 ,*华中农业大学植物科学技术学院, 湖北武汉 430070
,*华中农业大学植物科学技术学院, 湖北武汉 430070Cloning and functional analysis of GmNRT1.2a and GmNRT1.2b in soybean
LI Guo-Ji**, ZHU Lin**, CAO Jin-Shan, WANG You-Ning,*College of Plant Science and Technology, Huazhong Agricultural University, Wuhan 430070, Hubei, China通讯作者:
第一联系人:
收稿日期:2019-10-9接受日期:2020-03-24网络出版日期:2020-07-12
| 基金资助: |
Received:2019-10-9Accepted:2020-03-24Online:2020-07-12
| Fund supported: |
作者简介 About authors
李国纪,E-mail:lgj2389868525@163.com。
朱林,E-mail:onelin@163.com。

摘要
关键词:
Abstract
Keywords:
PDF (1848KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
李国纪, 朱林, 曹金山, 王幼宁. 大豆GmNRT1.2a和GmNRT1.2b基因的克隆及功能探究[J]. 作物学报, 2020, 46(7): 1025-1032. doi:10.3724/SP.J.1006.2020.94152
LI Guo-Ji, ZHU Lin, CAO Jin-Shan, WANG You-Ning.
氮素是植物生长所必需的大量元素之一, 其作为重要成分参与核酸、氨基酸、蛋白质和磷脂及酶的合成, 在植物生长发育及光合作用等多个重要过程中氮素都扮演着主要角色。缺乏氮素会显著影响植物的正常生长, 出现植株矮小和产量降低等诸多现象[1]。在农业生产中, 氮素营养对于粮食生产与农业发展都具有至关重要的作用[2]。
植物吸收土壤中的氮素是通过吸收、同化、转运几个步骤实现的[3]。植物可以吸收2种不同形式的氮, 一是无机氮, 如硝酸盐和氨盐等, 二是有机氮, 如尿素、氨基酸、蛋白质等。硝酸盐和铵盐是植物根系从土壤中获得氮素的最主要的2种形式, 植物根毛通过主动运输的方式吸收并利用NO3-和NH4+, 两者具有不同的理化性质, 因而对植物的生长和代谢也会产生不同的生理效应[4,5]。硝酸盐是植物生长过程中重要的无机氮源, 它不仅作为植物的营养元素, 调控叶面积指数和地上部分干物质累积量[6]以及根系发育等植物生长发育过程[7,8], 也作为一种信号分子, 调节植物生长发育的许多方面, 包括硝酸盐相关的基因表达[9]、根系构型[10]、种子休眠[11]、开花时间[12]等。
众所周知, 豆科作物与其他作物不同, 除可通过主动运输的方式吸收利用氮素营养外, 还可通过其根部特化的组织-根瘤进行共生固氮(symbiotic nitrogen fixation, SNF), 获取氮素营养。共生固氮过程是一个非常复杂的过程, 当土壤中氮元素缺乏时, 豆科植物的根系会释放出类黄酮类物质, 并以此吸引根瘤菌聚集到根系周围, 根瘤菌受到类黄酮类物质的诱导会释放结瘤因子(nod factor, NF)。结瘤因子被豆科植物根部的LysM型受体激酶NFR接受并由此激活NF信号通路。在百脉根(Lotus japonicus)中, 结瘤因子受体为LjNFR1及LjNFR5编码[13,14], 苜蓿(Medicago truncatula)中为MtLYK3/MtLYK4及MtNFP[15]; 在大豆(Glycine max)中编码结瘤因子受体蛋白的同源基因包括GmNFR1α/β及GmNFR5α/β [16,17]。结瘤因子受体接受NF后, 启动下游信号转导级联反应和激活质膜上的激酶(包括LjSYMRK、MtDMI2、GmNORK等), 引起“钙峰”进而导致根瘤菌和植物根系相互作用并形成侵染线。转录因子NIN在侵染线和根瘤原基形成过程中起着重要作用, 它与NSP1和NSP2结合共同调节表皮中的结瘤基因表达和结瘤过程。在豆科植物中相关基因报道已有许多, 但大豆中鉴定到的组分还为数不多, 除了经典的ENOD40[18]之外, 近年来miR172c-NNC1[19]、GmEXPB2[20]、GmPT7[21]、GmBEHL1[22]等已被证明参与调控大豆的结瘤过程。G-protein信号通路相关基因Gα、Gβ、Gγ[23,24]、RGS[24]等也被发现可以介导大豆根瘤的发生发育。与此同时, 近年来的多项研究结果证明, 生长素合成与生长素信号通路多个组分参与大豆根瘤形成过程, 例如GmYUC2a[25]、miR393- GmTIR1/GmAFB3[26]、miR160-ARF10/ ARF16/ARF17[27]和miR167-GmARF8a/GmARF8b[28]等。尽管如此, 对于复杂的菌植互作、根瘤发生发育和共生固氮过程, 还需要通过多种技术手段挖掘其调控的功能基因, 从而全面解析介导大豆共生固氮过程的分子调控网络。
硝酸盐转运蛋白(nitrate transporter, NRT)介导了硝酸盐的吸收、转运和分配, NRT类转运蛋白主要由NRT1和NRT2两个亚家族组成。在拟南芥中, AtNRT1.2属于NRT1亚家族, 并且由于其表达不受NO3-的诱导, 呈组成型表达[29,30]。之前我们已经对大豆GmNRT1.2s有比较详细的生物信息学分析, 相关结果暗示GmNRT1.2s可能参与大豆共生固氮过程。本研究通过对不同组织和不同浓度氮处理下GmNRT1.2a (Glyma.18G126500)和GmNRT1.2b (Glyma. 08G296000)[31]的表达检测, 并利用大豆毛状根转化体系研究二者在大豆结瘤过程中的功能, 证明二者在大豆根系结瘤过程中发挥重要作用。
1 材料与方法
1.1 材料
1.1.1 植物材料 大豆材料是已经完成基因组测序的品种Glycine max var. Williams 82, 后文简称为W82。1.1.2 菌株和质粒 克隆基因所采用的大肠杆菌(E. coli)菌株是DH5α, 大豆毛状根转化所用的发根农杆菌(Agrobacterium rhizogenes)为K599, 大豆慢生型根瘤菌菌株为Bradyrhizobium diaefficiens USDA 110。载体构建涉及到的质粒为T载体Blunt3, 基因过表达载体选用pEGAD。
1.1.3 酶和试剂 KOD plus neo高保真聚合酶购自TOYOBO, 限制性内切酶购自New England Biolabs (NEB), T4连接酶购自TaKaRa公司。
利用Adlab公司的TRI pure试剂提取总RNA, 用购自TaKaRa公司的RNA反转录试剂完成模板cDNA的合成, 选用北京天根公司的SuperReal PreMix Plus (SYBR Green)试剂盒进行荧光定量PCR, 由上海生工生物公司合成引物, 质粒小提试剂盒、DNA凝胶回收试剂盒购自Axygen公司。
1.2 方法
1.2.1 大豆实生苗接种根瘤菌处理 将大豆W82种子用70%的酒精灭菌30 s, 于低氮营养液浸泡的蛭石中萌发, 基础营养液配方参考之前发表文章[32], 其中以Ca(NO3)2·4H2O作为氮源调整氮浓度至0.25 mmol L-1, 期间以水和低氮营养液交替培养。培养室中, 16 h光/8 h暗, 光强140 μmol m-2 s-1, 温度26℃, 相对湿度为70%。大豆萌发15 d后, 将大豆慢生根瘤菌Bradyrhizobium diaefficiens USDA 110 (OD600为0.08)轻轻浇灌在大豆幼苗根系周围, 接种量为每棵30 mL, 分别在接菌后10 d和 28 d取叶片、根和根瘤样品, 于液氮中速冻后保存于-80℃超低温冰箱中备用。1.2.2 大豆实生苗氮浓度梯度处理 根据大豆生长发育过程的需氮量以Ca(NO3)2·4H2O作为氮源, 分别设置了无氮(0N, 0 mmol L-1)、低氮(LN, 0.25 mmol L-1)和高氮(HN, 15.75 mmol L-1) 3个不同浓度的硝酸盐营养液。将W82分别种植在含有不同浓度硝酸盐营养液浸泡的蛭石基质中, 期间补充相同浓度营养液保证处理一致性。于萌发后15 d取主根和侧根样品用于后续表达检测。
