ⱦ��ʩ�ʴ�����AM�����Ⱦ���������ڲ�ʩ�ʴ���,����NPK��������Ⱦ�����,Ϊ25.77 %;NPK��M��MNP��������Ⱦ��������������ʩ�ʴ���,˵��ʩ�ú��ط����ܴٽ�AM���������ϵ����Ⱦ,�һ�ѧ�ط�Ч�������л��ʡ�
2.4 ����ʩ�ʶ�AM���Ⱥ��ṹ��Ӱ��
��ͼ6��ʾ,����ʩ�ʽ�����AM�����Ϊ������Ⱥ,��ʩ���ʴ�����ʩ���л��ʴ�������������������,N��������������Ⱥ�����ƶȽ�Ϊ7%,˵�����ڵ�ʩ��ѧ���ʼ���ظı�������AM���Ⱥ��ṹ;ʩ���л��ʴ����������������ƶ�Ϊ18%,˵������ʩ���л��ʶ�����AM���Ⱥ��ṹҲ�нϴ�Ӱ��;��CK������NPK��NP֮������ƶȴﵽ52%,˵��������ʩ������AM ���Ⱥ��ṹ������Ӱ���С����ͼ7��֪,����ʩ�ʽ���ϵAM�����Ϊ������Ⱥ��NPK����������ʩ�ʴ���֮������ƶ�Ϊ66%,������Ϊһ����Ⱥ,˵��ʩ�û�ѧ�طʶ�AM�����Ⱦ�����˽ϴ�Ӱ��;M������NP������Ϊһ����Ⱥ,��MNP������N������CK������Ϊһ����Ⱥ,˵��������Ч����������;����AM�����Ⱦ����Ӱ�졣
ͼ4
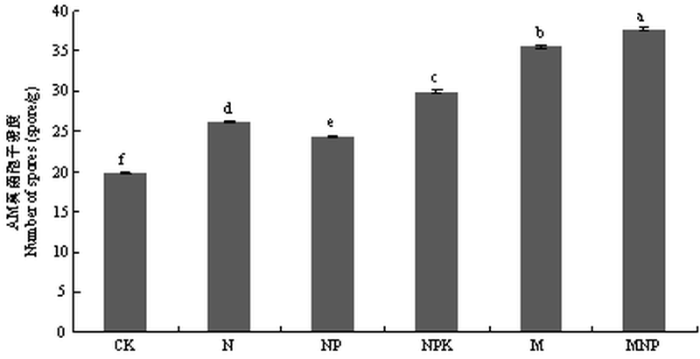 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ4��ͬʩ�ʴ���AM����������ܶ�
Fig. 4The spore density of AM fungi under different fertilizer treatments
ͼ5
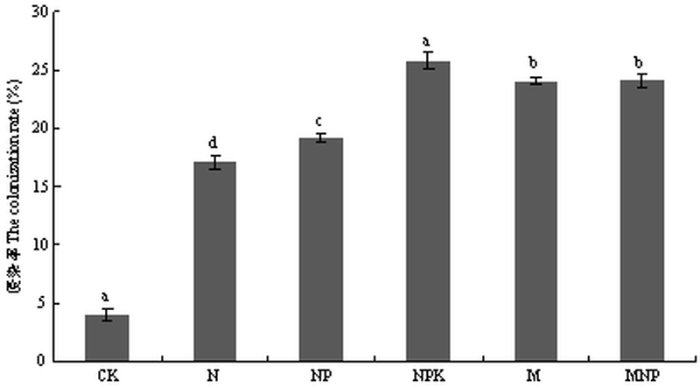 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ5��ͬʩ�ʴ���AM�������Ⱦ��
Fig. 5The colonization rate of AM fungi under different fertilizer treatments
ͼ6
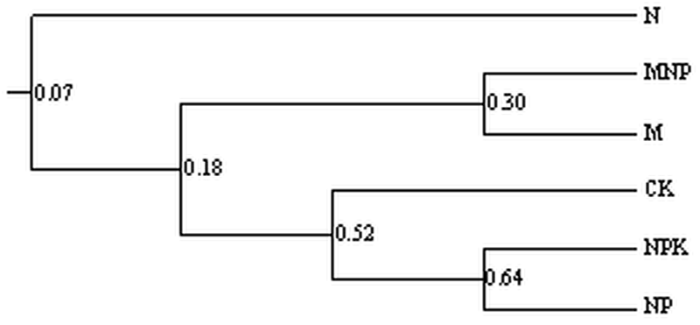 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ6��ͬʩ�ʴ�������AM��� UPGMA����ͼ��
Fig. 6UPGMA dendrogram of soil AM fungi under different fertilizer treatments��
ͼ7
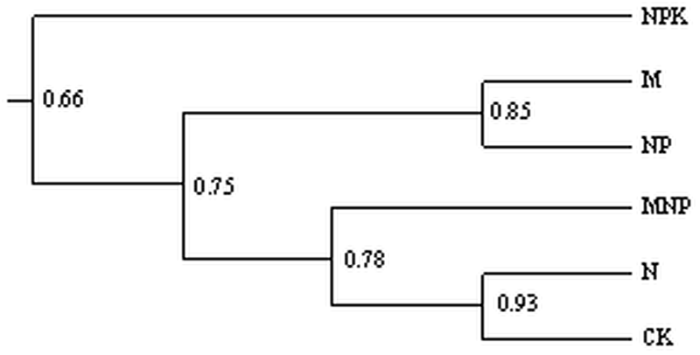 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ7��ͬʩ�ʴ�����ϵAM��� UPGMA����ͼ��
Fig. 7UPGMA dendrogram of infected AM fungi under different fertilizer treatments��
2.5 AM���Ⱥ��ṹӰ������
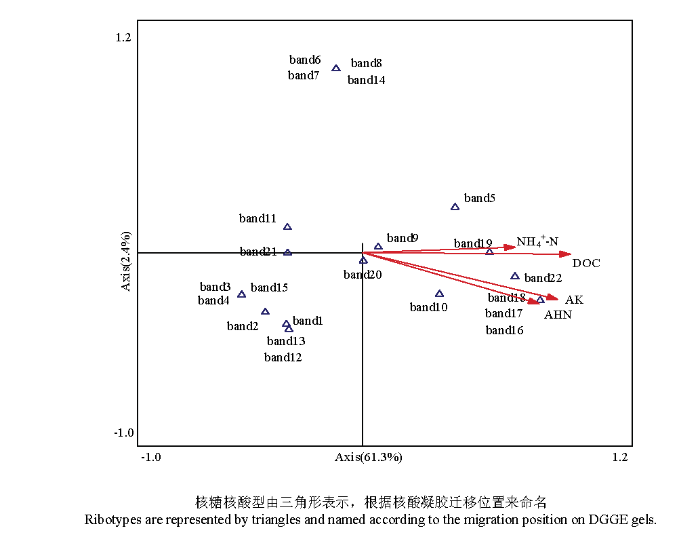
