 ,1, 邢蕾蕾1, 曹晓红1, 郭光艳1, 柴建芳,2,*, 秘彩莉,1,*
,1, 邢蕾蕾1, 曹晓红1, 郭光艳1, 柴建芳,2,*, 秘彩莉,1,*Cloning of Ta4CL1 and its function in promoting plant growth and lignin deposition in transgenic Arabidopsis plants
MENG Ying,1, XING Lei-Lei1, CAO Xiao-Hong1, GUO Guang-Yan1, CHAI Jian-Fang,2,*, BEI Cai-Li,1,*通讯作者: *秘彩莉, E-mail:beicaili@sina.com, Tel: 0311-80787574;柴建芳, E-mail:chai87652130@163.com
收稿日期:2020-12-23接受日期:2021-04-14网络出版日期:2021-06-02
| 基金资助: |
Corresponding authors: *E-mail:beicaili@sina.com, Tel: 0311-80787574;E-mail:chai87652130@163.com
Received:2020-12-23Accepted:2021-04-14Published online:2021-06-02
| Fund supported: |
作者简介 About authors
E-mail:mengying941027@tom.com

摘要
4-香豆酸辅酶A连接酶(4CL; EC 6.2.1.12)位于苯丙烷途径分支点的上游, 是苯丙烷代谢途径的核心酶, 可产生木质素、黄酮类等化合物, 这些化合物对植物的生长发育及环境适应性均具有重要作用。在双子叶植物中, 有关4CL的研究较多, 而在单子叶植物尤其是作物中的研究相对较少。本研究利用RACE技术从普通小麦中克隆了一个4CL基因Ta4CL1。系统发育分析表明, Ta4CL1与水稻、玉米和高粱等植物中在木质素合成中具有重要作用的4CLs聚成一类; 利用Ta4CL1过表达、拟南芥4CLs突变体at4cl1、at4cl3和at4cl14cl3及其功能回复株系进行的分析表明, Ta4CL1与At4CL1功能相似, 在植物木质素合成中具有重要作用, 但未参与黄酮类化合物生物合成的调控过程; Ta4CL1是转基因拟南芥中4CL酶活性的主要贡献者。过表达Ta4CL1的转基因拟南芥叶片增大、茎更粗; Ta4CL1的表达还受茉莉酸甲酯(Methyl jasmonic acid, MeJA)、赤霉素(Gibberellin, GA)和生长素(Indoleacetic acid, IAA)等激素处理的影响。本研究为利用基因工程将Ta4CL1应用于改善小麦秸秆的利用效率提供了理论依据。
关键词:
Abstract
4-Coumarate:coenzyme A ligase (4CL; EC 6.2.1.12) acts upstream of the branch point of phenylpropanoid, which is a key enzyme in the phenylpropanoid metabolic pathways for monolignol and flavonoid biosynthesis, and these compounds play important roles in plant growth and development as well as stress adaptability. Many 4CLs had been extensively studied in dicotyledons, but their function in monocotyledons, especially in crops, was relatively poorly understood. In this study, Ta4CL1, which encoded 4-Coumarate:coenzyme A ligase, was isolated from common wheat by RACE technique. Polygenetic analysis revealed that Ta4CL1 could be clustered to the same group of 4CLs from rice, maize, and sorghum, which was mainly involved in lignin biosynthesis. Ta4CL1-overexpressed Arabidopsis lines, at4cl1, at4cl3, and at4cl14cl3 as well as their corresponding functional recovery lines were used to elucidate the function of Ta4CL1 in the phenylpropanoid metabolic pathway. The results suggested that Ta4CL1 had similar function with At4CL1 in regulating lignin biosynthesis but it had no effect on flavonoid biosynthesis. Ta4CL was the major contributor of 4CL enzyme activity in transgenic Arabidopsis plants. Overexpression of Ta4CL led to enlarged leaves and thickened stems in transgenic Arabidopsis seedlings, the expression of Ta4CL was also affected by MeJA, GA, and IAA treatments. These results provide the theoretical basis for improving the utilizing efficiency of plant straws using Ta4CL1 by genetic engineering.
Keywords:
PDF (6678KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
孟颖, 邢蕾蕾, 曹晓红, 郭光艳, 柴建芳, 秘彩莉. 小麦Ta4CL1基因的克隆及其在促进转基因拟南芥生长和木质素沉积中的功能. 作物学报, 2022, 48(1): 63-75 DOI:10.3724/SP.J.1006.2022.01100
MENG Ying, XING Lei-Lei, CAO Xiao-Hong, GUO Guang-Yan, CHAI Jian-Fang, BEI Cai-Li.