1.2.3 结瘤因子提取 结瘤因子是在宿主植物根系分泌的类黄酮作用下, 根瘤菌合成并分泌的一类多糖信号分子(脂壳寡糖), 可以激活结瘤因子信号通路。试验中将根瘤菌Bradyrhizobium diaefficiens USDA 110在28℃, 150 ×g震荡培养, 使OD600达到0.4~0.6制成种子液。取5 mL种子液加入250 mL TY培养基中, 28℃, 150 ×g培养至OD600为0.8~1.0。加入0.25 mL 5 mmol L-1的染料木黄酮。将菌液7000 × g离心10 min去菌体, 上清液经1/5体积正丁醇抽提2次, 1/10体积正丁醇抽提1次, 之后利用旋转蒸发仪在80℃的条件下蒸干正丁醇, 用2 mL蒸馏水溶解, 过滤灭菌后4℃黑暗保存。
1.2.4 大豆实生苗结瘤因子处理 在接种结瘤因子的试验中, 首先于低氮营养液浸泡的蛭石中萌发大豆, 萌发5 d后于大豆根部接种1 mL结瘤因子, 3 d后收集根样品用于分析GmNRT1.2a和GmNRT1.2b的表达。分别用1 mL蒸馏水和1 mL根瘤菌(OD600为0.08)接种的根样品用作对照, 取样方法同上。
1.2.5 总RNA提取及基因表达检测 利用TRI pure试剂提取总RNA, 并且使用RNA反转录试剂完成模板cDNA的合成。根据GmNRT1.2a和GmNRT1.2b基因的CDS序列设计实时荧光定量PCR引物GmNRT1.2a-QF: 5'-TGTTCTTGGCAGGCTCAACT ACT-3', GmNRT1.2a-QR: 5'-CTTCTGGTTCCTTGTTT GCAAT-3', GmNRT1.2b-QF: 5'-TTCACTTAACAGTTGCTTCAACAGTAG-3', GmNRT1.2b-QR: 5'-TCTTA CCCCTTGAGCGTGG-3'。按照北京天根公司的SuperReal PreMix Plus (SYBR Green)试剂盒操作说明, 以GmELF1b基因为内参基因, 引物序列为GmELF1b-QF: 5'-GTTGAAAAGCCAGGGGACA-3', GmELF1b-QR: 5'-TCTTACCCCTTGAGCGTGG-3'。根据相对定量法ΔΔCT公式计算结果。
1.2.6 基因克隆及过重组质粒构建 在构建GmNRT1.2a和GmNRT1.2b过表达载体的过程中, 自Phytozome数据库(http://www.phytozome.net/)获得GmNRT1.2a和GmNRT1.2b的CDS序列, 二者序列长度分别为1758 bp和1785 bp。选择植物表达载体pEGAD作为GmNRT1.2a和GmNRT1.2b的过表达载体, 限制性酶切位点选择其上多克隆位点中的Sma I和Bam HI, 具体引物序列为GmNRT1.2a-F: 5'-TCC CCCGGGATGGAATTAGAACAAAACCAGAG-3', GmNRT1.2a-R: 5'-CGGGATCCTCAGTTGTTTGTAGT TCCTGTCC-3', GmNRT1.2b-F: 5'-TCCCCCGGGAT GGAATTAGAACAAAACCAGAG-3', GmNRT1.2b-R: 5'-CGGGATCCTCAGTTGTTGTTTGTAGTTCCTG-3'。以cDNA为模板扩增目的片段(图1-A), 将目的片段回收纯化后连接T载体进行测序。之后将测序正确的目的基因片段通过酶切酶连的方式连入植物表达载体pEGAD (图1-B)中, 获得GmNRT1.2a和GmNRT1.2b的过表达载体35S::GmNRT1.2a与35S::GmNRT1.2b。将构建完成的重组质粒转化到大肠杆菌DH5α中, 挑取阳性的单克隆接种LB培养基进行扩大培养, 之后保菌并提取质粒转化发根农杆菌K599, 挑取阳性单克隆扩繁, 将菌液以1:1体积与30%甘油混合, 在液氮中速冻后保存于-80℃冰箱备用。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1GmNRT1.2a和GmNRT1.2b过表达载体构建
A: PCR扩增后得到的GmNRT1.2a和GmNRT1.2b目的基因片段; B: pEGAD过表达载体图谱及酶切位点。
Fig. 1Construction of GmNRT1.2a and GmNRT1.2b overexpression vectors
A: the coding sequence of GmNRT1.2a and GmNRT1.2b were obtained through PCR amplification. B: the partial vector map of pEGAD.
1.2.7 大豆毛状根转化及根瘤表型观察 根据先前描述的方法[33,34]及本实验室已报道体系[19], 大豆萌发3~4 d后, 将带有重组质粒的农杆菌K599在3~4 mL LB液体培养基中28℃下150转 min-1震荡培养过夜(OD600为0.8~1.0)。以1:1000的比例将液体活化培养后的菌液接种于50 mL LB液体培养基中进行继代培养, 并在培养基中以1:1000的比例加入质粒及K599所带有的抗生素和0.2 mol L-1 Acetosyringone (AS), 菌液培养至OD600值约0.8后, 将菌液离心, 弃上清液, 用50 mL液体共培培养基(CCM)重悬菌液。挑选子叶完整未受损伤的发芽种子, 用解剖刀从胚轴上切下置于灭过菌的皿中(倒入少量菌液), 在子叶下端的0.1~0.2 cm下胚轴处切开, 切完之后将外植体在菌液中侵染1 h。侵染完成后将外植体放在带有被CCM浸湿的滤纸的灭菌培养皿中, 将其放在暗下共培养3 d后, 把外植体移到低氮浓度营养液浸泡的蛭石中培养, 期间使用低氮营养液与蒸馏水交替培养, 10 d后接种根瘤菌(OD600为0.8~1.0), 每棵接种30 mL, 在接种根瘤菌28 d后, 观察根瘤表型, 拍照并统计根瘤数目, 之后取根样品进行后续的基因表达水平分析。
1.3 数据分析所用软件
借助DNAMAN 7.0设计相关引物, 使用GraphPad Prism 5.0统计根瘤数目, 利用SigmaPlot 10.0软件分析表达结果, 使用SPSS 13.0分析差异显著性。2 结果与分析
2.1 GmNRT1.2a和GmNRT1.2b的组织表达模式分析
为了分析GmNRT1.2a和GmNRT1.2b的组织表达模式, 将W82大豆种子种植到用低氮营养液浸泡的蛭石中, 在其生长到15 d时接种根瘤菌, 分别在接菌后10 d和28 d取叶、根和根瘤不同组织的样品。提取所取样品RNA并反转录成cDNA, 对GmNRT1.2a和GmNRT1.2b基因进行组织表达分析。由图2可知, GmNRT1.2a和GmNRT1.2b在叶、根和瘤中均有表达, 且不同组织中表达量存在较大差异。GmNRT1.2a在10 d和28 d材料中有相同的表达趋势, 表达量根中最高, 叶中其次, 根瘤中最低。在28 d材料的叶、根和根瘤中, GmNRT1.2a的表达量较其在10 d材料中均有显著上升, 在根中表现更为明显。与此相比, 10 d和28 d材料中GmNRT1.2b在叶中表达量明显高于根和根瘤中, 而在28 d材料的叶和根中其表达量都有明显下降, 该结果在叶中表现尤为明显。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2GmNRT1.2a和GmNRT1.2b在大豆不同发育时期的组织表达模式分析
标以不同小写字母的大豆不同组织中对应基因的表达水平在P < 0.05时差异显著。DAI: 接种后天数。
Fig. 2Expression pattern of GmNRT1.2a and GmNRT1.2b in different developmental stages of soybean
Mean values indexed with different letters are significantly different at P < 0.05. DAI: days after inoculation.