����������������ͼ8��,���ڶ�λʩ��������,��������ϵ��Ⱦ��AM���������ָ�����������������������,���������ܶ���AHN��NH4+-N��AP��AK��DOC��SM�����������,��Ⱦ������������NO3--N����������ء���Ⱦ��������AM����������ܶ�֮���ɵ��Ͷ�Ӧ������֪,������AHN��AK��DOC��NH4+-N��Ӱ������AM���Ⱥ��ṹ����Ҫ���أ�ͼ9��,��һ�͵ڶ������������63.7%�ı�����P<0.05,ͨ�����ؿ����飩������band 5��band 10��band 16 ��band 17��band 18��band 19��band 22��������AM�����������AHN��AK��DOC��NH4+-N�����������;��band 1��band 2��band 3 ��band 4��band 11��band 12��band 13��band 15��band 21��������AM�����AHN��AK��DOC��NH4+-N�����������,����������������AM�������������������������ԡ�ͼ8
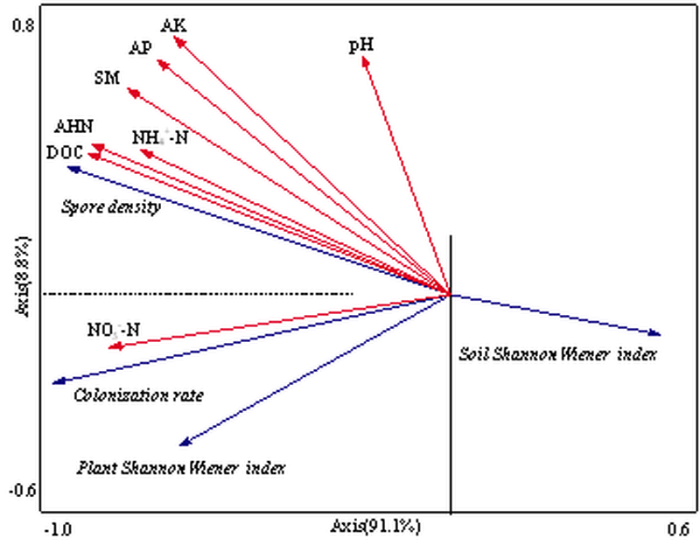 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ8AM����������������ʵ��������
Fig. 8Redundancy analysis depicting the relationship among soil physicochemical properties and AM fungi
ͼ9
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ9AM������Ǻ������������������ʵĵ��Ͷ�Ӧ����
Fig. 9Canonical Correlation analysis depicting the relationship among AM fungi ribotypes and physicochemical properties
3 ����
AM�����Ϊ��Ȼ��̬ϵͳ����ɲ���,�ܵ���ֲ���������ϵ,����̬ϵͳ������Ҫ����,���,�о���������Ⱦ��AM���Ⱥ��ṹ����Ӱ�����ؾ�����ʵ���塣���о��� AM�����DGGEͼ���д����������Ǩ��λ�á������������ڲ�ͬʩ�ʴ������в���,��˵�����ڲ�ͬʩ�ʶ�����AM���������������Ӱ�졣����ʩ�û�ѧ�طʼ�ʩ�ú����ϲ���������ή������AM����Ķ�����,����HARLEY��[25]���о����һ�¡�ֵ��ע�����,��ȻNPK��������AM���������ָ�����,�����ϵ��Ⱦ��AM���������ָ������Ⱦ�ʾ�Ϊ���,MNP��M������֮,�����������ʩ�ú��ط����ܴٽ�AM����������ϵ����Ⱦ[26],��ʩ�û�ѧ�ط������ʩ���л��ʸ�������AM�������Ⱦ�� CK��N������MNP������ϵAM���������ָ����������������ʩ�ʴ���,�����������������ˮƽ������;���Ӱ����Ⱦ����ϵ��AM��������ԡ������յ�[27]�о�����AM�������Ⱦ�����紺���,���ʼ����,�����^�ﵽ����,���о��и�ʩ�ʴ���AM�������Ⱦ�ʾ��ϵͣ�4.02%��25.77%��,���������AM���������������Ⱦ��������ǿ,��������������������,������յ��о�������ơ�BHADALUNG��[28]��27�곤��ʩ�ʴ�����AM��������Խ����˵���,���ֳ���ʩ�ʻή��AM����Ķ�����,��һЩ�о�����AM�����������������Ч�׳�����ع�ϵ[29],�����о���AM������������������������������,���ֲ������������AM�����ʩ������������ֲ��ͻ���������ѡ���Ժ���Ӧ������ͬ,����������е���ʷԭ��,���AM����������������ʵ������Բ�ͬ[30]��������о������ر�����ù���������ù����������ù����������ù�Լ�����AM������ܹ��ɹ���Ⱦ����ϵ[31],˵��AM�������������֮��û�����Ե�רһ�ԡ����о������з����AM�����ҪΪ����ù���;�����ù��,������ϵ��Ⱦ��AM���ֻ������ù��,����������������ʩ�ʹ������������͡��������������Ⱦ��ϵ��AM�����������Ⱥ��ͬ����[32]�����о��������ܶ�������AHN��NH4+-N��AP��AK��DOC��������ˮ�������������,���С����[33]�о����һ��,�����������ʩ�������������������̬���ֺ���,�Ӷ��������������������[34]��AM�����Ϊר�Թ�����,����Ⱦ�ʻ������������״���IJ�ͬ�����仯[35],���о�����һ����Χ��AM�������Ⱦ������������Ч�׳����������[36]�����о��з�����Ⱦ����������Ч���������,���������ʩ���л��ʴ�����������Ч�������߷���������AM�������Ⱦ,����Ⱦ�ʽ���������NO3--N�����������,���������Ϊ��̬������Խ��,ֲ���ϵ�������þ�Խ��ʢ[37],��ֲ���ϵ�������AM�����ֲ���γɹ�����ϵ������Ҫ������[38]��
Ф��Ƽ��[39]����������AM����Ķ����Ժ������ܶ�֮���м���������ع�ϵ,�����о���AM����Ķ������������ܶȼ��������;Ҳ���о�������Ⱦ��������ǿ��AM�������������,���߲�������ǿ��AM�����������Ⱦ�Բ�[40],�����о���AM�����Ⱦ���������ܶ�֮������������,���Ͽ����������������͡�ֲ�������ʩ�ʴ�ʩ�IJ�ͬ,����AM�������̼ˮ�������������ͬ,�Ӷ�AM�������������������Ⱦ�Ͳ�������Ҳ�ͬ[41]��
CCA�������������������������DOC��AHN��NH4+-N��AM����ķֲ���һ����Ӱ��,�����������������������벻��̼Դ�͵�Դ,���˵� C/N ��������Ӱ�������Ⱥ��ṹ[42]��AM�����Ⱥ��ֲ�Ҳ�ܵ�AK��Ӱ��,�طʵ�ʩ�������˲���AM�������������4��ͼ8��,�������о�����������Ч�ض�AM�����Ӱ��ܿ��ܲ���ô��Ҫ,��������AM����Ŀ��ܸ���Ҫ[43,44],����������Ч�ض�AM�����Ӱ���о�Ҳ����,������AM�����Ӱ���д���һ���о���
���о�ʹ��DGGE����������AM�����Ⱥ��ṹ,���˼������������ľ����ԡ���Ȼ���о�ͨ��DGGE�����ɹ�������AM���,������ʵ��ֵ֮����ܻ����ڲ��;����,DGGE����ȫ�����������ȫ������Ⱥ��,ֻ�ܶ�����Ⱥ������������1%��������Ⱥ���з���[45],���о���6���������������AM���������Ϊ22��,��ϵAM���������Ϊ9��,ͨ�����������13��OUT����˲���DGGE�������ܵ��������м���ϵ��Ⱦ��AM��������ԡ���Щ������������PCR�����������븲�ǶȲ������[46],������Ժ���о���Ӧ��AM�������������Ż������������,�����Illumina Miseq������������Pac Bio������������ȷ��������̶ȱ�֤�����ȷ�ԡ�
4 ����
���ڶ�λʩ�������ı�����������������,�Ӷ�Ӱ��AM�����Ⱥ��ṹ,������AM�������������ȾҲ����Ӱ�졣����ʩ��������������AM�����ҪΪ����ù���;�����ù��,����Ⱦ����ϵ��AM���ֻ������ù��,������Ч�ء��������л�̼���̬������������AM�����Ⱥ�仯����Ҫ���ء�����ʩ���ܴٽ�����AM�������������ϵ����Ⱦ,����AM��������ܶ���Ҫ�ܵ����������̬������Ч�ס���Ч�ء��������л�̼��������ˮ����Ӱ��,����Ⱦ����Ҫ�ܵ�������̬����Ӱ�졣�ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
URL [��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 2]
[��������: 2]
[��������: 1]
[��������: 1]
DOI:10.1007/s11104-010-0688-4URL [��������: 1]

Arbuscular mycorrhizal fungi (AMF) are crucial for ecosystem functioning, and thus have potential use for sustainable agriculture. In this study, we investigated the impact of organic and mineral...
