 ,2, 张效生3, 张金龙3, 储明星,1
,2, 张效生3, 张金龙3, 储明星,1Tissue Expression and Polymorphism of Sheep SMAD1 Gene and Their Association with Litter Size
TIAN ZhiLong1,2, TANG JiShun1, SUN Qing1, WANG YuQin,2, ZHANG XiaoSheng3, ZHANG JinLong3, CHU MingXing,1通讯作者:
收稿日期:2018-09-3接受日期:2018-12-29网络出版日期:2019-02-16
| 基金资助: |
Received:2018-09-3Accepted:2018-12-29Online:2019-02-16
作者简介 About authors
田志龙,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (1304KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
田志龙, 汤继顺, 孙庆, 王玉琴, 张效生, 张金龙, 储明星. 绵羊SMAD1基因组织表达及其多态性与产羔数关联分析[J]. 中国农业科学, 2019, 52(4): 755-766 doi:10.3864/j.issn.0578-1752.2019.04.015
TIAN ZhiLong, TANG JiShun, SUN Qing, WANG YuQin, ZHANG XiaoSheng, ZHANG JinLong, CHU MingXing.
0 引言
【研究意义】绵羊产羔数是一个极其复杂的性状,受诸多因素影响。大多数绵羊都是季节性发情、单羔品种,且绵羊产羔性状的遗传力较低,仅为0.1左右,常规育种技术在短期内很难改良产羔性状。因此,阐明绵羊高繁殖力形成的分子机理,为分子标记辅助选择提供有效位点,就变得尤为重要。【前人研究进展】在哺乳动物卵泡发育和颗粒细胞生长分化过程中,需要促性腺激素、性腺类固醇激素和细胞因子等共同调控,其中细胞因子以旁分泌和自分泌方式在生殖细胞间进行信号对话,实现对卵泡发育和颗粒细胞生长分化的调控作用,主要包括Wnt/β-catenin[1,2]、MAPK[3]和TGF-β/Smad[4]信号通路等。SMAD蛋白是TGF-β/SMADs信号通路下游重要的转录因子,介导细胞外TGF-β信号在细胞内的转导[5,6]。研究表明,TGF-β/SMADs信号通路在哺乳动物卵巢卵泡的发育,卵泡的闭锁与选择过程中发挥极其重要的调控作用[7]。SMAD1作为TGF-β/SMADs信号通路下游重要的转录因子,广泛分布于不同的组织并介导多种类型的生理过程[8]。绵羊FecB基因是学术界公认影响绵羊产羔数的主效基因,FecB基因属于TGF-β/SMADs信号通路。一个FecB基因拷贝可以增加排卵数1.3—1.6枚,两个FecB基因拷贝可以增加排卵数2.73枚;携带一个FecB基因拷贝的母羊产羔数增加0.9—1.2只,携带两个FecB基因拷贝的母羊产羔数增加1.1—1.7只[9]。【本研究切入点】国内外对TGF-β/SMADs信号通路相关基因的研究较多,但对SMAD1基因在绵羊中的表达及其多态性的研究并不多见。并且在实际生产中发现,在小尾寒羊群体中FecB基因野生型个体存在产多羔的现象。【拟解决的关键问题】本研究拟通过使用Taqman探针法对小尾寒羊进行分型,选择FecB基因突变野生型小尾寒羊,连续观察黄体数并记录产羔数(产羔数和黄体数均大于或等于2,则定义为多羔)。获得小尾寒羊FecB基因突变野生型单羔群体和多羔群体(下文简称单羔、多羔),以此为研究对象,采用反转录PCR结合实时荧光定量PCR技术检测SMAD1基因在不同繁殖力小尾寒羊群体中的表达差异,期望从转录水平初步揭示SMAD1基因在绵羊多羔繁殖中的作用,同时对小尾寒羊、湖羊、苏尼特羊、滩羊、策勒黑羊、草原型藏羊等6个绵羊品种的SMAD1基因g.12485895A>G、g.12487558G>A、g.12508874T>C、g.12487467A>G、g.12487190G>T等5个SNPs进行研究,为绵羊标记辅助选择提供新的参考位点。1 材料与方法
试验于2018年1—8月在中国农业科学院北京畜牧兽医研究所实验室进行。1.1 样品采集和主要试剂
不同繁殖力小尾寒羊(FecB基因野生型单羔和多羔)来自天津市畜牧兽医研究所种羊场,从中挑选健康状况良好成年母羊各3只,使用阴道孕酮栓(controlled internal drug release,CIDR)进行同期发情处理,12d后撤栓。在撤栓后45 h屠宰并立即采集大脑、小脑、下丘脑、垂体、子宫、卵巢、输卵管、心脏、肝脏、脾脏、肺脏、肾脏、肾上腺、甲状腺、大肠、小肠、胰腺、瘤胃、肾脂 19个组织,液氮中保存,直至提取RNA。动物组织总RNA提取试剂盒(DP431)和血液基因组DNA提取系统(DP349)购自天根生化科技有限公司(北京),Taq PCR Master Mix购于北京拓英坊科技有限公司,反转录试剂盒(RR037A)和荧光定量染料(RR820A)均购自大连宝生物公司,基因分型试剂和仪器均来自康普森生物技术有限公司(北京)。
1.2 组织总RNA/DNA的提取和检测
采用动物组织总RNA提取试剂盒/血液基因组DNA提取系统(天根,北京)提取各组织总RNA/DNA,并用Nanodrop 2000检测提取RNA/DNA的纯度和浓度,用1.2%的琼脂糖凝胶电泳检测RNA/DNA完整性。1.3 引物设计
根据GenBank提供的绵羊SMAD1基因mRNA序列(登录号为:XM_012097420.2)利用Primer Premier 6.0软件进行跨外显子引物设计,以甘油醛-3-磷酸脱氢酶(Glyceraldehyde-3-Phosphate Dehydrogenase , GAPDH)(XM_015089125.1)作为内参基因。FecB基因分型引物及探针参考文献[10]。引物由北京天一辉远生物科技有限公司合成,具体信息见表1。Table 1
表1
表1绵羊SMAD1基因的引物信息
Table 1
| 引物及探针名称 Primer names | 引物序列 Primer sequences | 退火温度 Annealing temperature (℃) | 扩增片段(bp) Product size | 用途 Usage |
|---|---|---|---|---|
| SMAD1 | F: TGGTTCCAAGACACAGCGAAT R: GGTGTATCTGCTGGCATCTGA | 60 | 253 | 半定量RT-PCR semi-quantitative RT-PCR |
| SMAD1Q | F: CCCGAGTGGGTGTAGTTT R: TCCTGGCGGTGGTATTCT | 60 | 162 | 荧光定量PCR qPCR |
| GAPDH | F: TGACGCTCCCATGTTTGTGA R: TCATAAGTCCCTCCACGATGC | 60 | 149 | 半定量RT-PCR 和荧光定量PCR semi-quantitative RT-PCR and qPCR |
| FecB-Taqman | F: CCAGCTGGTTCCGAGAGACA R: CTTATACTCACCCAAGATGTTTTCATG | 60 | 73 | FecB-Taqman分型 FecB-Taqman genotyping |
| FecB-Taqman P-G FecB-Taqman P-A | AAATATATCGGACGGTGTT-MGB AAATATATCAGACGGTGTTG-MGB | 60 |
新窗口打开|下载CSV
1.4 半定量RT-PCR反应及荧光定量PCR
试验用cDNA合成及RT-PCR和荧光定量PCR试验过程如周梅[11]所述。简言之,即按反转录试剂盒提供步骤进行cDNA的合成,使用2×Taq PCR Master Mix对SMAD1基因进行PCR,最后使用SYBR?Premix Ex TaqTM Ⅱ进行荧光定量检测。1.5 实时荧光定量检测及数据处理
荧光定量检测利用Roche Light Cycler?480Ⅱ型荧光定量PCR仪进行,以GAPDH为内参基因,每个样品重复检测3次。采用2-ΔΔCT法[12]计算目的基因相对表达量,利用SPSS19.0统计软件对数据进行单因素方差分析,使用邓肯氏(Duncan’s)法进行多重比较。1.6 基因分型及数据处理
使用Taqman分型技术对有产羔记录的380只小尾寒羊进行FecB(A746G)基因型检测,具体方法如刘秋月之前所述[10]。简言之,以小尾寒羊DNA为模板,利用表1中所述引物和探针,进行荧光定量PCR,检测荧光值的变化并对绵羊FecB(A746G)基因型进行判定。采用Sequenom MassARRAY?SNP技术[11,12,13]对SMAD1基因g.12485895A>G、g.12487558G>A、g.12508874T>C、g.12487467A>G、g.12487190G>T位点进行基因型检测。分型样品选择:有产羔数记录的小尾寒羊380只、湖羊101只、策勒黑羊52只、苏尼特羊21只、滩羊22只、草原型藏羊161只。分型样品为DNA,每个样品需要量为20 μL,DNA浓度为40—80 ng·μL-1。绵羊SMAD1基因5个SNPs位点的群体遗传学分析及其与小尾寒羊产羔数关联分析如笔者之前所述[14]。将与小尾寒羊产羔产生显著关联的位点,使用yijklmn=μ+Gi+Gj+Gi×Gj+Pl+Sm+eijklmn 进行多基因聚合分析。其中,yijklmn是性状观察值(产羔数);μ为总体均值;Gi为i标记基因主效应;Gj为j标记基因主效应;Gi×Gj为i和j基因的互作效应;Pl为胎次效应;Sm为季节效应;eijklmn为随机残差效应。用SPSS19.0软件程序中一般线性模型完成小尾寒羊基因型与产羔表型数据关联分析,所有数据以“平均值±标准误”表示。
2 结果
2.1 总RNA提取与cDNA合成
提取不同繁殖力小尾寒羊各组织的总RNA,经1.2%琼脂糖凝胶电泳检测。28S和18S条带清晰可见,且灰度值约为2﹕1,而5S带几乎没有,说明提取的总RNA无明显降解(图1),Nanodrop 2000检测各组织RNA,A260/A280均在1.8—2.1之间,表明提取的RNA纯度较高,可用于后续的荧光定量PCR反应。以反转录之后的cDNA为模板对GAPDH基因进行RT-PCR扩增,结果显示GAPDH基因在小尾寒羊不同组织扩增效果良好,具有明显内参基因特征。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1RNA电泳检测
M代表 DL2000 DNA Marker;1-7分别代表:大脑、小脑、下丘脑、垂体、子宫、卵巢、输卵管
Fig. 1Electrophoresis of the RNA
M:DL2000 DNA Marker;1-7:Brain, Cerebellum, Hypothalamus, Pituitary, Uterus, Ovary, Oviduct, respectively
2.2 SMAD1基因在不同繁殖力小尾寒羊群体各组织中的表达分析
半定量RT-PCR结果显示,SMAD1基因在全身性组织均有表达,且在小尾寒羊单、多羔群体中表达量趋势较为一致(图2)。在小尾寒羊单羔群体下丘脑-垂体-卵巢轴组织的表达量略高于多羔群体。随后使用实时荧光定量对小尾寒羊单、多羔群体5个繁殖相关组织SMAD1基因的表达进行研究,结果表明:SMAD1基因在小尾寒羊下丘脑-垂体-卵巢轴组织均有表达(图3),在小尾寒羊单羔群体卵巢的表达量极显著高于多羔群体(P<0.01),小尾寒羊单羔群体下丘脑、垂体的表达量显著高于多羔群体(P<0.05),在其余组织则不存在显著差异(P>0.05)。图2