2.2 GmNRT1.2a和GmNRT1.2b基因受硝酸盐诱导表达
为了验证不同浓度硝酸盐对GmNRT1.2a和GmNRT1.2b表达的影响, 本研究对大豆W82材料分别进行0、0.25和15.75 mmol L-1硝酸盐处理。取在不同氮浓度中萌发15 d后的主根和侧根的混合样品, 检测GmNRT1.2a和GmNRT1.2b的基因表达。由图3可知, 与其在无氮营养液中的表达水平相比, 随着氮浓度的升高, GmNRT1.2a和GmNRT1.2b的表达量有明显增加。表明GmNRT1.2a和GmNRT1.2b受硝酸盐尤其是高浓度硝酸盐的显著诱导表达。图3
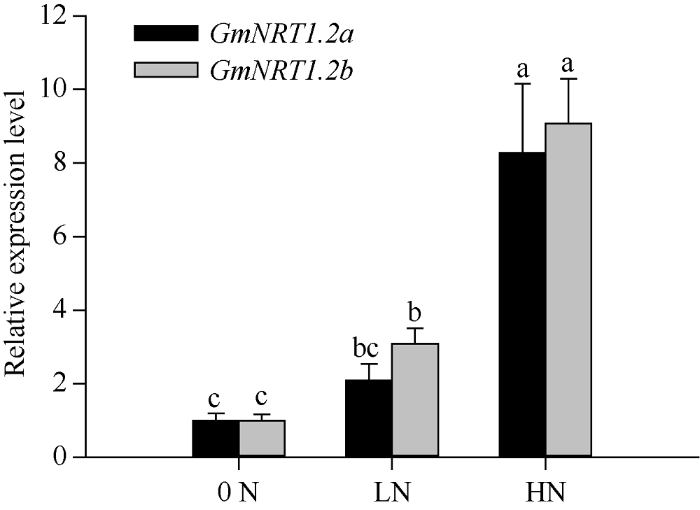 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3GmNRT1.2a和GmNRT1.2b在不同浓度下的硝酸盐下的表达模式
大豆在0N (0 mmol L-1)、LN (0.25 mmol L-1)和HN (15.75 mmol L-1) 3种硝酸盐浓度梯度下生长到15 d, 取大豆根部组织样品。用荧光定量PCR检测GmNRT1.2a (A)和GmNRT1.2b (B)的表达, GmELF1b为内参基因。图中标以不同字母的基因表达水平在P < 0.05时差异显著。
Fig. 3Expression pattern of GmNRT1.2a and GmNRT1.2b under different concentrations of nitrate
Soybeans were germinated in vermiculite at different nitrate concentrations (0N-0 mmol L-1, LN-0.25 mmol L-1, HN-15.75 mmol L-1). The roots were collected at 15 days after germination. The expression of GmNRT1.2a (A) and GmNRT1.2b (B) was analyzed by RT-qPCR. Bars with different lowercase letters in each figure are significantly different at P < 0.05.
2.3 GmNRT1.2a和GmNRT1.2b响应根瘤菌及结瘤因子的表达模式分析
为了分析GmNRT1.2a和GmNRT1.2b是否响应根瘤菌侵染, 本研究将W82材料种植在用低氮营养液浸泡的蛭石中, 于萌发后5 d每棵材料接种1 mL结瘤因子, 并以同时间点接种等体积根瘤菌或蒸馏水作为对照, 在处理3 d后取主根和侧根的混合样品, 分别对GmNRT1.2a和GmNRT1.2b进行表达分析, 并且以之前已报道的响应结瘤因子信号的GmENOD40-1作为阳性对照。由图4可知, GmENOD40-1同时受到根瘤菌和结瘤因子诱导, 且受根瘤菌诱导更强。GmNRT1.2a和GmNRT1.2b的表达同时受到根瘤菌和结瘤因子诱导, 与其对根瘤菌侵染的响应模式相比, 二者受结瘤因子的诱导更为明显。推测它们可能通过经典的结瘤因子信号通路响应根瘤菌的侵染。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4GmNRT1.2a和GmNRT1.2b响应接种根瘤菌和结瘤因子表达模式分析
标以不同小写字母的基因表达水平在根瘤菌和结瘤因子处理的大豆植株与对照间在P < 0.05水平上差异显著。
Fig. 4Expression patterns of GmNRT1.2a and GmNRT1.2b in response to rhizobium inoculation and nod factors treatment
Values followed by different letters are significantly different at P < 0.05.
2.4 过表达GmNRT1.2a和GmNRT1.2b增加根瘤数目
为了进一步探究GmNRT1.2a和GmNRT1.2b对根瘤数目的影响, 利用毛状根转化体系分别将带有GmNRT1.2a和GmNRT1.2b过表达重组质粒转化大豆, 以空载体转化作为对照(EV)。在接种根瘤菌28 d后观察根瘤表型并取样, 由图5可知, GmNRT1.2a或GmNRT1.2b过表达材料中, 其表达量同对照相比均显著升高(图5-A, B)。空载体对照(EV)转化的毛状根的根瘤平均数目为15.38, 而在过表达GmNRT1.2a与GmNRT1.2b的阳性毛状根中, 其平均结瘤数分别为39.60个与24.63个(图5-C, D)。说明低氮条件下过表达GmNRT1.2a或GmNRT1.2b都可以极大地增加大豆根瘤数目, 其中过表达GmNRT1.2a根瘤数目增加更明显, 表明GmNRT1.2a及GmNRT1.2b可能正向调控大豆结瘤。图5
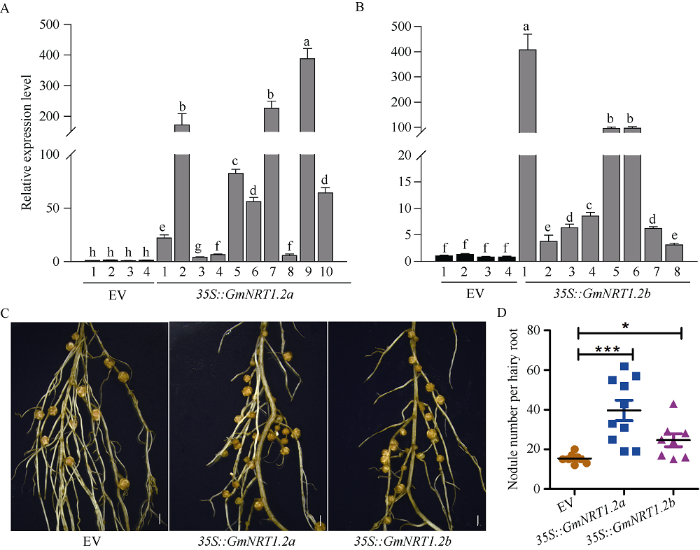 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5在低氮培养条件下过表达GmNRT1.2a或GmNRT1.2b增加结瘤数目
A, B: 在接种根瘤菌28 d后, 利用荧光定量PCR检测EV、35S::GmNRT1.2a和35S::GmNRT1.2b单条毛状根表达, 以GmELF1b基因作为内参基因。标以不同小写字母的柱值在不同转基因大豆植株间在P < 0.05时的差异显著。C: EV、35S::GmNRT1.2a和35S::GmNRT1.2b的单条根的根瘤表型, Bar = 5 mm。D: 统计EV、35S::GmNRT1.2a和35S::GmNRT1.2b的根瘤数目。*和***分别表示转基因株系35S::GmNRT1.2a和35S::GmNRT1.2b的根瘤数目与对照在P < 0.05和P < 0.001水平差异显著。
Fig. 5Nodule number increased by 35S::GmNRT1.2a or 35S::GmNRT1.2b under low nitrate condition
A, B: RT-qPCR analysis of GmNRT1.2a or GmNRT1.2b in single hairy root transformed with empty vector, 35S::GmNRT1.2a or 35S::GmNRT1.2b inoculated with Bradyrhizobium japonicum USDA110 at 10 and 28 days treatment. GmELF1b was used as an endogenous control for gene expression. Bars with different lowercase letters in each figure are significantly different at P < 0.05. C: the phenotype of nodule per hairy root transformed with empty vector (EV), 35S::GmNRT1.2a and 35S::GmNRT1.2b at 28 DAI. Bar = 5 mm. D: nodule number per hairy root transformed with empty vector (EV), 35S::GmNRT1.2a or 35S::GmNRT1.2b were counted at 28 DAI. The values followed by * and *** are significantly different at P < 0.05 and P < 0.001.