木质素是一种芳香族杂聚物, 主要沉积于植物的细胞壁中, 在细胞壁中的含量仅次于纤维素, 可为细胞壁提供机械支撑、防止水分散失以及保护植物组织免受病原菌的侵染[1]。木质素常与细胞壁中的纤维素、半纤维素共价结合形成具有复杂网状结构的木质纤维, 且木质素难以通过自然或生物途径降解; 在利用木质纤维进行制浆的过程中, 须使用强酸或强碱去除木质素。因此植物细胞壁中的木质素不仅影响饲草的营养价值、还影响木材的制浆效率, 并引发环境污染[2]。故此, 有关植物木质素合成途径及其调控机制的研究备受关注。
植物苯丙烷代谢途径(phenylpropanoid metabolic pathway)由莽草酸途径衍生而来, 通过该途径可以产生木质素、黄酮类以及芥子酰基苹果酸类化合物。4-香豆酸辅酶A连接酶(4-Coumarate:coenzyme A ligase, 4CL)位于苯丙烷途径分支点的上游, 可催化将肉桂酸、4-香豆酸、咖啡酸、阿魏酸和芥子酸等转化为相应的CoA硫酯[3], 后者可作为许多次级代谢产物生物合成的底物, 如类黄酮、异黄酮、木质素、木栓质、香豆素以及结合于细胞壁的酚类物质, 因而4CL对苯丙烷途径具有重要的调控作用[4,5,6]。
植物中的4CL常以基因家族的形式存在, 如在拟南芥基因组中有4个4CL基因(4CL1~4CL4)[6], 水稻、玉米和毛白杨的基因组中各有5个4CL基因[7,8,9]。系统发育分析表明, 双子叶植物中的4CL可分为两大类, I类和II类, I类4CL主要参与木质素的生物合成, 而II类4CL往往在苯丙烷途径中木质素以外的途径中起作用[10]。例如, 在拟南芥中, At4CL1和At4CL2主要参与木质素的合成, 而At4CL3主要影响类黄酮的合成[6]。与拟南芥中的情况相似, 在胡杨(Populus tremuloides)中, Pt4CL1在木质部中表达较高, 主要参与木质素的生物合成; Pt4CL2在叶和茎的表皮中表达, 参与黄酮类化合物等酚类物质的生物合成[10]。与双子叶植物相比, 单子叶植物的木质素单体组成不同, 其细胞壁结合的酚类化合物也不尽相同[11]。在水稻中的研究表明, 与双子叶植物中的4CL相比, 单子叶中的4CL具有不同的酶学特性; 抑制Os4CL3的表达不仅降低了转基因水稻中木质素含量, 还影响了水稻的生长发育过程, 如植株矮化和育性降低等[12]; 在玉米、高粱和谷子中均曾发现因自发变异产生的棕色中脉(brown midrib, bm/ bmr)突变体[13]。最近的研究表明, 高粱中的Bmr2编码4CL[14], 玉米中的bm5编码Zm4CL1[8]; 与野生型相比, 这些突变体中木质素含量均有不同程度的降低, 纤维素的消化率和糖化效率也大大提高。因此, 利用4CL改善植物秸秆的利用效率具有重要的实践意义。
迄今为止, 尚未见有关小麦4CL基因鉴定与功能的相关文献。在前期实验中, 本课题组利用RACE技术克隆了一个小麦4CL基因, 因其是小麦中克隆的第一个4CL基因, 故命名为Ta4CL1。为了研究Ta4CL1在植物苯丙烷途径中的功能, 本研究构建了35S:Ta4CL1表达载体并将其转化野生型拟南芥Col-0和拟南芥4CL突变体at4cl1、at4cl3和at4cl14cl3, 以期解析Ta4CL1在苯丙烷途径中的哪个分支途径发挥作用。此外, 本文还研究了Ta4CL1的组织特异表达模式及激素诱导表达模式, 本研究为利用基因工程将Ta4CL1用于改善小麦茎秆性状提供了理论依据。
1 材料与方法
1.1 试验材料
普通小麦(Triticum aestivum L.)品种中国春用于小麦Ta4CL1全长cDNA的克隆、启动子克隆及激素诱导表达分析; 拟南芥(Arabidopsis thaliana L.)哥伦比亚生态型Col-0用于Ta4CL1过表达载体及Ta4CL1pro:Gus载体的转化; 拟南芥4CL突变体种子at4cl1 (WiscDsLox473B01)、at4cl3 (Sail_636_b07)和at4cl14cl3均由Clint Chapple博士惠赠。1.2 Ta4CL1全长cDNA及启动子的克隆
将中国春小麦正常培养15 d, 取其叶片用RNAiso Plus试剂(TaKaRa, 大连)提取总RNA, 用Reverse Transcriptase M-MLV试剂盒(TaKaRa, 大连)反转录成cDNA。3′和5′RACE片段的扩增按SMART RACE cDNA Amplification kit (Clontech, USA)的说明书进行。以此前克隆的一个cDNA片段(500 bp)为基础设计引物Ta4CL1 F1/R1和Ta4CL1 F2/R2分别进行3′RACE反应的第1和第2轮PCR; 设计引物Ta4CL1 F3/R3和F4/R4分别进行5′RACE反应的第1和第2轮PCR。将3′RACE和5′RACE第2轮PCR中的扩增片段克隆至pGEM-T载体(Promega, USA)并经测序验证后, 将5′RACE、3′RACE序列及最初的cDNA序列进行拼接, 得到拼接后的Ta4CL1全长cDNA序列。根据拼接的Ta4CL1全长cDNA序列设计引物Ta4CL1 F5/R5扩增Ta4CL1的全长cDNA, 将扩增产物连接至pGEM-T载体并经测序验证。本研究所用引物序列见表1 (下同)。