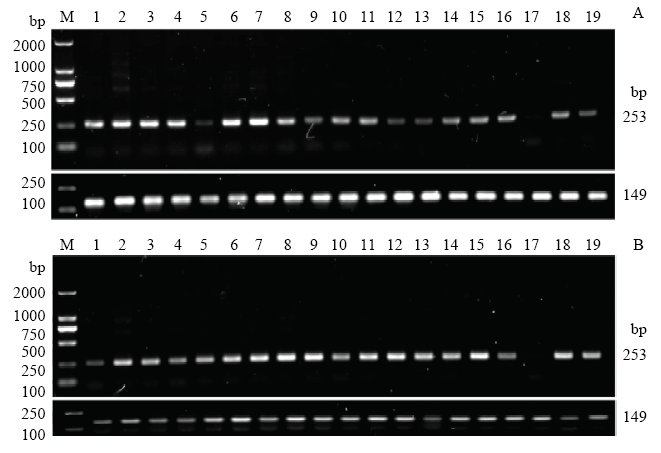 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2SMAD1基因在小尾寒羊不同繁殖力群体19个组织中的半定量RT-PCR
M:DL2000 DNA Marker;1-19:大脑、小脑、下丘脑、垂体、子宫、卵巢、输卵管、心脏、肝脏、脾脏、肺脏、肾脏、肾上腺、甲状腺、大肠、小肠、胰腺、瘤胃、肾脂;253 bp和149 bp分别代表SMAD1和GAPDH基因扩增产物大小;A代表单羔群体、B代表多羔群体
Fig. 2Semi-quantitative RT-PCR of SMAD1 in 19 tissues of Small Tail Han sheep
M:DL2000 DNA Marker;1-19:Brain, Cerebellum, Hypothalamus, Pituitary, Uterus, Ovary, Oviduct, Heart, Liver, Spleen, Lung, Kidney, Adrenal glands, Thyroid, Large intestine, Small intestine, Pancreas, Rumen, Kidney fat; 253 bp and 149 bp indicate the product size of SMAD1 and GAPDH, respectively; A: uniparous sheep B: multiparous sheep
图3
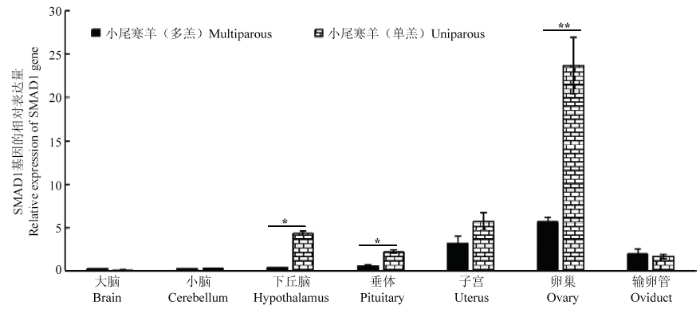 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3SMAD1基因在不同繁殖力小尾寒羊群体各组织间的表达
*表示差异显著(P<0.05);**表示差异极显著(P<0.01)
Fig. 3Expression of SMAD1 gene in 7 tissues in Small Tail Han sheep with different fertility
The * means significant difference (P<0.05); the** means highly significant difference (P<0.01)
2.3 SMAD1基因多态性分析
通过分型发现SMAD1基因5个SNPs在绵羊群体中均存在3种基因型,见图4。由表2可知,除了g.12508874T>C、g.12487467A>G位点外,其余3个SNPs在单、多羔绵羊品种中基因型和等位基因频率均差异显著(P<0.05)。群体遗传学分析发现(表3),SMAD1基因g.12485895A>G、g.12487558G>A、g.12487467A>G、g.12487190G>T位点在小尾寒羊、湖羊、草原型藏羊、策勒黑羊、苏尼特羊、滩羊群体中均为中度多态(0.25<PIC<0.5),g.12508874T>C位点在滩羊中表现为低度多态(PIC<0.25),在其他绵羊群体为中度多态(0.25<PIC<0.5)。g.12485895A>G、g.12487467A>G位点在小尾寒羊和草原型藏羊中表现为Hardy-Weinberg 不平衡状态(P<0.05);g.12487558G>A位点在小尾寒羊和湖羊中表现为Hardy-Weinberg 不平衡状态(P<0.05);g.12487190G>T位点在草原型藏羊中表现为Hardy-Weinberg 不平衡状态(P<0.05),其余SNPs在不同绵羊品种中均表现为Hardy-Weinberg 平衡状态(P>0.05)。图4
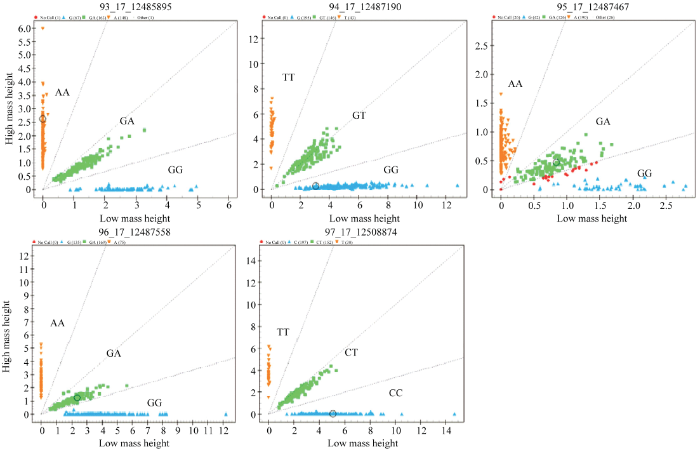 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4SMAD1基因5个SNPs位点分型结果
Fig. 4Genotyping results of five SNPs in SMAD1 gene
Table 2
表2
表2SMAD1基因位点在单、多羔绵羊品种中的基因型频率和等位基因频率
Table 2
| 位点 Locus | 基因型 Genotype | 多羔基因型频率 Genotype frequency in multiparous sheep | 单羔基因型频率 Genotype frequency in uniparous sheep | 卡方检验 χ2 test (P value) | 等位基因 Allele | 多羔中基因频率 Allele frequency in multiparous sheep | 单羔中基因频率 Allele frequency in uniparous sheep | 卡方检验 χ2 test (P value) |
|---|---|---|---|---|---|---|---|---|
| g.12485895A>G | AA | 0.30(167) | 0.52 (104) | 0.00 | A | 0.54 | 0.68 | 0.00 |
| GA | 0.47 (254) | 0.34 (70) | G | 0.46 | 0.32 | |||
| GG | 0.23 (124) | 0.14 (29) | ||||||
| g.12487558G>A | AA | 0.25 (139) | 0.15 (32) | 0.00 | A | 0.48 | 0.36 | 0.00 |
| GA | 0.45 (248) | 0.40 (81) | G | 0.52 | 0.64 | |||
| GG | 0.30(161) | 0.45 (91) | ||||||
| g.12508874T>C | CC | 0.56 (307) | 0.58 (118) | 0.69 | C | 0.74 | 0.76 | 0.50 |
| CT | 0.37 (203) | 0.37 (75) | T | 0.26 | 0.24 | |||
| TT | 0.07 (39) | 0.05(11) | ||||||
| g.12487467A>G | AA | 0.51 (249) | 0.59 (111) | 0.15 | A | 0.69 | 0.73 | 0.11 |
| GA | 0.35 (169) | 0.28 (52) | G | 0.31 | 0.27 | |||
| GG | 0.14 (68) | 0.13 (24) | ||||||
| g.12487190G>T | GG | 0.44 (250) | 0.56 (115) | 0.01 | G | 0.66 | 0.72 | 0.01 |
| GT | 0.43 (239) | 0.32 (65) | T | 0.34 | 0.28 | |||
| TT | 0.13 (73) | 0.12(24) |
新窗口打开|下载CSV
Table 3
表3
表3SMAD1基因不同位点在不同绵羊品种中的群体遗传学分析
Table 3
| 位点 Locus | 品种 Breed | 基因型频率 Genotype frequency | 基因频率 Allele frequency | 多态信息含量(PIC) Polymorphism information content | 杂合度 (HE) Heterozygosity (HE) | 有效等位 基因数 Effective number of allele (NE) | 卡方值(P) χ2 test (P value) | |||
|---|---|---|---|---|---|---|---|---|---|---|
| g.12485895A>G | AA | AG | GG | A | G | |||||
| 小尾寒羊 Small Tail Han sheep | 0.33 | 0.44 | 0.23 | 0.55 | 0.45 | 0.37 | 0.49 | 1.98 | 0.02 | |
| 湖羊 Hu sheep | 0.35 | 0.54 | 0.11 | 0.62 | 0.38 | 0.36 | 0.47 | 1.89 | 0.12 | |
| 草原型藏羊 Prairie Tibetan sheep | 0.58 | 0.32 | 0.10 | 0.73 | 0.27 | 0.31 | 0.39 | 1.64 | 0.02 | |
| 策勒黑羊 Cele black sheep | 0.06 | 0.50 | 0.44 | 0.31 | 0.69 | 0.34 | 0.43 | 1.74 | 0.21 | |
| 苏尼特羊 Sunite sheep | 0.29 | 0.48 | 0.23 | 0.52 | 0.48 | 0.37 | 0.50 | 2.00 | 0.83 | |
| 滩羊 Tan sheep | 0.27 | 0.41 | 0.32 | 0.48 | 0.52 | 0.37 | 0.50 | 2.00 | 0.40 | |
| g.12487558G>A | AA | GA | GG | A | G | |||||
| 小尾寒羊 Small Tail Han sheep | 0.25 | 0.43 | 0.32 | 0.47 | 0.53 | 0.37 | 0.50 | 1.99 | 0.01 | |
| 湖羊 Hu sheep | 0.18 | 0.34 | 0.48 | 0.36 | 0.64 | 0.35 | 0.46 | 1.85 | 0.00 | |
| 草原型藏羊 Prairie Tibetan sheep | 0.10 | 0.40 | 0.50 | 0.30 | 0.70 | 0.33 | 0.42 | 1.73 | 0.60 | |
| 策勒黑羊 Cele black sheep | 0.35 | 0.52 | 0.13 | 0.61 | 0.39 | 0.36 | 0.48 | 1.91 | 0.53 | |
| 位点 Locus | 品种 Breed | 基因型频率 Genotype frequency | 基因频率 Allele frequency | 多态信息含量(PIC) Polymorphism information content | 杂合度 (HE) Heterozygosity (HE) | 有效等位 基因数 Effective number of allele (NE) | 卡方值(P) χ2 test (P value) | |||
| 苏尼特羊 Sunite sheep | 0.33 | 0.29 | 0.38 | 0.48 | 0.52 | 0.37 | 0.50 | 2.00 | 0.05 | |
| 滩羊 Tan sheep | 0.41 | 0.45 | 0.14 | 0.64 | 0.36 | 0.36 | 0.46 | 1.86 | 0.93 | |
| g.12508874T>C | CC | CT | TT | C | T | |||||
| 小尾寒羊 Small Tail Han sheep | 0.61 | 0.34 | 0.05 | 0.78 | 0.22 | 0.29 | 0.35 | 1.53 | 0.64 | |
| 湖羊 Hu sheep | 0.34 | 0.50 | 0.16 | 0.59 | 0.41 | 0.37 | 0.48 | 1.94 | 0.67 | |
| 草原型藏羊 Prairie Tibetan sheep | 0.56 | 0.38 | 0.06 | 0.75 | 0.25 | 0.31 | 0.38 | 1.60 | 0.94 | |
| 策勒黑羊 Cele black sheep | 0.58 | 0.38 | 0.04 | 0.77 | 0.23 | 0.29 | 0.36 | 1.55 | 0.55 | |
| 苏尼特羊 Sunite sheep | 0.57 | 0.38 | 0.05 | 0.76 | 0.24 | 0.30 | 0.36 | 1.57 | 0.82 | |
| 滩羊 Tan sheep | 0.73 | 0.27 | 0.00 | 0.86 | 0.14 | 0.21 | 0.24 | 1.31 | 0.46 | |
| g.12487467A>G | AA | GA | GG | A | G | |||||
| 小尾寒羊 Small Tail Han sheep | 0.54 | 0.30 | 0.16 | 0.69 | 0.31 | 0.34 | 0.43 | 1.75 | 0.00 | |
| 湖羊 Hu sheep | 0.62 | 0.33 | 0.05 | 0.78 | 0.22 | 0.28 | 0.34 | 1.52 | 0.84 | |
| 草原型藏羊 Prairie Tibetan sheep | 0.65 | 0.25 | 0.10 | 0.78 | 0.22 | 0.28 | 0.34 | 1.52 | 0.00 | |
| 策勒黑羊 Cele black sheep | 0.22 | 0.55 | 0.23 | 0.49 | 0.51 | 0.37 | 0.50 | 2.00 | 0.46 | |
| 苏尼特羊Sunite sheep | 0.43 | 0.38 | 0.19 | 0.62 | 0.38 | 0.36 | 0.47 | 1.88 | 0.42 | |
| 滩羊 Tan sheep | 0.29 | 0.38 | 0.33 | 0.48 | 0.52 | 0.37 | 0.50 | 2.00 | 0.28 | |
| g.12487190G>T | GG | GT | TT | G | T | |||||
| 小尾寒羊 Small Tail Han sheep | 0.45 | 0.41 | 0.14 | 0.65 | 0.35 | 0.35 | 0.45 | 1.83 | 0.08 | |
| 湖羊 Hu sheep | 0.61 | 0.34 | 0.05 | 0.78 | 0.22 | 0.28 | 0.34 | 1.52 | 0.90 | |
| 草原型藏羊 Prairie Tibetan sheep | 0.62 | 0.29 | 0.09 | 0.77 | 0.23 | 0.29 | 0.35 | 1.55 | 0.00 | |
| 策勒黑羊 Cele black sheep | 0.19 | 0.58 | 0.23 | 0.48 | 0.52 | 0.37 | 0.50 | 2.00 | 0.26 | |
| 苏尼特羊 Sunite sheep | 0.38 | 0.48 | 0.14 | 0.62 | 0.38 | 0.36 | 0.47 | 1.89 | 0.96 | |
| 滩羊 Tan sheep | 0.27 | 0.41 | 0.32 | 0.48 | 0.52 | 0.37 | 0.50 | 2.00 | 0.40 | |
新窗口打开|下载CSV
2.4 SMAD1基因多态性与小尾寒羊产羔数的关系
由表4可知,SMAD1基因g.12487190G>T位点多态性与小尾寒羊前三胎产羔数均显著相关(P<0.05),TT基因型母羊产羔数显著高于GT和GG基因型母羊(P<0.05)。其他4个位点与小尾寒羊产羔数之间无显著关联。将该位点与小尾寒羊FecB(A746G)不同基因型进行组合,结果发现,共存在9种不同组合基因型(表5),组合基因型与小尾寒羊第一胎、第二胎、第三胎产羔数均显著关联(P<0.05),其中AA-GG、AA-GT、AA-TT型母羊产羔数显著低于其他组合基因型母羊(P<0.05)。Table 4
表4
表4SMAD1基因SNPs位点各基因型小尾寒羊产羔数关联分析
Table 4
| 位点 Locus | 基因型 Genotype | 第一胎样本数 Number of the first parity | 第一胎产羔数 Litter size of the first parity | 第二胎样本数 Number of the second parity | 第二胎产羔数 Litter size of the second parity | 第三胎样本数 Number of the third parity | 第三胎产羔数 Litter size of the third parity |
|---|---|---|---|---|---|---|---|
| g.12485895A>G | AA | 114 | 2.11±0.08 | 109 | 2.33±0.09 | 45 | 2.80±0.15 |
| GG | 80 | 2.14±0.09 | 74 | 2.24±0.11 | 27 | 2.93±0.20 | |
| GA | 144 | 2.12±0.07 | 139 | 2.37±0.08 | 55 | 2.70±0.14 | |
| g.12487190G>T | GG | 157 | 2.08±0.07a | 150 | 2.28±0.07a | 57 | 2.67±0.14a |
| GT | 144 | 2.16±0.07a | 138 | 2.34±0.08a | 60 | 2.72±0.13a | |
| TT | 51 | 2.39±0.12b | 46 | 2.65±0.13b | 16 | 3.75±0.25b | |
| g.12487467A>G | AA | 159 | 2.11±0.06 | 152 | 2.29±0.07 | 56 | 2.70±0.13 |
| GG | 48 | 2.35±0.12 | 43 | 2.47±0.14 | 15 | 2.80±0.24 | |
| GA | 67 | 2.17±0.09 | 85 | 2.31±0.10 | 36 | 2.67±0.16 | |
| g.12487558G>A | AA | 92 | 2.28±0.09 | 87 | 2.43±0.10 | 35 | 2.91±0.17 |
| GG | 107 | 2.06±0.08 | 99 | 2.31±0.09 | 44 | 2.82±0.15 | |
| GA | 142 | 2.16±0.07 | 138 | 2.38±0.08 | 50 | 2.64±0.14 | |
| g.12508874T>C | CC | 204 | 2.10±0.06 | 193 | 2.40±0.07 | 66 | 2.82±0.13 |
| CT | 120 | 2.27±0.08 | 113 | 2.35±0.09 | 54 | 2.82±0.14 | |
| TT | 18 | 2.11±0.02 | 19 | 2.21±0.21 | 9 | 2.56±0.35 |
新窗口打开|下载CSV
Table 5
表5
表5SMAD1与FecB组合基因型与小尾寒羊产羔数关联分析
Table 5
| 基因型 Genotype | 第一胎样本数 Number of the first parity | 第一胎产羔数 Litter size of the first parity | 第二胎样本数 Number of the second parity | 第二胎产羔数 Litter size of the second parity | 第三胎样本数 Number of the third parity | 第三胎产羔数 Litter size of the third parity |
|---|---|---|---|---|---|---|
| AA-GG | 23 | 1.00±0.16a | 23 | 1.00±0.18a | 23 | 1.04±0.19a |
| AA-GT | 14 | 1.36±0.21a | 14 | 1.36±0.22a | 14 | 1.07±0.24a |
| AA-TT | 10 | 1.10±0.25a | 10 | 1.30±0.27a | 10 | 1.10±0.29a |
| AG-GG | 77 | 2.04±0.09b | 75 | 2.40±0.10b | 27 | 2.67±0.18b |
| AG-GT | 70 | 2.07±0.09b | 67 | 2.16±0.10b | 34 | 2.65±0.16b |
| AG-TT | 27 | 2.14±0.15b | 24 | 2.50±0.17b | 10 | 3.40±0.29b |
| GG-GG | 57 | 2.33±0.21b | 53 | 2.38±0.12b | 22 | 3.00±0.19b |
| GG-GT | 60 | 2.40±0.10b | 57 | 2.67±0.11b | 18 | 3.22±0.21b |
| GG-TT | 14 | 2.29±0.10b | 13 | 2.31±0.23b | 6 | 3.33±0.37b |
新窗口打开|下载CSV
3 讨论
3.1 SMAD1基因表达与动物繁殖的关系
研究表明,TGF-β/SMADs信号通路参与了胚胎早期发生、骨形成和组织修复等过程,在细胞的增殖、分化、迁移和凋亡以及免疫和内分泌等方面发挥着重要的调节作用[15,16]。近来的研究发现,TGF-β/SMADs信号通路在哺乳动物的卵泡发育、卵泡的选择与闭锁过程中发挥极其重要的调控作用[17]。在这一过程中,该通路中的关键成员(配体、受体、信号分子等)在整个生殖系统中都有协调的时空表达模式、控制和调节卵巢的生理学功能[18]。此外,研究发现,有丝分裂原活化蛋白激酶(Ras-MAPK)信号通路以及骨形态发生蛋白(BMPs)通过改变类固醇合成酶基因表达来调节颗粒细胞类固醇的合成[3]。大量研究表明,颗粒细胞的发育与卵泡的发育成熟和排卵密切相关[19],而颗粒细胞的发育过程又受到多种激素和激素受体的调节,这其中雌二醇、孕酮、卵泡刺激素(follicle stimulating hormone,FSH)和促黄体素(luteinizing hormone, LH)通过与其它细胞因子互作影响颗粒细胞增殖、分化以及卵泡成熟[20,21]。体外大鼠颗粒细胞的培养试验表明BMP4和BMP7可以提高FSH依赖的E2分泌[22],而BMP4、BMP6、BMP7和BMP15抑制FSH诱导的P4的分泌[23,24,25]。其他研究也表明,阻断BMP/SMADs信号通路抑制了类固醇合成酶基因CYP19A1的表达[17]。据报道SMAD6作为BMP/SMAD信号通路抑制剂,可以通过SMAD1/5/8通路显著影响BMP15,GDF9和BMPRIB的功能表达,并导致绵羊排卵数增加[26]。因此,推测SMAD1通过调节BMP15,GDF9和BMPRIB功能来控制绵羊排卵数进而控制产羔数。目前,对SMAD1基因的研究多集中于小鼠,对小尾寒羊的研究相对较少。本研究中,RT-PCR和实时荧光定量PCR结果显示,SMAD1基因在全身性组织均有表达,且在小尾寒羊单、多羔群体中表达量趋势较为一致。这和牛家强[27,28]等研究结果一致,同样也与所有TGF-β超家族很多成员都需要通过SMAD1蛋白进行信号转导的研究结论一致。SMAD1基因在小尾寒羊单羔群体卵巢的表达量显著高于多羔群体(P<0.01),小尾寒羊单羔群体下丘脑、垂体的表达量显著高于多羔群体(P<0.05)。下丘脑-垂体-卵巢轴是控制哺乳动物性激素分泌的重要系统,与动物繁殖密切相关。SMAD1基因在此系统的差异表达,意味着该基因可能通过下丘脑-垂体-卵巢轴的负反馈途径调节卵泡的发育和排卵,进而影响不同繁殖力小尾寒羊群体的繁殖性状。但由于并未进行SMAD1蛋白的定量研究,该结果仍需继续研究。3.2 SMAD1基因多态性与动物繁殖的关系