3 讨论
近年来, 有关硝酸盐转运蛋白参与氮素营养吸收的研究已成为热点。目前的研究主要集中在拟南芥[35]、水稻[36,37,38,39]、番茄[40]、小麦[41]、油菜[42]等植物, 对于大豆中该类蛋白的功能研究报道较少, 尚处于起步阶段。由于物种的特异性, 不同物种的同源基因之间可能存在着功能的分化。大豆GmNRT1.2s 的相关生物信息学分析结果也显示, 大豆GmNRT1.2s的碱基和氨基酸序列与拟南芥中的NRT1.2存在一定差异[31]。另外, 拟南芥中ATNRT1.2s为组成型表达, 即不受硝酸盐的诱导[29,30], 而本研究中发现大豆GmNRT1.2a和GmNRT1.2b同时受到低浓度和高浓度硝酸盐诱导。除此之外, 大豆GmNRT1.2a在根中表达量最高, 而GmNRT1.2b主要在叶中表达, 二者主要的表达部位有着明显的不同, 这一差异也在一定程度上暗示着二者可能在某些功能上存在差异。相关研究结果证明, GmNRT1.2a和GmNRT1.2b过表达后导致大豆的根瘤数目均显著增加, 但是对于GmNRT1.2a和GmNRT1.2b的功能研究尚不完全。在之后的研究中可以利用RNA干扰(RNA interference, RNAi)技术和CRISPR基因编辑技术对GmNRT1.2a和GmNRT1.2b的功能进行深入探究, 建立快速稳定的RNAi与CRISPR大豆毛状根转化体系, 对进一步研究GmNRT1.2a和GmNRT1.2b在结瘤过程中的作用具有重要意义。
基于大豆GmNRT1.2a和GmNRT1.2b受高浓度硝酸盐诱导, 并且在接种根瘤菌和结瘤因子后表达上调, 推测它们可能受某些转录因子调控, 后续研究可以通过生物信息学分析预测其上游转录因子, 并通过酵母单杂技术进行文库筛选候选的转录因子, 进一步利用ChIP-PCR与EMSA技术验证转录因子对GmNRT1.2a和GmNRT1.2b的调控。为了完善对GmNRT1.2a和GmNRT1.2b调控大豆结瘤的分子机制研究, 还可以开展其互作蛋白的预测及鉴定, 例如, 通过string 10 (https://string-db.org/)网站进行互作预测, 同时利用酵母双杂交及IP-MS大规模寻找可能与GmNRT1.2a和GmNRT1.2b互作的蛋白, 从而更深入地解析GmNRT1.2a和GmNRT1.2b调控大豆结瘤的分子网络。
4 结论
GmNRT1.2a和GmNRT1.2b分别在大豆根和叶中表达最高。GmNRT1.2a和GmNRT1.2b受硝酸盐诱导表达, 且显著响应根瘤菌及结瘤因子处理。在毛状根转化体系中, 过表达GmNRT1.2a和GmNRT1.2b使大豆根瘤数目显著增加, 表明GmNRT1.2a和GmNRT1.2b在结瘤过程中发挥正向调控作用。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
DOI:10.7606/j.issn.1009-1041.2013.04.016URL [本文引用: 1]

为探讨豫北地区合理的氮肥施用模式,以周麦18和济麦22为材料,分析了拔节期追氮对冬小麦籽粒产量、经济效益及氮素吸收和利用的影响。结果表明,在底施纯氮120 kg·hm-2的基础上,随拔节期追氮量的增加,籽粒产量呈先增后降趋势,籽粒产量与追氮量之间可用二次曲线方程进行拟合。周麦18和济麦22在追氮140 kg·hm-2(N260)处理下籽粒产量和经济效益较高。随追氮量的增加,植株氮素积累总量增长缓慢且有下降趋势,籽粒氮素收获指数、追施氮素利用效率和氮肥偏生产力显著降低,追施氮肥农学利用效率呈先升后降变化趋势。拔节期追氮促进了营养器官氮素向籽粒中转运。各追氮处理下,周麦18和济麦22籽粒氮素分别有81.53%~88.62%和79.65%~89.10%来自营养器官氮素的转运。综合来看,底施氮肥120 kg·hm-2结合拔节期追氮140~180 kg·hm-2是豫北地区小麦高产高效的氮肥施用模式。
DOI:10.7606/j.issn.1009-1041.2013.04.016URL [本文引用: 1]
为探讨豫北地区合理的氮肥施用模式,以周麦18和济麦22为材料,分析了拔节期追氮对冬小麦籽粒产量、经济效益及氮素吸收和利用的影响。结果表明,在底施纯氮120 kg·hm-2的基础上,随拔节期追氮量的增加,籽粒产量呈先增后降趋势,籽粒产量与追氮量之间可用二次曲线方程进行拟合。周麦18和济麦22在追氮140 kg·hm-2(N260)处理下籽粒产量和经济效益较高。随追氮量的增加,植株氮素积累总量增长缓慢且有下降趋势,籽粒氮素收获指数、追施氮素利用效率和氮肥偏生产力显著降低,追施氮肥农学利用效率呈先升后降变化趋势。拔节期追氮促进了营养器官氮素向籽粒中转运。各追氮处理下,周麦18和济麦22籽粒氮素分别有81.53%~88.62%和79.65%~89.10%来自营养器官氮素的转运。综合来看,底施氮肥120 kg·hm-2结合拔节期追氮140~180 kg·hm-2是豫北地区小麦高产高效的氮肥施用模式。
URLPMID:20299346 [本文引用: 1]
URL [本文引用: 1]
A hydroponic experiment was conducted to study the efiect of partial replacement of NO3--N by NH4+-N on the seedling growth and organic acid content of tomato (Lycopersicon esculentum Mill). A completely randomized design was established with three replications and five treatments, i.e., NO3--N/NH4+-N of 100/0, 75/25, 50/50, 25/75 and 0/100. Results showed that 25% replacement of NO3--N by NH4+-N significantly (P = 0.05) improved fresh and dry weight, revealing that a proper percentage of NH4+-N was important for tomato nitrogen nutrition. This could increase the plant growth even though tomato was a crop that preferred nitrate nutrition. Also an increase in the proportion of NH4+-N in the nutrient solution led to a significant decrease (P = 0.05) in malate, citrate and fumarate. However, the 25% NH4+-N plus 75% NO3--N treatment had no significant effect (P = 0.05) on the 2-ketoglutarate, succinate or oxalic acid content, showing that only some organic acids in tomato plants were affected. Only pyruvate increased significantly (P = 0.05), and it only increased for 25% and 50% replacement of NO3--N by NH4+-N. Metabolism of these organic acids, especially malate, citrate and fumarate, should be further studied at the molecular level in vegetables applied with different nitrogen forms.