Table 1
表1
表1本研究的引物及序列
Table 1
| 引物Primer | 序列Sequence (5′-3′) | 用途Usage |
|---|---|---|
| Ta4CL1 F1 | CCGCGGGGAGCAGATCATGAAAGGTTAC | 3′RACE第1轮PCR 1st run PCR of 3′RACE |
| Ta4CL1 F2 | ACATCAAGAAATTCGTCGCAAAGGAGGTT | 3′RACE第2轮PCR 2nd run PCR of 3′RACE |
| Ta4CL1 R1/R2 | AAGCAGTGGTATCAACGCAGAGTAC(T)30N-1N (N=A, C, G or T; N-1=A, G or C) | 3′RACE |
| Ta4CL1 F3/F4 | (T25)N-1N (N=A, C, G or T; N-1=A, G or C) | 5′RACE |
| Ta4CL1 R3 | AACCTCCTTTGCGACGAATTTCTTGATGT | 5′RACE第1轮PCR 1st run PCR of 5′RACE |
| Ta4CL1 R4 | F: TGTCTCCGGTGTGCAGCCATCCAT | 5′RACE第2轮PCR 2nd run PCR of 5′RACE |
| Ta4CL1 F5/R5 | F: CGCACGCACGCACACGCACAA R: ATCAACATTACACAAGCAGGAAGAACCA | 全长cDNA克隆 Isolation of full-length cDNA |
| Ta4CL1 F6/R6 | F: TCTAGAATGGGGTCTGTGCCGGAG (Xba I) R: ggTACCCTAGCTTGGGATGCCGGC (Kpn I) | Ta4CL1过表达载体构建 Construction of Ta4CL1-overexpression vector |
| Ta4CL1pro F1/R1 | F:CCAGTGCCCTCCATCTCT R:CGCCGCCGCAACCGACTC | 克隆Ta4CL1启动子 Cloning of Ta4CL1 promoter |
| Ta4CL1pro F2/R3 | F: AAGCTTCCAGTGCCCTCCATCTCT (Hind III) R: GAATTCTTCAACGATCGTGGTAGA (EcoR I) | Ta4CL1pro:Gus载体构建 Construction of Ta4CL1 pro:Gus vector |
| GUS | F: GGGCAGGCCAGCGTATCG R: GTCCCGCTAGTGCCTTGTC | 鉴定Ta4CL1pro:Gus载体 Identification of Ta4CL1pro:Gus vector |
| Actin2 (At3g18780) | F: GCTGAGAGATTCAGATGCCC R: CTCGGCCTTGGAGATCCACA | RT-PCR |
| Actin2 (At3g18780) | F: TCGCTGACCGTATGAGCAAAG R: TGTGAACGATTCCTGGACCTG | qRT-PCR |
| AtC3H (At2g40890) | F: GTTGGACTTGACCGGATCTT R: ATTAGAGGCGTTGGAGGATG | qRT-PCR |
| AtCOMT1-1 (At5g54160) | F: TTCCATTGCTGCTCTTTGTC R: CATGGTGATTGTGGAATGGT | qRT-PCR |
| AtCCoAOMT1 (At4g34050) | F: CTCAGGGAAGTGACAGCAAA R: GTGGCGAGAAGAGAGTAGCC | qRT-PCR |
| AtCAD5 (At4g34230) | F: TTGGCTGATTCGTTGGATTA R: ATCACTTTCCTCCCAAGCAT | qRT-PCR |
| AtF5H (At4g36220) | F: CTTCAACGTAGCGGATTTCA R: AGATCATTACGGGCCTTCAC | qRT-PCR |
| AtHCT (At5g48930) | F: GCCTGCACCAAGTATGAAGA R: GACAGTGTTCCCATCCTCCT | qRT-PCR |
| AtCCR1 (At1g15950) | F: GTGCAAAGCAGATCTTCAGG R: GCCGCAGCATTAATTACAAA | qRT-PCR |
| Ta4CL1 F7/R7 | F: TCGTCGCAAAGGAGGTTGTT R: GCGGCCGAGTCTGGCTCT | qRT-PCR |
| TaEF 1a (M90077) | F: CAGATTGGCAACGGCTACG R: CGGACAGCAAAACGACCAAG | qRT-PCR |