[��������: 1]
[��������: 1]
DOI:10.1007/s003740000279URL [��������: 1]
A pot experiment was conducted with soil from a long-term (74-year) fertilization field experiment to compare the effects of organic and mineral fertilizers on mycorrhiza formation in clover, and mycorrhiza-mediated plant P uptake. Five treatments were selected from the field experiment representing different forms and levels of P. Mycorrhizal effects on plant growth and P uptake were estimated by comparing plants grown in untreated soil containing indigenous arbuscular mycorrhizal (AM) fungi, with plants grown in pasteurized soil. Short-term versus residual effects of fertilizer/manure were also measured by comparing treatments with or without fertilizers added at the start of the pot experiment. Mycorrhiza formation was greatest in soil that had received no P for 7465years, followed by soil having received 30 or 6065Mg ha �C1 farmyard manure (FYM), and soil having received 25 or 4465kg P ha �C1 in NPK fertilizers. Plant growth and P uptake were severely reduced in the absence of AM fungi for all mineral fertilizer treatments. In contrast, plants growing in soil that had received FYM grew equally well or better when non-mycorrhizal. Recent additions of NK fertilizer and FYM had no effect on mycorrhiza formation, while additions of NPK led to reduced colonization. It thus seems that moderate quantities of FYM have less adverse effects on AM than equivalent amounts of nutrients in NPK fertilizers, a phenomenon that is most likely due to a temporal difference in P availability and its gradual release that balance plant demand.
[��������: 1]
[��������: 1]
DOI:10.1007/s10457-009-9249-3URL [��������: 1]
A field study was carried out on a six-year-old on-farm field trial during long-rains season (April�CAugust) 2003 to investigate the effect of improved fallow systems and phosphorus application on...
[��������: 2]
[��������: 2]
DOI:10.1007/s12665-010-0741-xURL [��������: 1]
There is considerable discussion and uncertainty in the literature regarding the importance of fresh litter versus older soil organic matter as sources of soil dissolved organic carbon (DOC) in forest floor. In this study, the differences of organic carbon concentration and stable isotope composition were analyzed under different background conditions to identify the origins of DOC in forest soil. The data show that there is no significant difference in SOC content between these collected soil samples ( P 02>020.05), but the litter-rich surface soils have relatively higher DOC concentration than the litter-lacking ( P 02<020.01) ones, and the �� 13 C values of DOC ( �� 13 C DOC ) are closer to �� 13 C of litter than �� 13 C values of SOC ( �� 13 C SOC ). In the litter-lacking surface soil samples, the range of �� 13 C DOC is between �� 13 C SOC and �� 13 C of dominant plant leaves. These results suggest that DOC mainly derive from litter in the litter-rich surface soil with, and the main path of DOC sources may change with surrounding conditions. In addition, �� 13 C SOC and �� 13 C DOC become more positive, and the absolute values of �� ( �� 13 C DOC 026102 �� 13 C SOC ) decrease with depth in the soil profiles, which indicate that the percentage of DOC below 502cm, derived from degradation of humus, may increase with soil depth.
[��������: 1]
[��������: 1]
DOI:10.2307/3760048URL [��������: 1]
The community of endogonaceous mycorrhizal fungi present in plots with two different cropping histories in a western Kentucky soybean field was analyzed by wet-sieving of spores from field soil samples and determining viability of spores of certain species with a vital stain, and by conducting a "Most Probable Number" (MPN) bioassay procedure. The MPN procedure detected 17 species, while wet-sieving of field soils detected only 10 species. Population densities of viable spores of individual species were usually lower than those of total spores, although the differences were not always statistically significant. Information from MPN bioassays on population densities of individual species is more useful than information on population densities of total propagules determined only by analysis of colonization of roots because mycorrhizal effects on plants probably are due to effects of individual species. Wet-sieving of spores from field soils and MPN bioassay of propagules both yield different and useful information, and both may often be effectively employed.