DOI:10.1023/B:HYDR.0000025052.81373.f3URL [本文引用: 1]
URL [本文引用: 1]
为探讨不同滴灌施肥水平对春玉米产量及水肥利用效率的影响,应用滴灌施肥技术于2016和2017年在宁夏旱作节水科技园区试验站开展大田春玉米小区试验。以"先玉335"为试验材料,设置4个灌水水平(D75:75%ETc、D90:90%ETc、D105:105%ETc、D120:120%ETc,ETc为玉米需水量)和4个N-P2O5-K2O施肥水平:2016年为60-30-30 kg/hm2(F60)、120-60-60 kg/hm2(F120)、180-90-90 kg/hm2(F180)、240-120-120 kg/hm2(F240),2017年为150-70-70 kg/hm2(F150)、225-110-110 kg/hm2(F225)、300-150-150 kg/hm2(F300)、375-180-180 kg/hm2(F375),以1个充分灌水(120%ETc)无肥为对照(CK),共17个处理。研究不同水肥供应对春玉米株高、茎粗、叶面积指数(leaf area index,LAI)、地上部干物质累积量和产量的影响,并分析其水肥利用效率。2 a试验结果表明:灌水量和施肥量单因素对玉米株高、茎粗、LAI都有显著或极显著的影响,灌水量和施肥量耦合效应对玉米株高有极显著的影响;灌水量和施肥量对玉米成熟期地上部干物质的影响随着2 a施肥梯度的不同而有所差异,在低肥梯度的2016年,灌水量和施肥量对地上部干物质累积有显著的影响,其中D120F180处理籽粒地上部干物质最大,为12 691 kg/hm2,在高肥梯度的2017年,随着灌水量和施肥量的增加,75%ETc和105%ETc处理的地上部干物质累积量有先增加后减小的趋势,D90F300处理下籽粒地上部干物质累积量最大,为14 912 kg/hm2;在低肥梯度的2016年,灌水施肥量对春玉米产量有显著影响,D120F240处理产量最高,为14 400 kg/hm2,而在高肥梯度的2017年,D90F300处理玉米产量最高,为16 884 kg/hm2;2 a试验结果表明灌水量和施肥量对春玉米水分利用效率和肥料偏生产力都有极显著影响。基于春玉米产量和水分利用效率最大值的95%为置信区间优化水肥管理方案,兼顾节水节肥,推荐灌水量在323~446 mm、N-P2O5-K2O施肥量在210-104-104~325-163-163 kg/hm2。该研究结果对宁夏春玉米滴灌施肥管理具有重要指导意义。
URL [本文引用: 1]
为探讨不同滴灌施肥水平对春玉米产量及水肥利用效率的影响,应用滴灌施肥技术于2016和2017年在宁夏旱作节水科技园区试验站开展大田春玉米小区试验。以"先玉335"为试验材料,设置4个灌水水平(D75:75%ETc、D90:90%ETc、D105:105%ETc、D120:120%ETc,ETc为玉米需水量)和4个N-P2O5-K2O施肥水平:2016年为60-30-30 kg/hm2(F60)、120-60-60 kg/hm2(F120)、180-90-90 kg/hm2(F180)、240-120-120 kg/hm2(F240),2017年为150-70-70 kg/hm2(F150)、225-110-110 kg/hm2(F225)、300-150-150 kg/hm2(F300)、375-180-180 kg/hm2(F375),以1个充分灌水(120%ETc)无肥为对照(CK),共17个处理。研究不同水肥供应对春玉米株高、茎粗、叶面积指数(leaf area index,LAI)、地上部干物质累积量和产量的影响,并分析其水肥利用效率。2 a试验结果表明:灌水量和施肥量单因素对玉米株高、茎粗、LAI都有显著或极显著的影响,灌水量和施肥量耦合效应对玉米株高有极显著的影响;灌水量和施肥量对玉米成熟期地上部干物质的影响随着2 a施肥梯度的不同而有所差异,在低肥梯度的2016年,灌水量和施肥量对地上部干物质累积有显著的影响,其中D120F180处理籽粒地上部干物质最大,为12 691 kg/hm2,在高肥梯度的2017年,随着灌水量和施肥量的增加,75%ETc和105%ETc处理的地上部干物质累积量有先增加后减小的趋势,D90F300处理下籽粒地上部干物质累积量最大,为14 912 kg/hm2;在低肥梯度的2016年,灌水施肥量对春玉米产量有显著影响,D120F240处理产量最高,为14 400 kg/hm2,而在高肥梯度的2017年,D90F300处理玉米产量最高,为16 884 kg/hm2;2 a试验结果表明灌水量和施肥量对春玉米水分利用效率和肥料偏生产力都有极显著影响。基于春玉米产量和水分利用效率最大值的95%为置信区间优化水肥管理方案,兼顾节水节肥,推荐灌水量在323~446 mm、N-P2O5-K2O施肥量在210-104-104~325-163-163 kg/hm2。该研究结果对宁夏春玉米滴灌施肥管理具有重要指导意义。
DOI:10.3389/fpls.2017.02096URLPMID:29312372 [本文引用: 1]
Root is important in acquiring nutrients from soils. Developing marker-assisted selection for wheat root traits can help wheat breeders to select roots desirable for efficient acquisition of nutrients. A recombinant inbred line (RIL) population derived from wheat varieties Xiaoyan 54 and Jing 411 was used to detect QTLs for maximum root length and root dry weight (RDW) under control, low nitrogen and low phosphorus conditions in hydrophobic culture (HC). We totally detected 17 QTLs for the investigated root traits located at 13 loci on 11 chromosomes. These loci differentially expressed under different nutrient supplying levels. The RILs simultaneously harboring positive alleles or negative alleles of the most significant three QTLs for RDW, qRDW.CK-2A, qRDW.CK-2D, and qRDW.CK-3B, were selected for soil column culture (SC) trial to verify the effects of these QTLs under soil conditions. The RILs pyramiding the positive alleles not only had significantly higher shoot dry weight, RDW, nitrogen and phosphorus uptake in all the three treatments of the HC trial, but also had significantly higher RDW distribution in both the top- and sub-soils in the SC trial than those pyramiding the negative alleles. These results suggested that QTL analysis based on hydroponic culture can provide useful information for molecular design of wheat with large and deep root system.
DOI:10.1111/j.1365-313X.2008.03443.xURLPMID:18266918 [本文引用: 1]
Arabidopsis root architecture is highly responsive to changes in the nitrogen supply. External NO(3)(-) stimulates lateral root growth via a signalling pathway involving the ANR1 MADS box transcription factor, while the presence of exogenous l-glutamate (Glu) at the primary root tip slows primary root growth and stimulates root branching. We have found that NO(3)(-), in conjunction with Glu, has a hitherto unrecognized role in regulating the growth of primary roots. Nitrate was able to stimulate primary root growth, both directly and by antagonising the inhibitory effect of Glu. Each response depended on direct contact between the primary root tip and the NO(3)(-), and was not elicited by an alternative N source (NH(4)(+)). The chl1-5 mutant, which is defective in the NRT1.1 (CHL1) NO(3)(-) transporter, was insensitive to NO(3)(-) antagonism of Glu signalling, while an anr1 mutant retained its sensitivity. Sensitivity to NO(3)(-) was restored in a chl1-5 mutant constitutively expressing NRT1.1. However, expression in chl1-5 of a transport-competent but non-phosphorylatable form of NRT1.1 not only failed to restore NO(3)(-) sensitivity but also had a dominant-negative effect on Glu sensitivity. Our results indicate the existence of a NO(3)(-) signalling pathway at the primary root tip that can antagonise the root's response to Glu, and they further suggest that NRT1.1 has a direct NO(3)(-) sensing role in this pathway. We discuss how the observed signalling interactions between NO(3)(-) and Glu could provide a mechanism for modulating root architecture in response to changes in the relative abundance of organic and inorganic N.