现阶段关于SMAD1基因多态与动物繁殖的报道多集中于水生动物,在哺乳动物中的研究较少。池秋蝶等[29]通过外显子测序获得文蛤SMAD1基因多态信息,并发现其多态性可以影响文蛤的生长发育。本研究发现SMAD1基因g.12487190G>T位点在小尾寒羊、湖羊、草原型藏羊、策勒黑羊、苏尼特羊、滩羊群体中均为中度多态(0.25<PIC<0.5),并且其多态性与小尾寒羊前三胎产羔数显著相关(P<0.05)。对该位点进行基因定位研究发现,该位点处于SMAD1基因第5内含子区域。虽然内含子不编码氨基酸,无法影响蛋白质结构,但其突变可能控制基因转录活性并影响SMAD1基因的表达。关于内含子变异影响动物表型的报道很多。秦川牛SMAD3基因内含子3(g.101664C>G)和内含子5(g.114523A>G)变异显著影响其体重、胸围和臀部长度[30](P<0.05)。皖江白鹅SMAD9基因内含子2的C>T变异,显著影响其产蛋性能[31]。陕北白绒山羊[32]赖氨酸脱甲基酶6A (lysine demethylase 6A, KDM6A)基因第17内含子的16个碱基的插入缺失导致其mRNA表达量的不同,最终显著提高产羔数(P<0.05)。这些研究表明,内含子变异不仅影响转录和剪接过程,还可以通过控制基因表达来调控动物表型。XU等[33]研究发现位于SMAD1基因内含子区域的rs40635766突变与湖羊繁殖力密切相关,可能是控制湖羊多羔繁殖的一个主效基因,这和本研究的结果一致。g.12485895A>G、g.12487467A>G、g.12487558G>A、g.12487190G>T位点在绵羊群体中存在不同程度的Hardy-Weinberg不平衡状态。可能是由于参与分型计算的样本量不够多或者绵羊群体存在人工选育造成的此类现象。本研究发现g.12487190G>T位点TT基因型小尾寒羊母羊前三胎的产羔数显著高于GT和GG基因型母羊(P<0.05)。将该位点与小尾寒羊FecB(A746G)不同基因型进行组合,发现AA-GG、AA-GT、AA-TT型母羊产羔数显著低于其他组合基因型母羊(P<0.05),而AA-GG、AA-GT、AA-TT型母羊产羔数不存在差异(P>0.05)。该结果暗示,SMAD1基因调控绵羊产羔性状的机制与FecB(A746G)基因密不可分。4 结论
本研究发现,SMAD1基因在绵羊组织中呈全身性表达,在小尾寒羊单羔群体下丘脑、垂体、卵巢的表达量高于多羔群体(P<0.05)。g.12487190G>T突变中TT基因型母羊产羔数显著高于GT和GG基因型母羊(P<0.05)。因此,SMAD1基因可能是影响小尾寒羊产羔数的一个重要候选基因,且g.12487190G>T位点对小尾寒羊多羔性状选育具有一定的指导意义。(责任编辑 林鉴非)
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1530/REP-14-0685URLPMID:26130815 [本文引用: 1]

Abstract Wingless-type mouse mammary tumor virus integration site (WNT) signaling molecules are locally secreted glycoproteins that play important role in regulation of ovarian follicle maturation and steroid production. Components of the WNT signaling pathway have been demonstrated to impact reproductive functions, including embryonic development of the sex organs and regulation of follicle maturation controlling steroidogenesis in the postnatal ovary. Emerging evidence underscores the complexity of WNT signaling molecules in regulation of dynamic changes that occur in the ovary during the reproductive cycle. While disruption in the WNT signaling cascade has been recognized to have deleterious consequences to normal sexual development, more recent studies are beginning to highlight the importance of these molecules in adult ovarian function related to follicle development, corpus luteum formation, steroid production and fertility. Hormonal regulation of WNT genes and expression of members of the WNT signaling network, including WNT ligands, frizzled receptors, and downstream signaling components that are expressed in the postnatal ovary at distinct stages of the estrous cycle suggest a crucial role in normal ovarian function. Similarly, FSH stimulation of T-cell factor-dependent gene expression requires input from 0205-catenin, a lynchpin molecule in canonical WNT signaling, further indicating 0205-catenin participation in regulation of follicle maturation. This review will focus on the multiple functions of WNT signaling in folliculogenesis in the adult ovary. 0008 2015 Society for Reproduction and Fertility.
DOI:10.1016/j.mce.2013.11.007URLPMID:24246780 [本文引用: 1]
Wnt signaling is an evolutionarily conserved pathway that regulates cell proliferation, differentiation and apoptosis. To investigate the possible role of Wnt signaling in the regulation of ovarian follicular development, secondary follicles were isolated and cultured in vitro in the presence or absence of its activator (LiCl or Wnt3a) or inhibitor (IWR-1). We have demonstrated that activation of 尾-catenin signals by activators dramatically suppressed follicular development by increasing granulosa cell apoptosis and inhibiting follicle steroidogenesis. In contrast, inhibition of Wnt signaling by IWR-1 was observed with better developed follicles and increased steroidogenesis. Further studies have shown that the transcription factor Forkhead box O3a (Foxo3a) and its downstream target molecules were modulated by the activators or the inhibitor. These findings provide evidence that Wnt signaling might negatively regulate follicular development potentially through Foxo3a signaling components.
DOI:10.1126/science.1171396URLPMID:2847890 [本文引用: 2]
A surge of luteinizing hormone (LH) from the pituitary gland triggers ovulation, oocyte maturation, and luteinization for successful reproduction in mammals. Because the signaling molecules RAS and ERK1/2 (extracellular signal-regulated kinases 1 and 2) are activated by an LH surge in granulosa cells of preovulatory follicles, we disrupted Erk1/2 in mouse granulosa cells and provide in vivo evidence that these kinases are necessary for LH-induced oocyte resumption of meiosis, ovulation, and luteinization. In addition, biochemical analyses and selected disruption of the Cebpb gene in granulosa cells demonstrate that C/EBPβ (CCAAT/Enhancer-binding protein-β) is a critical downstream mediator of ERK1/2 activation. Thus, ERK1/2 and C/EBPβ constitute an in vivo LH-regulated signaling pathway that controls ovulation- and luteinization-related events.