新窗口打开|下载CSV
利用Ta4CL1 cDNA序列的ORF在
将Ta4CL1编码的氨基酸序列提交至NCBI网站, 利用Blast搜索其在其他植物物种中的同源蛋白序列, 用MEGA6.0软件按照邻接法(步长值>1000)构建这些4CLs的系统发育树。
1.3 载体构建及转化拟南芥
以连接有Ta4CL1启动子的T载体质粒为模板, 以分别添加Hind III和EcoR I酶切点的引物(Ta4CL1pro F2/R2)进行扩增, 通过Hind III/EcoR I双酶切将Ta4CL1启动子连接至pCAMBIA1391z的相应酶切点, 所得载体经PCR和酶切鉴定后命名为Ta4CL1pro:Gus; 以连接有Ta4CL1 ORF的T载体质粒为模板, 以带有Xba I和Kpn I酶切点的引物(Ta4CL1 F6/R6)进行PCR扩增, 通过Xba I/Kpn I双酶切将Ta4CL1 ORF连接至pCAMBIA1300, 所得克隆经PCR和酶切验证后命名为35S:Ta4CL1, 即Ta4CL1过表达载体。首先将构建好的35S:Ta4CL1和Ta4CL1pro:Gus 载体质粒转化农杆菌菌株GV3101, 然后利用浸花法将其转化野生型拟南芥Col-0[15]。为了深入研究Ta4CL1的功能, 本研究还将35S:Ta4CL1载体分别转化拟南芥4CLs突变体at4cl1、at4cl3和at4cl14cl3。最后, 利用潮霉素筛选、PCR和RT-PCR鉴定每个载体/受体组合的转基因拟南芥纯合株系。
1.4 Ta4CL1表达特性分析
利用潮霉素筛选Ta4CL1pro:Gus拟南芥株系至T3代。将T3代转基因拟南芥幼苗移至营养土中培养2周, 提取转基因拟南芥株系叶片的总RNA, 反转录成cDNA, 利用RT-PCR检测Gus在Ta4CL1pro:Gus拟南芥株系中的表达情况。选取表达量较高的拟南芥株系, 将其正常培养至4周, 参照张双双等[16]的方法对叶和根进行Gus染色; 对正常培养6周的转基因拟南芥的主茎基部进行Gus染色, 观察Ta4CL1在茎中的表达情况。1.5 转基因拟南芥株系的鉴定及表型观察
利用潮霉素筛选各载体/受体转基因拟南芥株系至T3代。将T3代转基因拟南芥幼苗移至营养土中培养2周, 提取各拟南芥株系叶片的总RNA, 反转录成cDNA。以这些cDNA为模板, 利用RT-PCR鉴定Ta4CL1在35S:Ta4CL1株系及Ta4CL1回复拟南芥4cls突变体株系中的表达情况。将野生型拟南芥、35S:Ta4CL1拟南芥株系、拟南芥4cls突变体及其Ta4CL1回复株系正常培养2周(22℃, 16 h光/8 h暗), 拍照记录各拟南芥株系叶片的大小; 将上述各株系正常培养6周后, 照相记录各株系的直立生长情况、测量主茎的株高和直径。每个株系统计测量30棵, 实验重复3次。对测量数据进行One-way ANOVA分析。
1.6 拟南芥株系中木质素、类黄酮和花青素含量测定
将野生型拟南芥、35S:Ta4CL1拟南芥株系、拟南芥4cls突变体及其Ta4CL1回复拟南芥4cls突变体各株系正常培养6周或8周, 取各株系20棵主茎, 65℃烘干过夜, 混合后加液氮研磨成粉末。利用TGA法[17]测定各株系的木质素含量, 各样品的木质素含量以其每毫克样品(干重)在208 nm处的吸光值表示(A280 mg-1 DW)。将上述各拟南芥株系正常培养1个月, 取各株系的莲座叶0.1 g, 经液氮研磨成粉末后, 加80%甲醇, 在4℃浸提24 h, 12,000×g离心30 min; 取上清液900 µL, 加100 µL 10% AlCl3至终浓度为1%。测定各样品在420 nm的吸光值。各样品的类黄酮含量以其每毫克样品(鲜重)的A420表示(A420 mg-1 FW)。
取1月龄各拟南芥株系的莲座叶0.1 g, 经液氮研磨成粉末后, 加1 mL盐酸∶甲醇混合液(HCl∶甲醇=1∶99), 4℃浸提过夜, 12,000×g离心30 min, 分别测定各样品上清液中的A530和A657, 各样品中花青素含量以(A530-A657) g-1 FW表示。上述实验均重复3次, 并进行One-way ANOVA分析。
1.7 4CL酶活性测定
将野生型拟南芥、35S:Ta4CL1拟南芥株系、拟南芥4cls突变体及其Ta4CL1回复拟南芥4cls突变体各株系正常培养1个月, 取各株系主茎100 mg, 参照文献[18]的方法处理样品、测定各样品的OD333, 以不加香豆酸的样品为对照, 按OD333/[样品鲜重(g)×反应时间(h)]计算各样品中4CL 酶活性, 以U h-1g-1 FW表示。实验重复3次, 并进行One-way ANOVA分析。1.8 qRT-PCR分析