DOI:10.1104/pp.103.021253URLPMID:12805587 [本文引用: 1]
The genomic response to low levels of nitrate was studied in Arabidopsis using the Affymetrix ATH1 chip containing more than 22,500 probe sets. Arabidopsis plants were grown hydroponically in sterile liquid culture on ammonium as the sole source of nitrogen for 10 d, then treated with 250 microm nitrate for 20 min. The response to nitrate was much stronger in roots (1,176 genes showing increased or decreased mRNA levels) than in shoots (183 responding genes). In addition to known nitrate-responsive genes (e.g. those encoding nitrate transporters, nitrate reductase, nitrite reductase, ferredoxin reductase, and enzymes in the pentose phosphate pathway), genes encoding novel metabolic and potential regulatory proteins were found. These genes encode enzymes in glycolysis (glucose-6-phosphate isomerase and phosphoglycerate mutase), in trehalose-6-P metabolism (trehalose-6-P synthase and trehalose-6-P phosphatase), in iron transport/metabolism (nicotianamine synthase), and in sulfate uptake/reduction. In many cases, only a few select genes out of several in small gene families were induced by nitrate. These results show that the effect of nitrate on gene expression is substantial (affecting almost 10% of the genes with detectable mRNA levels) yet selective and affects many genes involved in carbon and nutrient metabolism.
DOI:10.1016/j.pbi.2014.06.004URLPMID:24997289 [本文引用: 1]
Root system architecture is a fundamentally important trait for resource acquisition in both ecological and agronomic contexts. Because of the plasticity of root development and the almost infinite complexity of the soil, root system architecture is shaped by environmental factors to a much greater degree than shoot architecture. In attempting to understand how roots sense and respond to environmental cues, the striking effects of nitrate and other forms of nitrogen on root growth and branching have received particular attention. This minireview focuses on the latest advances in our understanding of the diverse nitrogen signalling pathways that are now known to act at multiple stages in the process of lateral root development, as well as on primary root growth.
DOI:10.1111/j.1365-3040.2005.01292.xURLPMID:16229082 [本文引用: 1]
Nitrate is an important nitrogen source for plants, but also a signal molecule that controls various aspects of plant development. In the present study the role of nitrate on seed dormancy in Arabidopsis was investigated. The effects of either mutations affecting the Arabidopsis nitrate reductase genes or of different nitrate regimes of mother plants on the dormancy of the seeds produced were analysed. Altogether, data show that conditions favouring nitrate accumulation in mother plants and in seeds lead to a lower dormancy of seeds with little other morphological or biochemical differences. Analysis of germination during seed development indicated that nitrate does not prevent the onset of dormancy but rather its maintenance. The effect of an exogenous supply of nitrate on seed germination was tested: nitrate in contrast to glutamine or potassium chloride clearly stimulated the germination of dormant seeds. Data show, moreover, that the Arabidopsis dual affinity nitrate transporter NRT1.1 (CHL1) may be involved in conveying the nitrate signal into seeds. Thus, nitrate provided exogenously or by mother plants to the produced seeds, acts as a signal molecule favouring germination in Arabidopsis. This signalling may involve interaction with the abscisic acid or gibberellin pathway.
URLPMID:21113723 [本文引用: 1]
DOI:10.1038/nature02045URLPMID:14534591 [本文引用: 1]
Plants belonging to the legume family develop nitrogen-fixing root nodules in symbiosis with bacteria commonly known as rhizobia. The legume host encodes all of the functions necessary to build the specialized symbiotic organ, the nodule, but the process is elicited by the bacteria. Molecular communication initiates the interaction, and signals, usually flavones, secreted by the legume root induce the bacteria to produce a lipochitin-oligosaccharide signal molecule (Nod-factor), which in turn triggers the plant organogenic process. An important determinant of bacterial host specificity is the structure of the Nod-factor, suggesting that a plant receptor is involved in signal perception and signal transduction initiating the plant developmental response. Here we describe the cloning of a putative Nod-factor receptor kinase gene (NFR5) from Lotus japonicus. NFR5 is essential for Nod-factor perception and encodes an unusual transmembrane serine/threonine receptor-like kinase required for the earliest detectable plant responses to bacteria and Nod-factor. The extracellular domain of the putative receptor has three modules with similarity to LysM domains known from peptidoglycan-binding proteins and chitinases. Together with an atypical kinase domain structure this characterizes an unusual receptor-like kinase.
URLPMID:14534578 [本文引用: 1]
URLPMID:16844829 [本文引用: 1]
URLPMID:21175888 [本文引用: 1]
DOI:10.1093/pcp/pcp178URLPMID:20007291 [本文引用: 1]
Chemically induced non-nodulating nod139 and nn5 mutants of soybean (Glycine max) show no visible symptoms in response to rhizobial inoculation. Both exhibit recessive Mendelian inheritance suggesting loss of function. By allele determination and genetic complementation in nod139 and nn5, two highly related lipo-oligochitin LysM-type receptor kinase genes in Glycine max were cloned; they are presumed to be the critical nodulation-inducing (Nod) factor receptor similar to those of Lotus japonicus, pea and Medicago truncatula. These duplicated receptor genes were called GmNFR5alpha and GmNFR5beta. Nonsense mutations in GmNFR5alpha and GmNFR5beta were genetically complemented by both wild-type GmNFR5alpha and GmNFR5beta in transgenic roots, indicating that both genes are functional. Both genes lack introns. In cultivar Williams82 GmNFR5alpha is located in chromosome 11 and in tandem with GmLYK7 (a related LysM receptor kinase gene), while GmNFR5beta is in tandem with GmLYK4 in homologous chromosome 1, suggesting ancient synteny and regional segmental duplication. Both genes are wild type in G. soja CPI100070 and Harosoy63; however, a non-functional NFR5beta allele (NFR5beta*) was discovered in parental lines Bragg and Williams, which harbored an identical 1,407 bp retroelement-type insertion. This retroelement (GmRE-1) and related sequences are located in several soybean genome positions. Paradoxically, putatively unrelated soybean cultivars shared the same insertion, suggesting a smaller than anticipated genetic base in this crop. GmNFR5alpha but not GmNFR5beta* was expressed in inoculated and uninoculated tap and lateral root portions at about 10-25% of GmATS1 (ATP synthase subunit 1), but not in trifoliate leaves and shoot tips.
DOI:10.1073/pnas.022664799URLPMID:11842184 [本文引用: 1]
ENOD40 is expressed at an early stage in root nodule organogenesis in legumes. Identification of ENOD40 homologs in nonleguminous plants suggests that this gene may have a more general biological function. In vitro translation of soybean ENOD40 mRNA in wheat germ extracts revealed that the conserved nucleotide sequence at the 5' end (region I) encodes two peptides of 12 and 24 aa residues (peptides A and B). These peptides are synthesized de novo from very short, overlapping ORFs. Appropriate ORFs are present in all legume ENOD40s studied thus far. In this case small peptides are directly translated from polycistronic eukaryotic mRNA. The 24-aa peptide B was detected in nodules by Western blotting. Both peptides specifically bind to the same 93-kDa protein, which was affinity purified from soybean nodules. Using peptide mass fingerprinting, we identified this binding protein as nodulin 100, which is a subunit of sucrose synthase. Based on our data we suggest that ENOD40 peptides are involved in the control of sucrose use in nitrogen-fixing nodules.