DOI:10.1093/humupd/dmw039URLPMID:5155571 [本文引用: 1]
Initially identified for their capability to induce heterotopic bone formation, bone morphogenetic proteins (BMPs) are multifunctional growth factors that belong to the transforming growth factor 尾 superfamily. Using cellular and molecular genetic approaches, recent studies have implicated intra-ovarian BMPs as potent regulators of ovarian follicular function. The bi-directional communication of oocytes and the surrounding somatic cells is mandatory for normal follicle development and oocyte maturation. This review summarizes the current knowledge on the physiological role and molecular determinants of these ovarian regulatory factors within the human germline-somatic regulatory loop. The regulation of ovarian function remains poorly characterized in humans because, while the fundamental process of follicular development and oocyte maturation is highly similar across species, most information on the regulation of ovarian function is obtained from studies using rodent models. Thus, this review focuses on the studies that used human biological materials to gain knowledge about human ovarian biology and disorders and to develop strategies for preventing, diagnosing and treating these abnormalities. Relevant English-language publications describing the roles of BMPs or growth differentiation factors (GDFs) in human ovarian biology and phenotypes were comprehensively searched using PubMed and the Google Scholar database. The publications included those published since the initial identification of BMPs in the mammalian ovary in 1999 through July 2016. Studies using human biological materials have revealed the expression of BMPs, GDFs and their putative receptors as well as their molecular signaling in the fundamental cells (oocyte, cumulus/granulosa cells (GCs) and theca/stroma cells) of the ovarian follicles throughout follicle development. With the availability of recombinant human BMPs/GDFs and the development of immortalized human cell lines, functional studies have demonstrated the physiological role of intra-ovarian BMPs/GDFs in all aspects of ovarian functions, from follicle development to steroidogenesis, cell ell communication, oocyte maturation, ovulation and luteal function. Furthermore, there is crosstalk between these potent ovarian regulators and the endocrine signaling system. Dysregulation or naturally occurring mutations within the BMP system may lead to several female reproductive diseases. The latest development of recombinant BMPs, synthetic BMP inhibitors, gene therapy and tools for BMP-ligand sequestration has made the BMP pathway a potential therapeutic target in certain human fertility disorders; however, further clinical trials are needed. Recent studies have indicated that GDF8 is an intra-ovarian factor that may play a novel role in regulating ovarian functions in the human ovary. Intra-ovarian BMPs/GDFs are critical regulators of folliculogenesis and human ovarian functions. Any dysregulation or variations in these ligands or their receptors may affect the related intracellular signaling and influence ovarian functions, which accounts for several reproductive pathologies and infertility. Understanding the normal and pathological roles of intra-ovarian BMPs/GDFs, especially as related to GC functions and follicular fluid levels, will inform innovative approaches to fertility regulation and improve the diagnosis and treatment of ovarian disorders.
DOI:10.1111/j.1440-1681.2011.05663.xURLPMID:22211842 [本文引用: 1]
Diabetic nephropathy (DN) is a major diabetic complication that is mediated by transforming growth factor (TGF)-02051 via Smad-dependent and -independent signalling pathways.Under diabetic conditions, many profibrotic factors, such as advanced glycation end-products and angiotensin II, can also activate the Smad signalling pathway via the extracellular signal-regulated kinase/p38 mitogen-activated protein kinase090009Smad signalling cross-talk pathway. Thus, Smads act as signal integrators and interact with other signalling pathways to mediate DN.In the context of renal fibrosis, Smad3 is pathogenic, but Smad2 is protective. Deletion of Smad3 inhibits, whereas disruption of Smad2 upregulates, connective tissue growth factor and vascular endothelial growth factor expression and promotes both epithelial090009myofibroblast and endothelial090009myofibroblast transition.Smad7 plays a protective role in DN because deletion of Smad7 enhances, whereas overexpression of Smad7 inhibits, Smad3-mediated renal fibrosis and nuclear factor-0202B-driven renal inflammation.Transforming growth factor-02051 activates Smad3 to regulate microRNAs that mediate renal fibrosis. Of these, miR-21 and miR-192 are upregulated, whereas the miR-29 and miR-200 families are downregulated.Targeting downstream TGF-0205/Smad signalling by overexpressing Smad7- or Smad3-dependent microRNA related to fibrosis may represent a novel and effective strategy for the treatment of DN.
DOI:10.1038/nrc3500URLPMID:23612460 [本文引用: 1]
Much of the focus on the transforming growth factor-β (TGFβ) superfamily in cancer has revolved around the TGFβ ligands themselves. However, it is now becoming apparent that deregulated signalling by many of the other superfamily members also has crucial roles in both the development of tumours and metastasis. Furthermore, these signalling pathways are emerging as plausible therapeutic targets. Their roles in tumorigenesis frequently reflect their function in embryonic development or in adult tissue homeostasis, and their influence extends beyond the tumours themselves, to the tumour microenvironment and more widely to complications of cancer such as cachexia and bone loss.
DOI:10.1111/j.1440-1797.2005.00334.xURLPMID:15705182 [本文引用: 1]
SUMMARY: Extensive studies have demonstrated that transforming growth factor-beta (TGF-0205) plays an important role in the progression of renal diseases. TGF-0205 exerts its biological functions mainly through its downstream signalling molecules, Smad2 and Smad3. It is now clear that Smad3 is critical for TGF-0205's pro-fibrotic effect, whereas the functions of Smad2 in fibrosis in response to TGF-0205 still need to be determined. Our recent studies have demonstrated that Smad signalling is also a critical pathway for renal fibrosis induced by other pro-fibrotic factors, such as angiotensin II and advanced glycation end products (AGE). These pro-fibrotic factors can activate Smads directly and independently of TGF-0205. They can also cause renal fibrosis via the ERK/p38 MAP kinase090009Smad signalling cross-talk pathway. In contrast, blockade of Smad2/3 activation by overexpression of an inhibitory Smad7 prevents collagen matrix production induced by TGF-0205, angiotensin II, high glucose and AGE and attenuates renal fibrosis in various animal models including rat obstructive kidney, remnant kidney and diabetic kidney diseases. Results from these studies indicate that Smad signalling is a key and final common pathway of renal fibrosis. In addition, TGF-0205 has anti-inflammatory and immune-regulatory properties. Our most recent studies demonstrated that TGF-0205 transgenic mice are protected against renal inflammation in mouse obstructive and diabetic models. Upregulation of renal Smad7, thereby blocking NF.0202B activation via induction of I0202B02±, is a central mechanism by which TGF-0205 inhibits renal inflammation. In conclusion, TGF-0205 signals through Smad2/3 to mediate renal fibrosis, whereas induction of Smad7 inhibits renal fibrosis and inflammation. Thus, targeting Smad signalling by overexpression of Smad7 may have great therapeutic potential for kidney diseases.
DOI:10.1093/emboj/19.8.1745URL [本文引用: 1]
Oxford University Press
DOI:10.1016/j.anireprosci.2005.06.001URLPMID:15982834 [本文引用: 1]
Twenty-one of the world's prolific sheep breeds and strains were tested for the presence of the FecB mutation of BMPR1B and the FecX I mutation of BMP15. The breeds studied were Romanov (2 strains), Finn (2 strains), East Friesian, Teeswater, Blueface Leicester, Hu, Han, D an, Chios, Mountain Sheep (three breeds), German Whiteheaded Mutton, Lleyn, Loa, Galician, Barbados Blackbelly (pure and crossbred) and St. Croix. The FecB mutation was found in two breeds, Hu and Han from China, but not in any of the other breeds. The 12 Hu sheep sampled were all homozygous carriers of FecB ( FecB B /FecB B) whereas the sample of 12 Han sheep included all three genotypes ( FecB B /FecB B, FecB B/ FecB +, FecB +/ FecB +) at frequencies of 0.33, 0.58 and 0.08, respectively. There was no evidence of FecX I in any of the breeds sampled.