将小麦中国春正常培养2周, 将幼苗分成3份, 分别用50 μmol L-1 GA、50 μmol L-1 IAA和50 μmol L-1 MeJA处理0、1、2、4、6、12和24 h。按前述方法提取各样品的总RNA和反转录成cDNA。以SYBR Green I作为荧光染料, 利用荧光定量PCR检测各样品中Ta4CL1的相对表达水平。用2-ΔΔCt法[19]计算在激素处理过程中Ta4CL1相对表达量的变化。以小麦TaEF1a基因(M90077)为内参, 实验重复3次。为了研究Ta4CL1对木质素合成途径4CL下游基因表达的影响, 将野生型拟南芥和Ta4CL1过表达拟南芥株系正常培养6周, 按上述方法提取总RNA和反转录成cDNA, 利用qRT-PCR分析Ta4CL1过表达株系和野生型拟南芥中4CL下游木质素合成途径基因表达的影响, 以拟南芥Actin2 (At3g18780)基因做内参。实验重复3次并进行Student’s t-test分析。
2 结果与分析
2.1 Ta4CL1全长cDNA 和启动子的克隆
以一个500 bp的cDNA片段为基础, 按前述方法分别扩增获得Ta4CL1的5′RACE片段(1697 bp, 图1-A)和3′RACE片段(375 bp, 图1-B); 以拼接后的全长序列为基础设计引物, 利用RT-PCR从小麦中国春中扩增Ta4CL1的全长cDNA序列。Ta4CL1的cDNA全长1925 bp (图1-C), 包含1647 bp的开放阅读框(ORF), 共编码548个氨基酸残基。因其是小麦中克隆的第1个4CL基因, 因此命名为Ta4CL1。图1
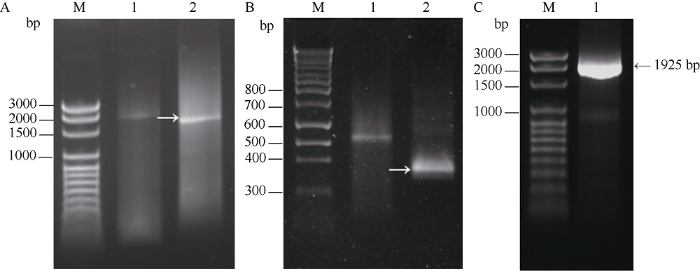 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1利用RACE技术克隆Ta4CL1全长cDNA
扩增Ta4CL1的5′ RACE (A)和3′ RACE (B)片段。1: 第1轮PCR; 2: 第2轮PCR; C: Ta4CL1的全长cDNA的扩增。1: Ta4CL1的全长cDNA。M: marker; 箭头所示为目的条带。
Fig. 1Cloning of full-length cDNA of Ta4CL1 by RACE technique
Amplifying the fragments of 5′ RACE (A) and 3′ RACE (B) of Ta4CL1. 1: the 1st run PCR; 2: the 2nd run PCR; C: amplifying the full-length cDNA of Ta4CL. 1: the full-length cDNA of Ta4CL. M: marker; the arrows point to the target bands.
2.2 Ta4CL1及其同源蛋白的系统发育分析
将Ta4CL1编码的氨基酸序列提交至NCBI网站进行Blast搜索, 下载了Ta4CL1在8个植物物种中的31个同源蛋白序列, 包括单子叶植物水稻(Oryza sativa L.)、玉米(Zea mays L.)、高粱(Sorghum bicolor L.)和柳枝稷(Panicum virgatum L.)以及双子叶植物大豆(Giycine max L.)、烟草(Nicotiana tabacum L.)、拟南芥(Arabidopsis thaliana L.)和毛白杨(Populus tomentosa Carr.)。利用MEGA6的邻接法构建的进化树表明, 这些4CL蛋白可聚成4组, 其中Group I即双子叶植物中的I类4CL; Group II可细分为两个亚族, Subgroup I为双子叶植物中的II类4CL, Subgroup II成员均为单子叶植物中的4CL, 推测其与Subgroup I中的4CL有相似的功能; 包括本文中的Ta4CL1在内的一些单子叶植物中的4CL组成了Group III, 其中, Zm4CL1与Ta4CL1的亲缘关系最近, 其次是Os4CL3和Sb4CL3。据文献报道, Zm4CL1、Os4CL3和Sb4CL3均与木质素的合成密切相关, 推测Ta4CL1也主要参与木质素的生物合成。Group IV的成员均来自单子叶植物(图2), 推测它们参与苯丙烷途径木质素以外的其他酚类物质的合成。图2
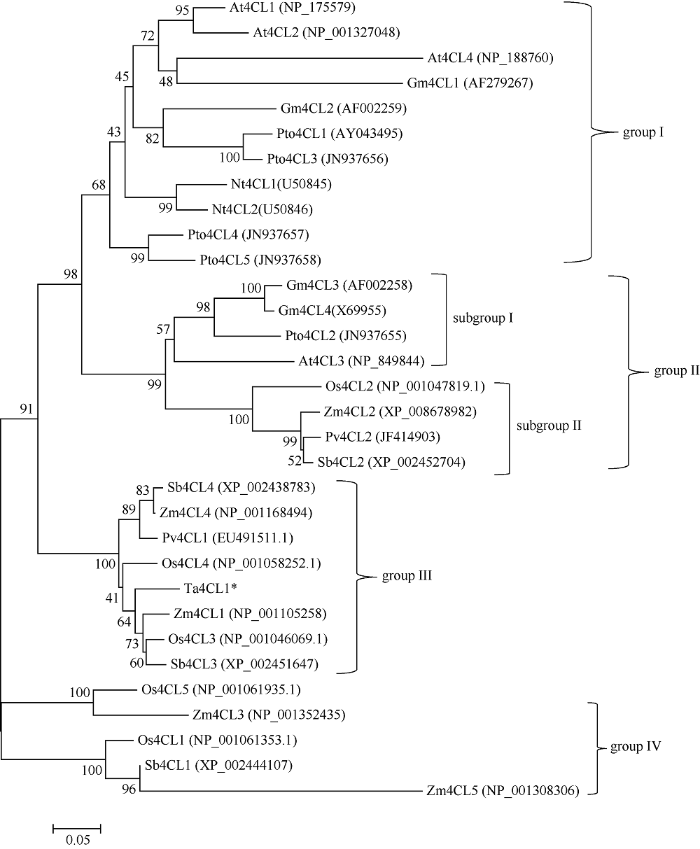 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2Ta4CL1及其同源蛋白的系统发育分析
利用各物种4CL的氨基酸序列构建的无根树(步长值: 1000次重复); 标尺0.05对应于0.05个氨基酸替换/位置。
Fig. 2Phylogeneic tree of Ta4CL1 and its homologues from other plants species
Unrooted phylogenetic tree of 4CL isoforms from plant species is constructed (Bootstrap value: 1000 replicates). The scale bar equals to 0.05 amino acid substitutions per position in the sequence.