[本文引用: 2]
URLPMID:26432877 [本文引用: 1]
DOI:10.1111/nph.15541URLPMID:30317659 [本文引用: 1]
Symbiotic nitrogen (N2 ) fixation plays a vital role in sustainable agriculture. Efficient N2 fixation requires various materials, including phosphate (Pi); however, the molecular mechanism underlying the transport of Pi into nodules and bacteroids remains largely unknown. A nodule-localized Pi transporter, GmPT7, was functionally characterized in soybean (Glycine max) and its role in N2 fixation and yield was investigated via composite and whole transgenic plants. GmPT7 protein was localized to the plasma membrane and showed transport activity for Pi in yeast. Altered expression of GmPT7 changed (33) Pi uptake from rhizosphere and translocation to bacteroids. GmPT7 was mainly localized to the outer cortex and fixation zones of the nodules. Overexpression of GmPT7 promoted nodulation, and increased plant biomass, shoot nitrogen and phosphorus content, resulting in improved soybean yield by up to 36%. Double suppression of GmPT5 and GmPT7 led to nearly complete elimination of nodulation and over 50% reduction in plant biomass, shoot nitrogen and phosphorus content, indicating that both GmPT7 and GmPT5 contribute to Pi transport for N2 fixation. Taken together, our results indicate that GmPT7 is a transporter responsible for direct Pi entry to nodules and further to fixation zones, which is required for enhancing symbiotic N2 fixation and grain yield of soybean.
URLPMID:29769571 [本文引用: 1]
DOI:10.1104/pp.113.215400URLPMID:23569109 [本文引用: 1]
Heterotrimeric G proteins comprising Galpha, Gbeta, and Ggamma subunits regulate many fundamental growth and development processes in all eukaryotes. Plants possess a relatively limited number of G-protein components compared with mammalian systems, and their detailed functional characterization has been performed mostly in Arabidopsis (Arabidopsis thaliana) and rice (Oryza sativa). However, the presence of single Galpha and Gbeta proteins in both these species has significantly undermined the complexity and specificity of response regulation in plant G-protein signaling. There is ample pharmacological evidence for the role of G proteins in regulation of legume-specific processes such as nodulation, but the lack of genetic data from a leguminous species has restricted its direct assessment. Our recent identification and characterization of an elaborate G-protein family in soybean (Glycine max) and the availability of appropriate molecular-genetic resources have allowed us to directly evaluate the role of G-protein subunits during nodulation. We demonstrate that all G-protein genes are expressed in nodules and exhibit significant changes in their expression in response to Bradyrhizobium japonicum infection and in representative supernodulating and nonnodulating soybean mutants. RNA interference suppression and overexpression of specific G-protein components results in lower and higher nodule numbers, respectively, validating their roles as positive regulators of nodule formation. Our data further show preferential usage of distinct G-protein subunits in the presence of an additional signal during nodulation. Interestingly, the Galpha proteins directly interact with the soybean nodulation factor receptors NFR1alpha and NFR1beta, suggesting that the plant G proteins may couple with receptors other than the canonical heptahelical receptors common in metazoans to modulate signaling.
DOI:10.1105/tpc.15.00517URLPMID:26498905 [本文引用: 2]
Signaling pathways mediated by heterotrimeric G-protein complexes comprising Galpha, Gbeta, and Ggamma subunits and their regulatory RGS (Regulator of G-protein Signaling) protein are conserved in all eukaryotes. We have shown that the specific Gbeta and Ggamma proteins of a soybean (Glycine max) heterotrimeric G-protein complex are involved in regulation of nodulation. We now demonstrate the role of Nod factor receptor 1 (NFR1)-mediated phosphorylation in regulation of the G-protein cycle during nodulation in soybean. We also show that during nodulation, the G-protein cycle is regulated by the activity of RGS proteins. Lower or higher expression of RGS proteins results in fewer or more nodules, respectively. NFR1 interacts with RGS proteins and phosphorylates them. Analysis of phosphorylated RGS protein identifies specific amino acids that, when phosphorylated, result in significantly higher GTPase accelerating activity. These data point to phosphorylation-based regulation of G-protein signaling during nodule development. We propose that active NFR1 receptors phosphorylate and activate RGS proteins, which help maintain the Galpha proteins in their inactive, trimeric conformation, resulting in successful nodule development. Alternatively, RGS proteins might also have a direct role in regulating nodulation because overexpression of their phospho-mimic version leads to partial restoration of nodule formation in nod49 mutants.
[本文引用: 1]
DOI:10.1111/nph.14632URLPMID:28598036 [本文引用: 1]
Auxins play important roles in the nodulation of legumes. However, the mechanism by which auxin signaling regulates root nodulation is largely unknown. In particular, the role of auxin receptors and their regulation in determinate nodule development remains elusive. We checked the expression pattern of the auxin receptor GmTIR1/GmAFB3 genes in soybean. We analyzed the functions of GmTIR1/AFB3 in the regulation of rhizobial infection and nodule number, and also tested the functions of miR393 during nodulation and its relationship with GmTIR1/AFB3. The results showed that GmTIR1 and GmAFB3 genes exhibit diverse expression patterns during nodulation and overexpression of GmTIR1 genes significantly increased inflection foci and eventual nodule number. GmTIR1/AFB3 genes were post-transcriptionally cleaved by miR393 family and knock-down of the miR393 family members significantly increased rhizobial infection and the nodule number. Overexpression of the mutated form of GmTIR1C at the miR393 cleavage site that is resistant to miR393 cleavage led to a further increase in the number of infection foci and nodules, suggesting that miR393s modulate nodulation by directly targeting GmTIR1C. This study demonstrated that GmTIR1- and GmAFB3-mediated auxin signaling, that is spatio-temporally regulated by miR393, plays a crucial role in determinate nodule development in soybean.
DOI:10.1071/FP13123URLPMID:32481189 [本文引用: 1]
Auxin action is mediated by a complex signalling pathway involving transcription factors of the auxin response factor (ARF) family. In Arabidopsis, microRNA160 (miR160) negatively regulates three ARF genes (ARF10/ARF16/ARF17) and therefore controls several developmental processes, including primary and lateral root growth. Here, we analysed the role of miR160 in root development and nodulation in Medicago truncatula Gaertn. Bioinformatic analyses identified two main mtr-miR160 variants (mtr-miR160abde and mtr-miR160c) and 17 predicted ARF targets. The miR160-dependent cleavage of four predicted targets in roots was confirmed by analysis of parallel analysis of RNA ends (PARE) data and RACE-PCR experiments. Promoter-GUS analyses for mtr-miR160d and mtr-miR160c genes revealed overlapping but distinct expression profiles during root and nodule development. In addition, the early miR160 activation in roots during symbiotic interaction was not observed in mutants of the nodulation signalling or autoregulation pathways. Composite plants that overexpressed mtr-miR160a under two different promoters exhibited distinct defects in root growth and nodulation: the p35S:miR160a construct led to reduced root length associated to a severe disorganisation of the RAM, whereas pCsVMV:miR160a roots showed gravitropism defects and lower nodule numbers. Our results suggest that a regulatory loop involving miR160/ARFs governs root and nodule organogenesis in M. truncatula.
DOI:10.1104/pp.15.00265URLPMID:25941314 [本文引用: 1]
Legume root nodules convert atmospheric nitrogen gas into ammonium through symbiosis with a prokaryotic microsymbiont broadly called rhizobia. Auxin signaling is required for determinant nodule development; however, the molecular mechanism of auxin-mediated nodule formation remains largely unknown. Here, we show in soybean (Glycine max) that the microRNA miR167 acts as a positive regulator of lateral root organs, namely nodules and lateral roots. miR167c expression was up-regulated in the vasculature, pericycle, and cortex of soybean roots following inoculation with Bradyrhizobium japonicum strain USDA110 (the microsymbiont). It was found to positively regulate nodule numbers directly by repressing the target genes GmARF8a and GmARF8b (homologous genes of Arabidopsis [Arabidopsis thaliana] AtARF8 that encode auxin response factors). Moreover, the expression of miR167 and its targets was up- and down-regulated by auxin, respectively. The miR167-GmARF8 module also positively regulated nodulation efficiency under low microsymbiont density, a condition often associated with environmental stress. The regulatory role of miR167 on nodule initiation was dependent on the Nod factor receptor GmNFR1alpha, and it acts upstream of the nodulation-associated genes nodule inception, nodulation signaling pathway1, early nodulin40-1, NF-YA1 (previously known as HAEM activator protein2-1), and NF-YA2. miR167 also promoted lateral root numbers. Collectively, our findings establish a key role for the miR167-GmARF8 module in auxin-mediated nodule and lateral root formation in soybean.