DOI:10.11843/j.issn.0366-6964.2017.01.005URLMagsci [本文引用: 2]
<p>基于前期分型基础,为了更进一步提高分型的效率和准确性,本试验建立了Taqman探针和SNaPshot两种高通量<em>FecB</em>突变分型方法。结果显示,与传统的PCR-RFLP和PCR-SSCP分型方法相比,这两种高通量分型方法在节省检测成本的同时,大大缩短了检测周期,并在检测效率上也有了大幅度的提高,对利用<em>FecB</em>进行分子育种具有潜在的应用价值。自2003年至今,本实验室应用这两种方法对10个中国本地绵羊品种、3个培育品种以及一些杂交群体的23 264只绵羊的<em>FecB</em>频率进行了检测,结果显示,湖羊和小尾寒羊携带<em>FecB</em>基因的频率最高,小尾寒羊<em>FecB</em>基因的B等位基因频率为 0.432~0.833。跟踪以小尾寒羊作为母本和杜泊羊级进杂交后代群体发现,随着杂交的进行<em>FecB</em>基因在培育的鲁西黑头肉羊中的B等位基因频率提升到了0.432,但在横交固定阶段有所下降。本研究所得结果可为今后在肉用绵羊品种多羔改良和多羔肉用新品种培育过程中通过引入<em>FecB</em>基因提高繁殖性能提供参考数据。</p>
DOI:10.11843/j.issn.0366-6964.2017.01.005URLMagsci [本文引用: 2]
<p>基于前期分型基础,为了更进一步提高分型的效率和准确性,本试验建立了Taqman探针和SNaPshot两种高通量<em>FecB</em>突变分型方法。结果显示,与传统的PCR-RFLP和PCR-SSCP分型方法相比,这两种高通量分型方法在节省检测成本的同时,大大缩短了检测周期,并在检测效率上也有了大幅度的提高,对利用<em>FecB</em>进行分子育种具有潜在的应用价值。自2003年至今,本实验室应用这两种方法对10个中国本地绵羊品种、3个培育品种以及一些杂交群体的23 264只绵羊的<em>FecB</em>频率进行了检测,结果显示,湖羊和小尾寒羊携带<em>FecB</em>基因的频率最高,小尾寒羊<em>FecB</em>基因的B等位基因频率为 0.432~0.833。跟踪以小尾寒羊作为母本和杜泊羊级进杂交后代群体发现,随着杂交的进行<em>FecB</em>基因在培育的鲁西黑头肉羊中的B等位基因频率提升到了0.432,但在横交固定阶段有所下降。本研究所得结果可为今后在肉用绵羊品种多羔改良和多羔肉用新品种培育过程中通过引入<em>FecB</em>基因提高繁殖性能提供参考数据。</p>
DOI:10.11843/j.issn.0366-6964.2018.03.008URL [本文引用: 2]
旨在探究绵羊FGF 7基因组织表达水平及其多态性与产羔数之间的关系.本研究利用半定量反转录聚合酶链式反应(sqRT-PCR)和实时荧光定量PCR(qPCR)技术对绵羊FGF 7基因在多羔小尾寒羊和单羔苏尼特羊以及小尾寒羊FecB不同基因型(BB、B+、++)各组织中的表达水平进行研究,同时采用Sequenom MassARRAY(R) SNP技术,对多羔绵羊品种(小尾寒羊380只)和单羔绵羊品种(滩羊、苏尼特羊、萨福克羊、杜泊羊和草原型藏羊共380只)FGF 7基因g.57842893C>T位点多态性进行检测,并与小尾寒羊产羔数进行关联分析.结果表明,FGF 7基因在单、多羔绵羊的心和肺中高表达,在其它各组织呈中等或低丰度表达;FGF 7基因在多羔小尾寒羊绝大部分组织中的表达量均高于单羔苏尼特羊,但差异均不显著(P>0.05),FGF 7在小尾寒羊FecB不同基因型中的表达并无明显差异.g.57842893C>T位点共存在CC、CT和TT 3种基因型,且基因型频率和等位基因频率在单、多羔品种间差异均达到显著水平(P≤0.05);g.57842893C>T位点在苏尼特羊、萨福克羊和杜泊羊中表现为中度多态(0.25<PIC<0.50),在其它品种中表现为低度多态(PIC<0.25);g.57842893C>T位点仅在苏尼特羊、萨福克羊以及杜泊羊3个品种中均处于哈代温伯格平衡状态(P>0.05);g.57842893C>T位点与小尾寒羊第1、2以及第3胎产羔数均无显著关联(P>0.05),但CC型各胎产羔数均高于TT型.综上表明,FGF 7基因的表达水平与绵羊产羔数可能存在一定程度的正相关,虽然可能不是影响绵羊产羔数的关键基因,但g.57842893C>T位点对绵羊产羔数性状的选育具有一定的指导意义.
DOI:10.11843/j.issn.0366-6964.2018.03.008URL [本文引用: 2]
旨在探究绵羊FGF 7基因组织表达水平及其多态性与产羔数之间的关系.本研究利用半定量反转录聚合酶链式反应(sqRT-PCR)和实时荧光定量PCR(qPCR)技术对绵羊FGF 7基因在多羔小尾寒羊和单羔苏尼特羊以及小尾寒羊FecB不同基因型(BB、B+、++)各组织中的表达水平进行研究,同时采用Sequenom MassARRAY(R) SNP技术,对多羔绵羊品种(小尾寒羊380只)和单羔绵羊品种(滩羊、苏尼特羊、萨福克羊、杜泊羊和草原型藏羊共380只)FGF 7基因g.57842893C>T位点多态性进行检测,并与小尾寒羊产羔数进行关联分析.结果表明,FGF 7基因在单、多羔绵羊的心和肺中高表达,在其它各组织呈中等或低丰度表达;FGF 7基因在多羔小尾寒羊绝大部分组织中的表达量均高于单羔苏尼特羊,但差异均不显著(P>0.05),FGF 7在小尾寒羊FecB不同基因型中的表达并无明显差异.g.57842893C>T位点共存在CC、CT和TT 3种基因型,且基因型频率和等位基因频率在单、多羔品种间差异均达到显著水平(P≤0.05);g.57842893C>T位点在苏尼特羊、萨福克羊和杜泊羊中表现为中度多态(0.25<PIC<0.50),在其它品种中表现为低度多态(PIC<0.25);g.57842893C>T位点仅在苏尼特羊、萨福克羊以及杜泊羊3个品种中均处于哈代温伯格平衡状态(P>0.05);g.57842893C>T位点与小尾寒羊第1、2以及第3胎产羔数均无显著关联(P>0.05),但CC型各胎产羔数均高于TT型.综上表明,FGF 7基因的表达水平与绵羊产羔数可能存在一定程度的正相关,虽然可能不是影响绵羊产羔数的关键基因,但g.57842893C>T位点对绵羊产羔数性状的选育具有一定的指导意义.
DOI:10.1038/nprot.2008.73URL [本文引用: 2]
DOI:10.1111/age.12420URLPMID:26923315 [本文引用: 1]
Summary We evaluated 69 SNPs in genes previously related to fertility and production traits for their relationship to daughter pregnancy rate (DPR), cow conception rate (CCR) and heifer conception rate (HCR) in a separate population of Holstein cows grouped according to their predicted transmitting ability (PTA) [≤611 ( n = 1287) and ≥1.5 ( n = 1036)] for DPR. Genotyping was performed using Sequenom MassARRAY03. There were a total of 39 SNPs associated with the three fertility traits. The SNPs that explained the greater proportion of the genetic variation for DPR were COQ9 (3.2%), EPAS1 (1.0%), CAST (1.0%), C7H19orf60 (1.0%) and MRPL48 (1.0%); for CCR were GOLGA4 (2.4%), COQ9 (1.8%), EPAS1 (1.1%) and MRPL48 (0.8%); and for HCR were HSD17B7 (1.0%), AP3B1 (0.8%), HSD17B12 (0.7%) and CACNA1D (0.6%). Inclusion of 39 SNPs previously associated with DPR in the genetic evaluation system increased the reliability of PTA for DPR by 0.20%. Many of the genes represented by SNPs associated with fertility are involved in steroidogenesis or are regulated by steroids. A large proportion of SNPs previously associated with genetic merit for fertility in Holstein bulls maintained their association in a separate population of cows. The inclusion of these genes in genetic evaluation can improve reliabilities of genomic estimates for fertility.
URL [本文引用: 1]
URL [本文引用: 1]
DOI:10.1074/jbc.M112.380493URLPMID:22942286 [本文引用: 1]
Abstract SARA has been shown to be a regulator of epithelial cell phenotype, with reduced expression during TGF-β1-mediated epithelial-to-mesenchymal transition. Examination of the pathways that might play a role in regulating SARA expression identified phosphatidylinositol 3-kinase (PI3K) pathway inhibition as sufficient to reduce SARA expression. The mechanism of PI3K inhibition-mediated SARA down-regulation differs from that induced by TGF-β1 in that, unlike TGF-β1, PI3K-dependent depletion of SARA was apparent within 6 h and did not occur at the mRNA or promoter level but was blocked by inhibition of proteasome-mediated degradation. This effect was independent of Akt activity because neither reducing nor enhancing Akt activity modulated the expression of SARA. Therefore, this is likely a direct effect of p85α action, and co-immunoprecipitation of SARA and p85α confirmed that these proteins interact. Both SARA and PI3K have been shown to be associated with endosomes, and either LY294002 or p85α knockdown enlarged SARA-containing endocytic vesicles. Inhibition of clathrin-mediated endocytosis blocked SARA down-regulation, and a localization-deficient mutant SARA was protected against down-regulation. As inhibiting PI3K can activate the endosomal fusion-regulatory small GTPase Rab5, we expressed GTPase-deficient Rab5 and observed endosomal enlargement and reduced SARA protein expression, similar to that seen with PI3K inhibition. Importantly, either interference with PI3K via LY294002 or p85α knockdown, or constitutive activity of the Rab5 pathway, enhanced the expression of smooth muscle α-actin. Together, these data suggest that although TGF-β1 can induce epithelial-to-mesenchymal transition through reduction in SARA expression, SARA is also basally regulated by its interaction with PI3K.
DOI:10.1002/stem.1208URLPMID:22911900 [本文引用: 1]
Upon secretion, transforming growth factor β (TGFβ) is maintained in a sequestered state in extracellular matrix as a latent form. The latent TGFβ is considered as a molecular sensor that releases active TGFβ in response to the perturbations of the extracellular matrix at the situations of mechanical stress, wound repair, tissue injury, and inflammation. The biological implication of the temporal discontinuity of TGFβ storage in the matrix and its activation is obscure. Here, using several animal models in which latent TGFβ is activated in vascular matrix in response to injury of arteries, we show that active TGFβ controls the mobilization and recruitment of mesenchymal stem cells (MSCs) to participate in tissue repair and remodeling. MSCs were mobilized into the peripheral blood in response to vascular injury and recruited to the injured sites where they gave rise to both endothelial cells for re-endothelialization and myofibroblastic cells to form thick neointima. TGFβs were activated in the vascular matrix in both rat and mouse models of mechanical injury of arteries. Importantly, the active TGFβ released from the injured vessels is essential to induce the migration of MSCs, and cascade expression of monocyte chemotactic protein-1 stimulated by TGFβ amplifies the signal for migration. Moreover, sustained high levels of active TGFβ were observed in peripheral blood, and at the same time points following injury, Sca1+CD29+CD11b61CD4561 MSCs, in which 91% are nestin+ cells, were mobilized to peripheral blood and recruited to the remodeling arteries. Intravenously injection of recombinant active TGFβ1 in uninjured mice rapidly mobilized MSCs into circulation. Furthermore, inhibitor of TGFβ type I receptor blocked the mobilization and recruitment of MSCs to the injured arteries. Thus, TGFβ is an injury-activated messenger essential for the mobilization and recruitment of MSCs to participate in tissue repair/remodeling. STEM CELLS2012;30:2498–2511
DOI:10.1631/jzus.B1000079URLPMID:2932882 [本文引用: 2]
Bone morphogenetic proteins (BMPs) play a critical role in the growth and steroidogenesis of granulosa cells (GCs). BMP signals act through membrane-bound heteromeric serine/threonine kinase receptors. Upon ligand binding, BMPs activate intracellular Smad proteins and regulate growth and apoptosis in various cell types. The objective of this study was to demonstrate the effects of BMP/Smad signal on growth and steroidogenesis of porcine GCs.A strategy of RNA interference (RNAi)-mediated 'gene silencing' of Smad4, a core molecule mediating the intracellular BMP/Smad signal transduction pathways, was used to interrupt endogenous BMP/Smad signaling. Results indicate that Smad4-small interfering RNA (siRNA) caused specific inhibition of Smad4 mRNA and protein expression after transfection. Interrupted endogenous BMP/Smad signaling significantly inhibited growth, and induced apoptosis of porcine GCs, while decreasing estradiol production. In addition, interrupted BMP/Smad signaling significantly (P 0.05)changed the expression of Cyclin D2, CDK4, Bcl-2, and Cyp19a1. These findings provide new insights into how BMP/Smad signaling regulates the growth and steroidogenesis of porcine GCs.