2.3 Ta4CL1的表达特性分析
2.3.1 组织表达特性 按前述方法构建Ta4CL1pro:Gus载体并将其转化拟南芥Col-0, 利用Gus染色研究了Ta4CL1的组织表达特性。Gus染色说明Ta4CL1主要在老叶的叶脉中表达(图3-A, B), 在茎的基部表达较强, 在根中的表达较弱(图3-C); Ta4CL1在主茎的木质部也有表达(图3-D)。图3
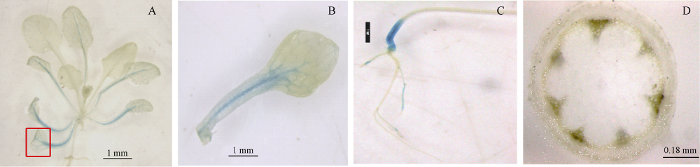 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3利用Ta4CL1pro:Gus转基因拟南芥研究Ta4CL1的组织表达特性
A: 3周大的Ta4CL1pro:Gus转基因拟南芥幼苗的叶; B: A图中方框内叶片的放大图; C: 2周大的Ta4CL1pro:Gus转基因拟南芥幼苗的根; D: Ta4CL1pro:Gus转基因拟南芥主茎(6周)的横切面。A~C图中的标尺表示1 mm; D图中的标尺为0.18 mm。
Fig. 3Tissue-specific expression of Ta4CL1 in Arabidopsis seedlings carrying Ta4CL1pro:Gus
A: the leaves of 3-week-old Arabidopsis seedlings harbouring Ta4CL1pro:Gus; B: enlarged picture of the leave with box in figure A; C: the roots of 2-week-old Arabidopsis seedlings harbouring Ta4CL1pro:Gus; D: the cross section of inflorescence stem of 6-week-old Arabidopsis seedlings harbouring Ta4CL1pro:Gus. Bar: 1 mm (A-C) and 0.18 mm (D).
2.3.2 诱导表达特性 在PlantCARE网站 (http://bioinformatics.psb.ugent.be/webtools/plantcare/ html/)进行的分析表明, Ta4CL1启动子中含有响应MeJA、GA和IAA等激素的元件。因此, 本研究按前述方法用3种激素对正常生长2周的中国春小麦进行处理, 并利用qRT-PCR分析了在不同激素处理过程中Ta4CL1表达的变化。
结果显示, 经GA (50 µmol L-1)处理后, Ta4CL1的表达迅速下降(1 h表达量最低), 此后表达量逐渐升高, 并在4 h达到最高, 之后表达量逐渐减少(图4-A); 经IAA (50 µmol L-1)处理后, Ta4CL1的表达趋势与GA处理相似, 均呈现先下降后上升再下降的趋势, 不同之处在于处理6 h时表达量达到最高(图4-B); 经MeJA (50 µmol L-1)处理后, Ta4CL1表达升高, 在1 h达到最高水平, 此后Ta4CL1的表达基本呈下降趋势(图4-C)。上述结果表明, MeJA、GA和IAA等激素处理均影响Ta4CL1的表达。
图4
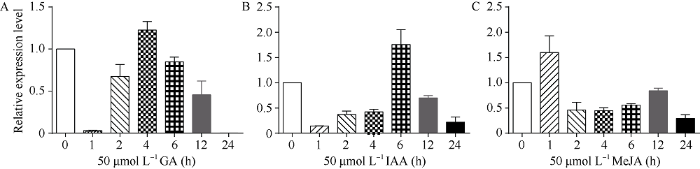 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4GA、IAA和MeJA处理对Ta4CL1表达的影响
Fig. 4Relative expression levels of Ta4CL1 on GA, IAA, and MeJA treatments
2.4 过量表达Ta4CL1促进转基因拟南芥的生长
为系统研究Ta4CL1的功能, 本研究将35S:Ta4CL1表达载体分别转化野生型拟南芥Col-0以及at4cl1、at4cl3和at4cl14cl3等拟南芥4cl突变体, 并通过RT-PCR对各转基因拟南芥纯合株系中Ta4CL1的表达水平进行了鉴定(图5)。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5异源表达Ta4CL1的拟南芥纯合株系RNA水平的鉴定
Fig. 5Identification of homozygous lines harbouring Ta4CL1 in transgenic Arabidopsis seedlings on RNA level
对拟南芥Col-0、Ta4CL1过表达拟南芥株系、at4cl1、at4cl3和at4cl14cl3突变体及其相应的Ta4CL1回复株系的生长和发育过程进行了观察和统计。各株系正常生长至2周时, at4cl1和at4cl14cl3的叶片明显比野生型拟南芥的叶片小, at4cl3的叶片与野生型相比无显著变化, 而Ta4CL1过表达株系及3种突变体相应的功能回复株系的叶片均比野生型大(图6-A)。当各株系正常生长至6周时, at4cl1和at4cl14cl3的植株易倒伏, 而它们相应的功能回复拟南芥株系的茎则保持直立生长; Ta4CL1过表达株系、at4cl3及其相应的功能回复株系则与野生型拟南芥一样呈直立生长状态(图6-B)。测量各拟南芥株系成熟后的株高和直径, 结果表明, 与野生型拟南芥相比, Ta4CL1过表达株系、3种4cls突变体及其功能回复拟南芥株系的株高均没有显著变化(图6-C); 与野生型相比, at4cl1及at4cl14cl3拟南芥的茎更细, at4cl3的直径与野生型相比无显著差异; 但Ta4CL1过表达拟南芥株系及3种突变体相应的功能回复株系的直径均比野生型粗且差异显著(图6-D)。
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6异源表达Ta4CL1的拟南芥株系的表型观察
A: 正常生长2周的拟南芥幼苗; B: 正常生长6周的拟南芥幼苗; C: 各拟南芥株系的株高统计; D: 各拟南芥株系的直径统计; one-way ANOVA分析(字母不同表示差异显著, 含有相同字母表示差异不显著; P < 0.05)。Col-0: 野生型拟南芥; OX-1、-2和-3: 过表达Ta4CL1拟南芥纯合株系; Ta4CL1/at4cl1 L3、L9和L11: 在at4cl背景中过表达Ta4CL1的3个纯合株系; Ta4CL1/at4cl3 L10、L12和L13: 在at4cl3背景中过表达Ta4CL1的3个纯合株系; Ta4CL1/at4cl1 4cl3 L6、L10和L11: 在at4cl1 4cl3背景中过表达Ta4CL1的3个纯合株系。
Fig. 6Ectopic expression of Ta4CL1 affects plant growth in transgenic Arabidopsis seedlings
A: two-week-old Arabidopsis seedlings; B: six-week-old Arabidopsis seedlings; the measurement of plant height (C) and diameter (D) of the inflorescence stem of the eight-week-old Arabidopsis seedlings, One-way ANOVA followed by Duncan’s test, columns with different letters indicate significant differences, dataset share same letter means no significant differences at P < 0.05. Col-0, wild-type Arabidopsis plants; OX-1, OX-2, and OX-3: Ta4CL1-overexpressing Arabidopsis lines; Ta4CL1/at4cl1 L3, L9, and L11: Arabidopsis lines overexpressing Ta4CL1 in the at4cl1 background; Ta4CL1/at4cl3 L10, L12, and L13: Arabidopsis lines overexpressing Ta4CL1 in the at4cl3 background; Ta4CL1/at4cl1 4cl3 L6, L10, and L11: Arabidopsis lines overexpressing Ta4CL1 in the at4cl1 4cl3 background.
上述结果说明过表达Ta4CL1促进了转基因拟南芥叶片的增大和茎的增粗; at4cl1及at4cl14cl3在生长中的缺陷均可通过过表达Ta4CL1得以回复, 即Ta4CL1在促进植物生长中的功能与拟南芥中的At4CL1相似。
2.5 过量表达Ta4CL1的转基因拟南芥中木质素含量增加
为了研究Ta4CL1在木质素合成途径的功能, 本研究使用巯基乙酸法测定了6周龄以及8周龄的各拟南芥株系主茎中的木质素含量。结果表明, 与野生型相比, Ta4CL1过表达拟南芥株系的木质素含量显著增加, 说明Ta4CL1在木质素合成中起正调控作用; at4cl3突变体的木质素含量没有显著变化, 而at4cl1及at4cl14cl3主茎的木质素含量均显著降低(图7-A, B), 说明At4CL1与木质素的合成有关, At4CL3不影响木质素的合成; 在at4cl1和at4cl14cl3中异源表达Ta4CL1均回复了这些突变体中木质素降低的表型, 甚至高于野生型, 这是Ta4CL1过量表达的结果; 在at4cl3中异源表达Ta4CL1得到的结果类似于Ta4CL1过表达的结果。整体而言, 各株系8周龄植株的主茎中木质素含量均显著高于6周龄的植株, 说明木质素在生长发育的后期积累较多。以上结果说明, Ta4CL1与拟南芥中At4CL1的功能相似, 主要参与木质素的生物合成。图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7异源表达Ta4CL1拟南芥株系中木质素含量测定
6周龄(A)和8周龄(B)拟南芥株系茎中木质素含量测定(P < 0.05)。
Fig. 7Determination of total lignin content in the Arabidopsis lines ectopic expressing Ta4CL1
Measurement of total lignin content in stems of 6-week-old (A) and 8-week-old (B) Arabidopsis lines (P < 0.05).