DOI:10.1016/j.tplants.2012.04.006URLPMID:22658680 [本文引用: 2]
Plants need to acquire nitrogen (N) efficiently from the soil for growth. Nitrate is one of the major N sources for higher plants. Therefore, nitrate uptake and allocation are key factors in efficient N utilization. Membrane-bound transporters are required for nitrate uptake from the soil and for the inter- and intracellular movement of nitrate inside the plants. Four gene families, nitrate transporter 1/peptide transporter (NRT1/PTR), NRT2, chloride channel (CLC), and slow anion channel-associated 1 homolog 3 (SLAC1/SLAH), are involved in nitrate uptake, allocation, and storage in higher plants. Recent studies of these transporters or channels have provided new insights into the molecular mechanisms of nitrate uptake and allocation. Interestingly, several of these transporters also play versatile roles in nitrate sensing, plant development, pathogen defense, and/or stress response.
DOI:10.1105/tpc.11.8.1381URLPMID:10449574 [本文引用: 2]
The Arabidopsis CHL1 (AtNRT1) gene encodes an inducible component of low-affinity nitrate uptake, which necessitates a
[本文引用: 2]
[本文引用: 2]
DOI:10.1016/j.bbrc.2008.11.140URLPMID:19084500 [本文引用: 1]
miRNAs are approximately 21nt non-coding RNAs and play important roles in plant development and response to stress. Symbiotic nitrogen fixation (SNF) is agronomically important for reducing the need of nitrogen fertilizers. The soybean root nodule is the place where SNF takes place. To identify miRNAs that are possibly involved in nitrogen fixation in soybean functional nitrogen-fixing nodules, a small library of RNAs was constructed from the functional nodules harvested 28 days after inoculation with rhizobium. Thirty-two small RNA sequences were identified as belonging to 11 miRNA families. Eight miRNAs are conserved across plant species, twenty are specific to soybean, and the four remaining miRNAs are novel. Expression analysis revealed that miRNAs were differentially expressed in the different tissues. Combinatorial miRNA target prediction identified genes that are involved in multiple biological processes. The results suggest that miRNAs play critical and diverse roles in SNF, nutrient acquisition, and plant development.
URLPMID:17446894 [本文引用: 1]
DOI:10.1186/1471-2229-9-78URLPMID:19555486 [本文引用: 1]
BACKGROUND: Transgenic approaches provide a powerful tool for gene function investigations in plants. However, some legumes are still recalcitrant to current transformation technologies, limiting the extent to which functional genomic studies can be performed on. Superroot of Lotus corniculatus is a continuous root cloning system allowing direct somatic embryogenesis and mass regeneration of plants. Recently, a technique to obtain transgenic L. corniculatus plants from Superroot-derived leaves through A. tumefaciens-mediated transformation was described. However, transformation efficiency was low and it took about six months from gene transfer to PCR identification. RESULTS: In the present study, we developed an A. rhizogenes-mediated transformation of Superroot-derived L. corniculatus for gene function investigation, combining the efficient A. rhizogenes-mediated transformation and the rapid regeneration system of Superroot. The transformation system using A. rhizogenes K599 harbouring pGFPGUSPlus was improved by validating some parameters which may influence the transformation frequency. Using stem sections with one node as explants, a 2-day pre-culture of explants, infection with K599 at OD(600) = 0.6, and co-cultivation on medium (pH 5.4) at 22 degrees C for 2 days enhanced the transformation frequency significantly. As proof of concept, Superroot-derived L. corniculatus was transformed with a gene from wheat encoding an Na+/H+ antiporter (TaNHX2) using the described system. Transgenic Superroot plants were obtained and had increased salt tolerance, as expected from the expression of TaNHX2. CONCLUSION: A rapid and efficient tool for gene function investigation in L. corniculatus was developed, combining the simplicity and high efficiency of the Superroot regeneration system and the availability of A. rhizogenes-mediated transformation. This system was improved by validating some parameters influencing the transformation frequency, which could reach 92% based on GUS detection. The combination of the highly efficient transformation and the regeneration system of Superroot provides a valuable tool for functional genomics studies in L. corniculatus.
URLPMID:12668777 [本文引用: 1]
DOI:10.1270/jsbbs.56.295URL [本文引用: 1]
DOI:10.1038/ng.3337URLPMID:26053497 [本文引用: 1]
Asian cultivated rice (Oryza sativa L.) consists of two main subspecies, indica and japonica. Indica has higher nitrate-absorption activity than japonica, but the molecular mechanisms underlying that activity remain elusive. Here we show that variation in a nitrate-transporter gene, NRT1.1B (OsNPF6.5), may contribute to this divergence in nitrate use. Phylogenetic analysis revealed that NRT1.1B diverges between indica and japonica. NRT1.1B-indica variation was associated with enhanced nitrate uptake and root-to-shoot transport and upregulated expression of nitrate-responsive genes. The selection signature of NRT1.1B-indica suggests that nitrate-use divergence occurred during rice domestication. Notably, field tests with near-isogenic and transgenic lines confirmed that the japonica variety carrying the NRT1.1B-indica allele had significantly improved grain yield and nitrogen-use efficiency (NUE) compared to the variety without that allele. Our results show that variation in NRT1.1B largely explains nitrate-use divergence between indica and japonica and that NRT1.1B-indica can potentially improve the NUE of japonica.
DOI:10.1038/s41477-019-0384-1URLPMID:30911122 [本文引用: 1]
To ensure high crop yields in a sustainable manner, a comprehensive understanding of the control of nutrient acquisition is required. In particular, the signalling networks controlling the coordinated utilization of the two most highly demanded mineral nutrients, nitrogen and phosphorus, are of utmost importance. Here, we reveal a mechanism by which nitrate activates both phosphate and nitrate utilization in rice (Oryza sativa L.). We show that the nitrate sensor NRT1.1B interacts with a phosphate signalling repressor SPX4. Nitrate perception strengthens the NRT1.1B-SPX4 interaction and promotes the ubiquitination and degradation of SPX4 by recruiting NRT1.1B interacting protein 1 (NBIP1), an E3 ubiquitin ligase. This in turn allows the key transcription factor of phosphate signalling, PHR2, to translocate to the nucleus and initiate the transcription of phosphorus utilization genes. Interestingly, the central transcription factor of nitrate signalling, NLP3, is also under the control of SPX4. Thus, nitrate-triggered degradation of SPX4 activates both phosphate- and nitrate-responsive genes, implementing the coordinated utilization of nitrogen and phosphorus.
DOI:10.1038/s41587-019-0104-4URLPMID:31036930 [本文引用: 1]
Nitrogen-use efficiency of indica varieties of rice is superior to that of japonica varieties. We apply 16S ribosomal RNA gene profiling to characterize root microbiota of 68 indica and 27 japonica varieties grown in the field. We find that indica and japonica recruit distinct root microbiota. Notably, indica-enriched bacterial taxa are more diverse, and contain more genera with nitrogen metabolism functions, than japonica-enriched taxa. Using genetic approaches, we provide evidence that NRT1.1B, a rice nitrate transporter and sensor, is associated with the recruitment of a large proportion of indica-enriched bacteria. Metagenomic sequencing reveals that the ammonification process is less abundant in the root microbiome of the nrt1.1b mutant. We isolated 1,079 pure bacterial isolates from indica and japonica roots and derived synthetic communities (SynComs). Inoculation of IR24, an indica variety, with an indica-enriched SynCom improved rice growth in organic nitrogen conditions compared with a japonica-enriched SynCom. The links between plant genotype and root microbiota membership established in this study will inform breeding strategies to improve nitrogen use in crops.
URLPMID:8755617 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}