[本文引用: 1]
DOI:10.1016/j.theriogenology.2014.05.020URLPMID:24985562 [本文引用: 1]
61Two steroid hormones were used for in02vitro growth culture of bovine oocytes.61Androstenedione and 17β-estradiol supported in02vitro growth of oocytes for 1402days.61Oocytes grew from around 9502μm to full size (12002μm) in this culture medium.61More than half of the cultured oocytes acquired the maturational competence to MII.61Transzonal projections were maintained during culture with steroid hormones.
DOI:10.5713/ajas.15.0310URLPMID:4782083 [本文引用: 1]
The aim of the present study was to examine the effects of testosterone (T) and estradiol-() on the production of progesterone () by granulosa cells, and of the on the production of and T by theca internal cells. In the first experiment, granulosa cells isolated from the largest () and third largest () preovulatory follicle were incubated for 4 h in short-term culture system, production by granulosa cells of both and was increased in a dose-dependent manner by ovine luteinizing hormone (oLH), but not T or . In the second experiment, and granulosa cells cultured for 48 h in the developed monolayer culture system were recultured for an additional 48 h with increasing doses of various physiological active substances existing in the ovary, including T and . Basal production for 48 h during 48 to 96 h of the cultured was about nine fold greater by granulosa cells than by granulosa cells. In substances examined oLH, chicken vasoactive intestinal polypeptide (cVIP) and T, but not , stimulated in a dose-dependent manner production in both and granulosa cells. In addition, when the time course of production by granulosa cells in response to oLH, cVIP, T and was examined for 48 h during 48 to 96 h of culture, although had no effect on production by granulosa cells of during the period from 48 to 96 h of culture, production with oLH was found to be increased at 4 h of the culture, with a maximal 9.14 fold level at 6 h. By contrast, production with cVIP and T increased significantly (p<0.05) from 8 and 12 h of the culture, respectively, with maximal 6.50 fold response at 12 h and 6, 48 fold responses at 36 h. Furthermore, when granulosa cells were precultured with for various times before 4 h culture with oLH at 96 h of culture, the increase in production in response to oLH with a dose-related manner was only found at a pretreatment time of more than 12 h. In the third experiment, theca internal cells of , and the largest third to fifth preovulatory follicles () were incubated for 4 h in short-term culture system with increasing doses of . The production of and T by theca internal cells were increased with the addition of of . These increases were greater in smaller follicles. These results indicate that, in granulosa cells of the hen, T may have a direct stimulatory action in the long term on production, and on in long-term action which may enhance the sensitivity to LH for production, and thus, in theca internal cells, in short term action may stimulate the production of and T.
DOI:10.1530/REP-14-0581URLPMID:25948249 [本文引用: 1]
The aim of the present study was to determine the direct cause of the mutation-induced, increased rate in Booroola Merino (BB) . Granulosa were removed from antral follicles before and post-from BB (n=5) and WT (n=12) Merino ewes. Direct immunofluorescence measurement of mature receptors using flow cytometry demonstrated a significant up-regulation of FSH receptor (), transforming beta type 1, bone receptor (), and LH receptor () in BB . The increased density of and provide novel evidence of a mechanism for increasing the number of follicles that are recruited during dominant follicle selection. The compounding increase in receptors with increasing follicle size maintained the multiple follicles and reduced the apoptosis, which contributed to a high rate in BB . In addition, we report a mutation-independent mechanism of down-regulation to reduce receptor density of the leading dominant follicle in . The suppression of receptor density coincides with the cessation of mitogenic and steroidogenic differentiation as part of the of the follicle. The BB mutation-induced attenuation of led to an increased density of the and and a concurrent reduction in apoptosis to increase the rate. The role of in receptor modulation is implicated in the development of multiple .
DOI:10.1073/pnas.96.13.7282URLPMID:10377406 [本文引用: 1]
Bone morphogenetic proteins (BMPs) comprise a large group of polypeptides in the transforming growth factor superfamily with essential physiological functions in morphogenesis and organogenesis in both vertebrates and invertebrates. At present, the role of BMPs in the reproductive system of any species is poorly understood. Here, we have established the existence of a functional BMP system in the ovary, replete with ligand, receptor, and novel cellular functions. In situ hybridization histochemistry identified strong mRNA labeling for BMP-4 and -7 in the theca cells and BMP receptor types IA, IB, and II in the granulosa cells and oocytes of most follicles in ovaries of normal cycling rats. To explore the paracrine function of this BMP system, we examined the effects of recombinant BMP-4 and -7 on FSH (follicle-stimulating hormone)-induced rat granulosa cytodifferentiation in serum-free medium. Both BMP-4 and -7 regulated FSH action in positive and negative ways. Specifically, physiological concentrations of the BMPs enhanced and attenuated the stimulatory action of FSH on estradiol and progesterone production, respectively. These effects were dose- and time-dependent. Furthermore, the BMPs increased granulosa cell sensitivity to FSH. Thus, BMPs have now been identified as molecules that differentially regulate FSH-dependent estradiol and progesterone production in a way that reflects steroidogenesis during the normal estrous cycle. As such, it can be hypothesized that BMPs might be the long-sought "luteinization inhibitor" in Graafian follicles during their growth and development.
DOI:10.1095/biolreprod65.4.994URL [本文引用: 1]
DOI:10.1677/joe.0.1690001URLPMID:11250641 [本文引用: 1]
Abstract Responsiveness of the fetal sheep adrenal gland to adrenocorticotrophin (ACTH) increases in late pregnancy, resulting in increased glucocorticoid production. Development of this responsiveness is an important determinant of fetal hypothalamic-pituitary-adrenal function and depends, in part, on the potential for ACTH binding to adrenal tissue. In the present study, we have examined the developmental pattern of ACTH receptor (ACTH-R) expression during the latter half of pregnancy and in neonatal and adult life. As hypoxaemia induces increases in cortisol and ACTH secretion, in addition to increasing fetal adrenal responsiveness, a further aim of this study was to investigate whether hypoxaemia was associated with altered expression of the ACTH-R gene. Whole adrenal glands were removed from fetal sheep, lambs and adult sheep at different stages of development for measurement of ACTH-R mRNA. Moderate hypoxaemia was induced for 48 h beginning on days 124-128, or on days 132-134 of gestation, by decreasing the maternal fractional inspired oxygen. ACTH-R mRNA was detected by northern blotting using a cDNA cloned in our laboratory and by in situ hybridisation. ACTH-R mRNA (3.6 kb major transcript) was detected in adrenal tissue at day 63 of gestation. Its relative abundance increased significantly (P<0.05) between days 126-128 and 140-141 of pregnancy, increased further with the onset of spontaneous labour, and remained increased in newborn lambs at 7 h-7 days after birth. ACTH-R mRNA levels then decreased in adrenal tissue from lambs and adult sheep (P<0.05). Hypoxaemia for 48 h significantly increased ACTH-R mRNA expression in adrenals of the older fetuses (days 134-136) compared with that in controls (P<0.05), but was without effect in younger fetuses. We conclude that levels of ACTH-R mRNA in the fetal adrenal gland increase as term approaches, coincident with the endogenous prepartum surge in plasma ACTH and cortisol. Sustained hypoxaemia resulted in an upregulation of mRNA encoding for ACTH-R, but only in older fetuses and in association with a sustained increase in plasma cortisol. These results are consistent with cortisol, ACTH, or both, contributing to increased fetal adrenal responsiveness, by increasing expression of fetal adrenal receptors for ACTH.
DOI:10.1074/jbc.M007428200URL [本文引用: 1]
DOI:10.1093/biolre/iox133URLPMID:29088317 [本文引用: 1]
react-text: 115 Using microarrays, researchers are able to obtain a genome wide snapshot of a biological system under a given experimental context. Fortunately, a significant amount of gene regulation data is publicly available through various databases. We present a system that uses extra knowledge in published gene regulation relationships to examine findings in a microarray experiment and to aid biologists... /react-text react-text: 116 /react-text [Show full abstract]
URL [本文引用: 1]
为探索牦牛Smad1基因与bta-miR-1434-5p是否存在靶向关系,探究bta-miR-1434-5p分子调控可能机制,利用RT-PCR技术对bta-miR-1434-5p和Smad1 mRNA在牦牛组织中的表达谱进行检测,并构建pmiR-RBREPORTTM双荧光素酶报告载体对牦牛Smad1基因3′端非翻译区(3′UTR)与bta-miR-1434-5p的靶向关系进行研究。结果表明:Smad1基因mRNA在牦牛下丘脑、垂体、心、肝、脾、肺、肾、骨骼肌、淋巴结、卵巢、输卵管、子宫组织中均有广泛表达;bta-miR-1434-5p除在牦牛输卵管中没有表达外,在其他组织均与Smad1基因mRNA共表达;获得牦牛Smad1基因3′UTR序列并成功构建野生型及突变型pmiR-RB-REPORTTM双荧光素酶报告质粒;荧光素酶活性检测发现bta-miR-1434-5p mimics对Smad1野生型质粒报告荧光没有明显的下调作用,由此推测btamiR-1434-5p与牦牛Smad1基因的该段3′UTR之间未发现明显互作。
URL [本文引用: 1]
为探索牦牛Smad1基因与bta-miR-1434-5p是否存在靶向关系,探究bta-miR-1434-5p分子调控可能机制,利用RT-PCR技术对bta-miR-1434-5p和Smad1 mRNA在牦牛组织中的表达谱进行检测,并构建pmiR-RBREPORTTM双荧光素酶报告载体对牦牛Smad1基因3′端非翻译区(3′UTR)与bta-miR-1434-5p的靶向关系进行研究。结果表明:Smad1基因mRNA在牦牛下丘脑、垂体、心、肝、脾、肺、肾、骨骼肌、淋巴结、卵巢、输卵管、子宫组织中均有广泛表达;bta-miR-1434-5p除在牦牛输卵管中没有表达外,在其他组织均与Smad1基因mRNA共表达;获得牦牛Smad1基因3′UTR序列并成功构建野生型及突变型pmiR-RB-REPORTTM双荧光素酶报告质粒;荧光素酶活性检测发现bta-miR-1434-5p mimics对Smad1野生型质粒报告荧光没有明显的下调作用,由此推测btamiR-1434-5p与牦牛Smad1基因的该段3′UTR之间未发现明显互作。
[D].