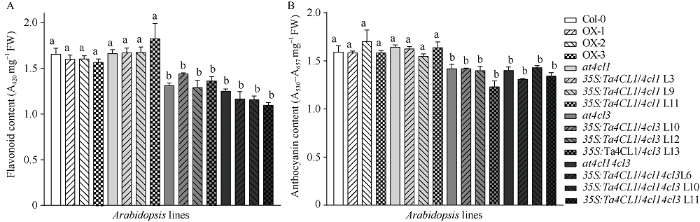
2.6 过量表达Ta4CL1对黄酮类化合物的合成没有显著影响
黄酮合成途径产生各种黄酮类化合物, 包括花青素和类黄酮等。为了研究Ta4CL1对黄酮合成途径的影响, 本实验按前述方法测定了正常生长1个月的各拟南芥株系中类黄酮和花青素含量。结果显示, 与野生型相比, 3个Ta4CL1过表达拟南芥株系的类黄酮和花青素含量均没有显著差异(图8-A, B), 表明Ta4CL1在黄酮合成途径中可能不起作用。与野生型相比, at4cl1突变体及其功能回复株系的类黄酮含量和花青素含量也没有显著变化; 而at4cl3及at4cl14cl3拟南芥的类黄酮和花青素含量均显著降低, 说明At4CL3影响类黄酮和花青素的合成, 而At4CL1在黄酮合成途径中不起作用。与at4cl1相比, at4cl1的功能回复拟南芥株系的类黄酮含量和花青素含量均没有显著差异, 且均与野生型含量相近; 与at4cl3或at4cl14cl3相比, at4cl3和at4cl14cl3各自的功能回复株系中类黄酮含量和花青素含量也均没有显著差异, 仍显著低于野生型水平, 即异源表达Ta4CL1无法回复at4cl3或at4cl14cl3中类黄酮含量和花青素含量降低的表型。上述实验说明, Ta4CL1与拟南芥中At4CL1的功能相似, 在黄酮合成途径中均没有显著作用。
图8
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8异源表达Ta4CL1拟南芥株系中类黄酮和花青素含量测定
正常生长1个月的拟南芥株系的叶片中类黄酮(A)和花青素(B)含量(P < 0.05)。
Fig. 8Determination of flavonoid and anthocyanin content in the Arabidopsis lines ectopic expressing Ta4CL1
Measurement of flavonoid content (A) and anthocyanin content (B) of the one-month-old Arabidopsis seedlings (P < 0.05).
2.7 过表达Ta4CL1 提高了转基因拟南芥中4CL酶活性
4-香豆酸是4CL酶最常见、催化效率最高的底物。因此本研究以4-香豆酸为底物测定了各拟南芥株系的4CL酶活性。按前述方法测定正常生长1个月的野生型、Ta4CL1过表达、3种4cls突变体及其功能回复拟南芥株系的叶片和茎中的4CL酶活性。结果表明, 无论在叶片中还是在茎中, 3个Ta4CL1过表达拟南芥株系的酶活性均显著高于野生型(图9-A, B)。at4cl1及at4cl14cl3拟南芥中4CL酶活性显著低于野生型, 而两种突变体相应的功能回复拟南芥株系的酶活性则回复到野生型水平甚至高于野生型; 与野生型相比, at4cl3突变体的4CL酶活性没有显著差异, 但其功能回复株系中4CL酶活性显著高于野生型, 与Ta4CL1过表达拟南芥株系酶活性水平相当, 说明Ta4CL1与At4CL1在提高转基因拟南芥中4CL酶活性方面的功能相似。整体而言, 各拟南芥株系茎中的4CL酶活性均显著高于其在叶片中的酶活性(图9-A, B), 说明4CL在茎中的酶活性更高, 这与其在木质素合成中的功能是一致的。
图9
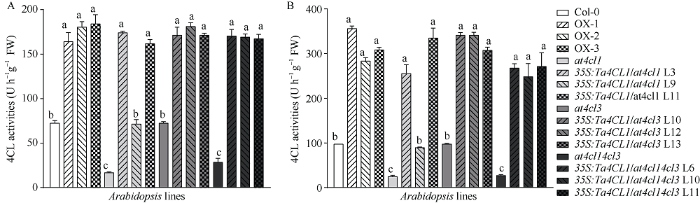 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9异源表达Ta4CL1拟南芥株系中4CL酶活性测定
正常生长4周的拟南芥株系莲座叶(A)和茎(B)中4CL酶活性的测定(P < 0.05)。
Fig. 9Activity of 4CL enzyme in the Arabidopsis lines ectopic expressing Ta4CL1
Measurement of 4CL enzyme activity in rosette leaves (A) and inflorescent stems (B) of 4-week-old Arabidopsis lines ectopic expressing Ta4CL1 (P < 0.05).
2.8 过量表达Ta4CL1影响了木质素合成途径下游基因的表达
为了研究过量表达Ta4CL1对木质素合成途径下游基因表达的影响, 本研究对野生型拟南芥和Ta4CL1过表达拟南芥株系中位于4CL下游、编码7种酶的基因的表达变化进行了分析。结果表明, 与野生型相比, Ta4CL1过表达拟南芥株系中AtCOMT1、AtCAD5和AtCCR1的表达无显著差异, AtCCoAOMT1仅在OX-3株系中有显著差异; 而3个Ta4CL1过表达株系中, AtC3H、AtF5H和AtHCT的表达与野生型间均有显著或极显著差异, 说明Ta4CL1过表达影响了木质素途径下游部分基因的表达。3 讨论
4CL位于苯丙烷途径分支点的上游, 对下游各分支途径具有重要的调控作用。在植物中, 4CL往往以基因家族的形式存在, 不同成员的功能可能有冗余, 也可能有分化。如在拟南芥的基因组中有4个4CL基因, 其中, At4CL1、At4CL2和At4CL4均属于I类4CL, 但At4CL1在木质素生物合成中的贡献最大, 且At4CL1或At4CL2对植物的正常生长是必需的; At4CL4对木质素合成的贡献甚微。At4CL3属于II类4CL, At4CL3突变使拟南芥中黄酮醇苷含量降低了约80%, 而At4CL1和At4CL2的突变没有显著影响黄酮醇苷含量, 但4cl1中总木质素含量显著降低, 说明At4CL1 主要在木质素合成途径起作用, At4CL3主要参与黄酮类化合物的合成[6]。本实验室利用RACE技术克隆了小麦中的第1个4CL1基因Ta4CL1 (图1)。对来自8个物种(4个单子叶植物和4个双子叶植物)的4CL进行的系统发育分析表明, 这些4CL可聚成四大类(图2), group I的成员均来自单子叶植物, 主要参与木质素的合成[6,10]; group II中的subgroup I和subgroup II分别来自单子叶植物和双子叶植物, 这些4CL往往与黄酮类化合物的合成有关[1,6-7]。