[��������: 1]
[��������: 1]
DOI:10.1007/s00572-002-0214-5URLPMID:12938031 [��������: 1]
Identification of arbuscular mycorrhizal fungi (AMF) on roots is almost impossible with morphological methods and, due to the presence of contaminating fungi, it is also difficult with molecular biological techniques. To allow broad investigation of the population structure of AMF in the field, we have established a new method to selectively amplify the internal transcribed spacer (ITS) region of most AMF with a unique primer set. Based on available sequences of the rDNA , one primer pair specific for AMF and a few other fungal groups was designed and combined in a nested PCR with the already established primer pair ITS5/ITS4. Amplification from contaminating organisms was reduced by an Alu I restriction after the first reaction of the nested PCR. The method was assessed at five different field sites representing different types of habitats. Members of all major groups within the Glomeromycota (except Archaeosporaceae) were detected at the different sites. Gigasporaceae also proved detectable with the method based on cultivated strains.
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.1007/s11104-004-1829-4URL [��������: 1]
Diversity of arbuscular mycorrhizal fungi (AMF) in 27-year long-term NP-fertilization plots under a maize cropping system in Thailand was studied through spore morphological characterization. The plots received 0�C0, 60�C60, 120�C120 and 180�C18002kg N-P 2 O 5 ha 611 year 611 as ammonium sulfate and triple superphosphate. The plots were sampled monthly for one year, the AMF spores were counted and morphotyped, and taxa were identified after morphotyping and monospecific pot culture. Spore number g 611 soil, relative spore abundance and Shannon-Wiener indexes were calculated. Sixteen putative taxa were recorded from the field of which nine sporulated on maize roots in pot culture. The long-term fertilization caused decreases in AMF total spore numbers and variation in species diversity depended on sampling time. Effects of fertilization on spore number and also relative spore abundance varied with species and sampling time. Among the nine species sporulating under maize, only Acaulospora sp.1 showed no change ( P > 0.003 after Bonferroni correction) in spore number with fertilization in the field; and was therefore classified as an AMF species insensitive to fertilization. Spores of Entrophospora schenckii, Glomus mosseae , Glomus sp.1, Glomus geosporum -like and Scutellospora fulgida , though they decreased in absolute numbers in response to fertilization, showed no change ( P > 0.003 after Bonferroni correction) in relative abundance; these species were classified as AMF species slightly sensitive to fertilization. Three unidentified species of Glomus , though they decreased in absolute numbers in response to fertilization, showed decreases ( P < 0.003 after Bonferroni correction) in relative abundance; these species were classified as AMF species highly sensitive to fertilization.
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.1016/j.apsoil.2012.02.004URL [��������: 1]
Plants trigger various responses in the organisms living around them using a wide array of phytochemicals, which are components of their adaptation to a biological environment. The roots of five varieties of chickpea inoculated with Glomus intraradices were extracted, and extracts were fractionated, first based on solubility in methanol and further by HPLC. We found a relationship between chickpea genotype and root phytochemical composition. Several HPLC fractions repressed the germination of AM fungal spores in bioassays conducted in multi-well plates with extracts from the variety CDC Anna. This repression may be an expression of the control of the plant on the AM fungal symbionts. Glomus etunicatum and Gigaspora rosea spore germination responded differently to exposure to the HPLC fractions soluble in 25% methanol. A differential response of AM fungal species to plant phytochemicals could be involved in the so called ��host preference�� of AM fungi. Whereas extensin and other proteins were identified in a bioactive root extract fraction, other proteins undetected by LC�CMS/MS analysis and non-peptidic compounds may be involved in AMF spore germination suppression.