[本文引用: 1]
[D].
[本文引用: 1]
DOI:10.7541/2018.035URL [本文引用: 1]
为探讨Smad1/5基因在贝类生长发育中的调控作用,利用RACE技术克隆获得文蛤Smad1/5(Mm-Smad1/5)基因的cDNA全长序列,对其生物信息学、不同组织和不同发育时期时空表达特征进行分析,并利用直接测序法分析了外显子区域SNP位点与生长性状的相关性。结果表明:Mm-Smad1/5的cDNA全长序列为1832bp,开放阅读框1380bp,编码459个氨基酸;氨基酸多序列比对显示,Mm-Smad1/5蛋白与太平洋牡蛎Smad5、大西洋舟螺Smad1的一致性分别为83.7%和80.2%,与人、鸡、非洲爪蟾等脊椎动物Smad1、Smad5氨基酸序列的一致性达到70.5%以上,说明该基因具有较高的保守性;结构域预测发现,Mm-Smad1/5含有Smads蛋白家族特有MH1、MH2两个高度保守结构域。荧光定量PCR(qRT-PCR)结果表明,Mm-Smad1/5基因在成体6个组织均有表达,尤其在斧足、外套膜中表达量显著高于其他组织(P<0.05);Mm-Smad1/5基因在各发育时期广泛表达,从原肠胚期开始大量表达,一直持续到壳顶幼虫期,而从眼点幼虫大量下降,稚贝时期又有所上升。Mm-Smad1/5基因外显子区域SNP位点相关分析表明,共发现了9个SNP位点,其中936G>T位点与文蛤的生长性状显著相关(P<0.05)。Mm-Smad1/5基因在文蛤生长发育中发挥重要调控作用,可作为高产良种选育的候选基因,而生长关联SNP位点分析将为文蛤分子标记辅助育种研究奠定重要基础。
DOI:10.7541/2018.035URL [本文引用: 1]
为探讨Smad1/5基因在贝类生长发育中的调控作用,利用RACE技术克隆获得文蛤Smad1/5(Mm-Smad1/5)基因的cDNA全长序列,对其生物信息学、不同组织和不同发育时期时空表达特征进行分析,并利用直接测序法分析了外显子区域SNP位点与生长性状的相关性。结果表明:Mm-Smad1/5的cDNA全长序列为1832bp,开放阅读框1380bp,编码459个氨基酸;氨基酸多序列比对显示,Mm-Smad1/5蛋白与太平洋牡蛎Smad5、大西洋舟螺Smad1的一致性分别为83.7%和80.2%,与人、鸡、非洲爪蟾等脊椎动物Smad1、Smad5氨基酸序列的一致性达到70.5%以上,说明该基因具有较高的保守性;结构域预测发现,Mm-Smad1/5含有Smads蛋白家族特有MH1、MH2两个高度保守结构域。荧光定量PCR(qRT-PCR)结果表明,Mm-Smad1/5基因在成体6个组织均有表达,尤其在斧足、外套膜中表达量显著高于其他组织(P<0.05);Mm-Smad1/5基因在各发育时期广泛表达,从原肠胚期开始大量表达,一直持续到壳顶幼虫期,而从眼点幼虫大量下降,稚贝时期又有所上升。Mm-Smad1/5基因外显子区域SNP位点相关分析表明,共发现了9个SNP位点,其中936G>T位点与文蛤的生长性状显著相关(P<0.05)。Mm-Smad1/5基因在文蛤生长发育中发挥重要调控作用,可作为高产良种选育的候选基因,而生长关联SNP位点分析将为文蛤分子标记辅助育种研究奠定重要基础。
DOI:10.17221/34/2015-CJASURL [本文引用: 1]
SMAD3, a member of SMAD transcription factors, plays a key role in transforming growth factor-beta (TGF-尾) signalling pathway and regulation of muscle growth. However, there was no strong evidence of association between SMAD3 polymorphisms and body traits in animals. In this study, single nucleotide polymorphisms (SNPs) in SMAD3 gene were detected in four Chinese cattle breeds (Qinchuan, Jiaxian, Nanyang, and Caoyuan) by using DNA pool sequencing and PCR-RFLP, and their effects on gene expression and growth traits were evaluated in Qinchuan cattle. The results showed that four novel SNPs (NC_007308.5 c.-2017A>G, g.101664C>G, g.105829A>G, and g.114523A>G) in promoter, intron 3, and intron 5 were found in four cattle breeds. NC_007308.5 c.-2017A>G and g.114523A>G were significantly ( P
DOI:10.1007/s11033-014-3772-7URLPMID:25280542 [本文引用: 1]
Elevation of egg performance is vital to goose farming. Many poultry scientists are seeking for efficient molecular genetic markers associated with egg yield. In this study, mRNA differential display was adopted to investigate gene expression profiling in the follicular development of goose. For the first time, a novel SMAD family protein SMAD9 (EST CJ111007) was found to be involved in follicular initiation and used to be a candidate gene. Functional regions analysis of Smad9 indicated that SMAD9 protein is highly conserved in MH1 and MH2 domains, and the connection area is highly variable region. 6 pairs of primers (p1-p6) were designed to detect the SNPs of Smad9 by PCR-SSCP method. The results revealed that polymorphisms were discovered in the PCR products amplified with P1 primers in exon1 and P3 primers in intron2. In Smad9 exon1, 5 genotypes were found: FK, FF, JJ, JK and KK, including 2 SNPs: 243 bp G → A, 309 bp T → G, the mutations did not result in amino acid mutations; In intron2, 3 genotypes were found: AA, BB and AB, only 1 SNP (C → T). The annual egg yield of FK genotype geese or allele gene A in intron2 are significantly more than those of other genotypes on the average (p < 0.05). Taken together, it is suggested that Smad9 is a promising candidate gene affecting egg performance in goose.
DOI:10.3389/fgene.2018.00091URL [本文引用: 1]
A previous whole-genome association analysis identifiedlysine demethylase 6A(KDM6A), which encodes a type of histone demethylase, as a candidate gene associated to goat fecundity.KDM6Agene knockout mouse disrupts gametophyte development, suggesting that it has a critical role in reproduction. In this study, goatKDM6AmRNA expression profiles were determined, insertion/deletion (indel) variants in the gene identified, indel variants effect onKDM6Agene expression assessed, and their association with first-born litter size analyzed in 2326 healthy female Shaanbei white cashmere goats.KDM6AmRNA was expressed in all tissues tested (heart, liver, spleen, lung, kidney, muscle, brain, skin and testis); the expression levels in testes at different developmental stages [1-week-old (wk), 2, 3 wk, 1-month-old (mo), 1.5 and 2 mo] indicated a potential association with the mitosis-to-meiosis transition, implying thatKDM6Amay have an essential role in goat fertility. Meanwhile, two novel intronic indels of 16 bp and 5 bp were identified. Statistical analysis revealed that only the 16 bp indel was associated with first-born litter size (P< 0.01), and the average first-born litter size of individuals with an insertion/insertion genotype higher than that of those with the deletion/deletion genotype (P< 0.05). There was also a significant difference in genotype distributions of the 16 bp indel between mothers of single-lamb and multi-lamb litters in the studied goat population (P= 0.001). Consistently, the 16 bp indel also had a significant effect onKDM6Agene expression. Additionally, there was no significant linkage disequilibrium (LD) between these two indel loci, consistent with the association analysis results. Together, these findings suggest that the 16 bp indel inKDM6Amay be useful for marker-assisted selection (MAS) of goats.
DOI:10.3389/fgene.2018.00118URL [本文引用: 1]
Reproduction is an important trait in sheep breeding as well as in other livestock. However, despite its importance the genetic mechanisms of litter size in domestic sheep (Ovis aries) are still poorly understood. To explore genetic mechanisms underlying the variation in litter size, we conducted multiple independent genome-wide association studies in five sheep breeds of high prolificacy (Wadi, Hu, Icelandic, Finnsheep, and Romanov) and one low prolificacy (Texel) using the Ovine Infinium HD BeadChip, respectively. We identified different sets of candidate genes associated with litter size in different breeds:BMPR1B, FBN1, andMMP2in Wadi;GRIA2, SMAD1, andCTNNB1in Hu;NCOA1in Icelandic;INHBB, NF1, FLT1, PTGS2, andPLCB3in Finnsheep;ESR2in Romanov andESR1, GHR, ETS1, MMP15, FLI1, andSPP1in Texel. Further annotation of genes and bioinformatics analyses revealed that different biological pathways could be involved in the variation in litter size of females: hormone secretion (FSH and LH) in Wadi and Hu, placenta and embryonic lethality in Icelandic, folliculogenesis and LH signaling in Finnsheep, ovulation and preovulatory follicle maturation in Romanov, and estrogen and follicular growth in Texel. Taken together, our results provide new insights into the genetic mechanisms underlying the prolificacy trait in sheep and other mammals, suggesting targets for selection where the aim is to increase prolificacy in breeding projects.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}