group III的成员来自单子叶植物, 已有研究表明, 其中的某些成员主要参与木质素的合成[8,12,14]; group IV的成员也均来自单子叶植物, 推测与木质素和黄酮类以外的酚类物质的合成有关。本研究中的Ta4CL1与Zm4CL1以及Os4CL3等参与木质素合成的4CL均聚到group III类, 推测其与木质素的合成密切相关。本研究利用拟南芥4CL突变体at4cl1、at4cl3和at4cl14cl3及Ta4CL1过表达拟南芥株系深入研究了Ta4CL1在苯丙烷各分支途径中的功能。木质素测定结果表明, Ta4CL1过表达拟南芥株系的木质素含量较野生型显著升高, 过表达Ta4CL1能够回复at4cl1及at4cl14cl3中木质素含量降低的表型(图7), 说明Ta4CL1主要影响木质素的合成; 类黄酮和花青素含量的测定结果表明, 过表达Ta4CL1不影响转基因拟南芥中黄酮类化合物的合成; at4cl3及at4cl14cl3中的类黄酮和花青素含量较野生型均显著下降, 但在这两个突变体中过表达Ta4CL1也无法将其回复至野生型水平(图8)。上述结果表明, Ta4CL1与At4CL1功能相似, 主要参与木质素的生物合成, 但不影响黄酮类化合物的合成[6]。在拟南芥中, At4CL1是4CL酶活性的主要贡献者[6]。本研究中, at4cl1和at4cl14cl3中4CL酶活性较野生型均显著降低, 在at4cl1及at4cl14cl3中过表达Ta4CL1也均能回复这些突变体中4CL酶活性降低的表型, 且过表达Ta4CL1拟南芥株系中4CL酶活性显著高于野生型(图9), 说明Ta4CL1与At4CL1类似, 是转基因拟南芥株系中4CL酶活性的主要贡献者。在拟南芥中, AtC3H、AtF5H和AtHCT都对拟南芥的苯丙烷途径及生长发育具有重要的调控功能。AtC3H的突变体ref8表现为植株极度矮小, 且易受真菌侵染; 突变体植株中缺乏芥子酰酯, 但H木质素单体的比例增加[20,21]; AtF5H突变体fah1-2中缺乏S木质素单体[22]; 沉默AtHCT表达的拟南芥植株中木质素合成受到抑制, 株高显著降低[23]。对过表达Ta4CL1拟南芥株系中4CL下游木质素合成相关基因的表达分析表明, 过表达Ta4CL1显著升高了AtC3H、AtF5H和AtHCT等基因的表达(图10), 说明Ta4CL1可能通过影响下游部分基因的表达进而促进了转基因拟南芥株系中木质素的沉积, 并间接影响了转基因植株的生长发育。
图10
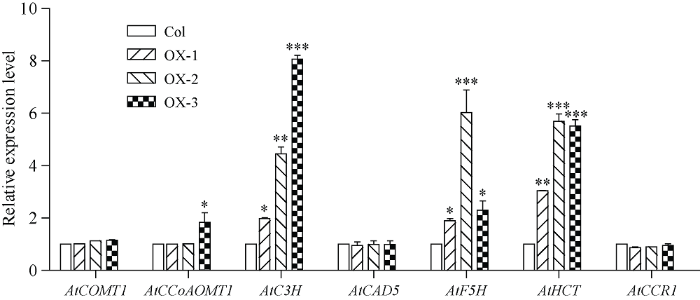 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图10Ta4CL1过表达拟南芥株系中木质素合成途径4CL下游基因的表达分析
Col-0: 野生型拟南芥; OX-1, OX-2, OX-3: Ta4CL1过表达拟南芥株系。Student’s t测验, *P < 0.05, **P < 0.01, ***P < 0.001。
Fig. 10Relative expression levels of genes involving in lignin biosynthesis downstream of 4CL in Ta4CL1-overexpressed Arabidopsis lines
Col-0: wild-type Arabidopsis plants; OX-1, OX-2, and OX-3: Ta4CL1-overexpressing Arabidopsis lines. Student’s t-test, *P<0.05, **P < 0.01, and ***P < 0.001.
研究表明, 降低4CL的表达可能影响转基因株系的生长和/或发育过程, 如4CL的表达降低可导致植株矮小、育性降低等[12,24]。本研究表明, 过量表达Ta4CL1的拟南芥株系叶片增大, 茎更粗; 在at4cl1及at4cl14cl3中过表达Ta4CL1也能回复/改善这些突变体叶片较小、茎较细、易倒伏的性状(图6), 说明Ta4CL1也影响植物的生长发育过程。据报道, 某些4CL的表达还受激素处理、生物/非生物胁迫的诱导。如水稻中的 Os4CL3、Os4CL4和Os4CL5 的表达受伤害的诱导, 而Os4CL1 和 Os4CL2在受到伤害时表达下降; 经过暗适应的水稻在受到紫外线照射时, Os4CL1、Os4CL3、Os4CL4和Os4CL5 的表达均下降, 而Os4CL2 的表达升高[7]; 烟草中的4CL1和4CL2的表达受伤害和MeJA处理的诱导[25]; 在钝鳞紫背苔(Plagiochasma appendiculatum)的叶状体中, Pa4CL1的表达受SA或MeJA处理的诱导[26]。在小麦中, 包括4CL1在内的参与木质素合成途径的很多基因的表达受小麦赤霉病菌(Fusarium graminearum)侵染的诱导[27]; 本研究表明, MeJA、GA和IAA等激素处理均影响Ta4CL1的表达(图4)。这些结果提醒研究者在利用4CL改良植物的木质素含量时应综合考虑因此带来的植物生长发育及适应性的变化。
在利用基因工程改良植物的过程中, 不应严重影响植物正常的生长发育过程, 也不应以牺牲植物对不利环境条件的适应性为代价。本研究表明, Ta4CL1是小麦中重要的4CL基因, 主要参与木质素的生物合成, 是转基因株系中4CL酶活性的主要贡献者; 此外, Ta4CL1还促进植物的生长发育过程, 并可能参与激素及生物胁迫反应。本研究为将Ta4CL1应用于改善植物秸秆的利用效率提供了理论依据。
4 结论
本研究通过RACE技术克隆了小麦Ta4CL1。在转基因拟南芥中的研究表明, Ta4CL1主要参与木质素的生物合成, 基本不影响黄酮类化合物的生物合成; Ta4CL1可能通过调控木质素途径4CL下游某些基因的表达影响木质素的合成; Ta4CL1还影响植物的生长发育及参与对激素处理的响应。致谢:
本文作者诚挚感谢美国普渡大学Clint Chapple博士惠赠的拟南芥at4cl1 (WiscDsLox473B01)、at4cl3 (Sail_636_b07)和at4cl14cl3突变体种子。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 8]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}