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.1038/nature03608URLPMID:15944706 [��������: 1]
Arbuscular mycorrhizal (AM) fungi form mutualistic, symbiotic associations with the roots of more than 80% of land plants(1). The fungi are incapable of completing their life cycle in the absence of a host root. Their spores can germinate and grow in the absence of a host, but their hyphal growth is very limited. Little is known about the molecular mechanisms that govern signalling and recognition between AM fungi and their host plants. In one of the first stages of host recognition, the hyphae of AM fungi show extensive branching in the vicinity of host roots before formation of the appressorium(2-4), the structure used to penetrate the plant root. Host roots are known to release signalling molecules that trigger hyphal branching(5-7), but these branching factors have not been isolated. Here we have isolated a branching factor from the root exudates of Lotus japonicus and used spectroscopic analysis and chemical synthesis to identify it as a strigolactone, 5-deoxy-strigol. Strigolactones are a group of sesquiterpene lactones, previously isolated as seed-germination stimulants for the parasitic weeds Striga and Orobanche(8). The natural strigolactones 5-deoxy-strigol, sorgolactone and strigol, and a synthetic analogue, GR24, induced extensive hyphal branching in germinating spores of the AM fungus Gigaspora margarita at very low concentrations.
[��������: 1]
[��������: 1]
DOI:10.1007/s00572-002-0206-5URLPMID:12836080 [��������: 1]
This paper explores if Mimosa species (Fabaceae-Mimosoideae) can serve as arbuscular mycorrhizal (AM) and nutrient "resource islands" in six plant communities in the semiarid valley of Tehuacán-Cuicatlán, Mexico. Spatial heterogeneity related to the occurrence of Mimosa species results in temporal differences in AM-fungal spore numbers and soil nutrients. A higher number of AM-fungal spores were found in the soil below the canopies of six endemic Mimosa species than in the soil from non-vegetated areas. For four species, Mimosa adenantheroides , Mimosa calcicola , Mimosa luisana and Mimosa polyantha , the soil below their canopies had more AM-fungal spores than the soil in non-vegetated areas during the wet season than during the dry season. Two species, Mimosa lacerata and Mimosa texana var. filipes , however, had more spores under their canopies during the dry season than during the wet season. Although physical differences are present within and between sites, in general the soil below the canopies of Mimosa species had significantly higher nutrient levels than the soil from non-vegetated areas. Mimosa species thus form "resource islands" that are not only rich in nutrients but also in mycorrhizal propagules. Mimosa species can serve as mycorrhizal "resource islands" by directly affecting AM-fungal spore dynamics and/or by serving as spore-traps. A range of plants associated with Mimosa species may benefit from the higher number of AM propagules. We believe that the use of Mimosa resource islands as an option for biodiversity conservation and for land restoration ought to be considered in the Valley.
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.1007/s00572-005-0014-9URLPMID:16133255 [��������: 1]
Arbuscular mycorrhizal fungal (AMF) spore communities were surveyed in a long-term field fertilization experiment in Switzerland, where different amounts of phosphorus (P) were applied to soil. Plots receiving no P as well as plots systematically fertilized in excess to plant needs for 31 years were used to test the hypothesis that application of P fertilizer changes the composition and diversity of AMF communities. AMF spores were isolated from the field soil, identified, and counted so as to quantify the effect of P fertilization on AMF spore density, composition, and diversity. Trap cultures were established from field soil with four host plants (sunflower, leek, maize, and Crotalaria grahamiana ), and the spore communities were then analyzed in substrate samples from the pots. Altogether, nine AMF species were detected in the soil. No evidence has been acquired for effect of P fertilization on spore density, composition, and diversity of AMF in both the field soil and in trap cultures. On the other hand, we observed strong effect of crop plant species on spore densities in the soil, the values being lowest under rapeseed and highest under Phacelia tanacetifolia covercrop. The identity of plant species in trap pots also significantly affected composition and diversity of associated AMF communities, probably due to preferential establishment of symbiosis between certain plant and AMF species. AMF spore communities under mycorrhizal host plants (wheat and Phacelia in the fields and four host plant species in trap pots) were dominated by a single AMF species, Glomus intraradices . This resulted in exceptionally low AMF spore diversity that seems to be linked to high clay content of the soil.
URL [��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}