 ,1,*, 袁玺垒1, 李剑峰1, 王永芳2, 张小梅1, 张博1, 全建章2, 董志平,2,*
,1,*, 袁玺垒1, 李剑峰1, 王永芳2, 张小梅1, 张博1, 全建章2, 董志平,2,*Photo-thermal interaction model under different photoperiod-temperature conditions and expression analysis of SiCCT gene in foxtail millet (Setaria italica L.)
JIA Xiao-Ping,1,*, YUAN Xi-Lei1, LI Jian-Feng1, WANG Yong-Fang2, ZHANG Xiao-Mei1, ZHANG Bo1, QUAN Jian-Zhang2, DONG Zhi-Ping,2,*通讯作者:
收稿日期:2019-09-27接受日期:2020-01-15网络出版日期:2020-07-12
| 基金资助: |
Received:2019-09-27Accepted:2020-01-15Online:2020-07-12
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (1313KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
贾小平, 袁玺垒, 李剑峰, 王永芳, 张小梅, 张博, 全建章, 董志平. 不同光温条件谷子光温互作模式研究及SiCCT基因表达分析[J]. 作物学报, 2020, 46(7): 1052-1062. doi:10.3724/SP.J.1006.2020.94144
JIA Xiao-Ping, YUAN Xi-Lei, LI Jian-Feng, WANG Yong-Fang, ZHANG Xiao-Mei, ZHANG Bo, QUAN Jian-Zhang, DONG Zhi-Ping.
光周期和温度是决定植物开花时间的重要环境因素, 对农作物的生态适应性和产量有重要影响[1]。光周期和温度存在复杂的互作效应, 如拟南芥在长日照(16 h)诱导条件下温度从23℃升高到27℃能够有效促进开花[2]; 大麦在长日照条件下25℃比15℃培养能达到更高级的生殖发育期, 而在短日照条件下结果则相反[3]; 长日照加高温是最不利于开花诱导的光温组合, 长日照条件下, 高温抑制大豆的生殖发育, 低温减弱长日照对发育的抑制作用; 低温处理的大豆能正常出现花序, 而在高温处理下, 大多数植株一直处于营养生长期或部分发生花序的逆转, 表明高温增强了长日照对大豆生殖生长的抑制[4,5,6]。虽然目前通过对拟南芥、水稻的研究, 光周期调控植物开花的分子机制已经较为清晰[7,8], 有关环境温度调控植物开花分子机制的报道也逐渐增多[9], 但主要集中在光周期或者温度单一因素对植物开花调控作用, 对光温互作分子机制的研究极少。目前的研究初步发现拟南芥CO蛋白在光温互作调控机制中发挥着重要作用, 短日照条件低温(16℃)增加了组蛋白H2A.Z在成花素FT位点的占领, 阻止了光敏色素互作因子4 (PIF4)与FT基因启动子的结合, 高温(27℃)则使H2A.Z与FT位点分离, 诱导CO蛋白在中午积累, 并与PIF4形成复合体, 结合FT启动子, 促进基因表达, 诱导开花; 而长日照条件低温使短营养期蛋白(SVP)与FT启动子结合抑制其表达, 温度改变使SVP在黄昏时活性增加, 同时也增加了夜晚CO的稳定性, 从而使FT基因表达模式改变[10]。
CO作为光周期调控途径的关键基因, 属于CCT结构域基因家族, 该家族分为CMF、COL和PRR三个亚家族, 其中COL亚家族含有B-box锌指结构域和一个CCT结构域, PRR亚家族含有PRR结构域和一个CCT结构域, 而CMF亚家族只有一个CCT结构域[11]。研究表明许多CCT结构域基因都直接或者间接参与植物开花调控, 是光信号途径中的重要组成部分[12,13]。水稻中首先被克隆的CCT域基因Hd1是拟南芥中CO的同源基因, 长日照(LD)处理可以推迟水稻抽穗开花, 而短日照(SD)处理促进水稻抽穗开花[14]; LD、SD条件下, Hd1节律性都在夜间达到一个表达峰值, 该基因还可以通过影响每穗的小穗数(粒数)来控制水稻产量[15]。大麦Hd1的同源基因HvCO1不管日照长短, 其过表达都上调了HvFT1的表达, 使大麦提前开花[16]。高粱CO同源基因SbCO是COL亚家族一个成员, 在缺失SbGHD7和SbPRR37的背景下无论长日照还是短日照均能促进植株开花[17]。继Hd1之后在水稻中相继克隆了Ghd7、DTH2、OsCOL10、Ghd7.1 (OsPRR37)和Ghd2等含有CCT结构域的基因, 这些基因多数参与光周期调控的水稻开花过程, 但是具体功能有差异, 在LD条件下Ghd7、Ghd7.1通过抑制Ehd1和Hd3a的表达延迟水稻抽穗期[18,19], 与Ghd7、Ghd7.1功能类似, 高粱SbGhd7、玉米ZmCCT9在LD条件下同样能够推迟开花[20,21]; ZmCCT基因启动子中的一个CACTA转座子元件可以抑制ZmCCT的表达, 从而降低光周期敏感性, 使玉米在LD条件下得以传播种植[22]。DTH2则是通过上调成花素基因Hd3a和RFT1的表达来促进水稻提前开花[23]; OsCOL10在LD和SD条件下均抑制水稻开花, 还受到Ghd7的调控[24]; Ghd2参与对叶片衰老和干旱的调节[25]。从以上研究概述可以发现, CCT结构域基因普遍参与植物光周期开花调控, 但是这些基因是否参与温度调控的开花过程则极少报道, 特别是在C4作物中, 有关CCT结构域基因参与光温互作调控开花的研究尚属空白。
谷子(Setaria italica)属于禾本科狗尾草属, 是自花授粉的二倍体(2n=2x=18)作物, 具有抗旱、耐瘠和高光效等优良性状, 基因组较小(约515 Mb), 且全基因组序列已被测定[26,27], 可以作为C4禾谷类理想的模式作物[28,29]。作为一个短日照喜温作物, 谷子对光温较敏感, 前期的研究表明不同光周期环境对谷子株高、叶片数及穗部性状均有显著影响[30], 然而自然环境中除了光周期外, 温度的变化对作物生长发育也有作用, 光周期和温度往往存在复杂的互作效应, 因此通过研究光温互作对谷子等C4作物生长、发育及产量的影响, 为深入揭示以谷子为代表的C4禾谷类作物光温互作调控开花机制奠定基础, 具有重要的实践和理论意义。本研究设置长日照低温、长日照高温、短日照低温、短日照高温4个不同光温处理研究了光温互作对谷子抽穗期、叶片数、穗长、穗码数4个性状的影响, 明确谷子的光温互作模式, 进一步利用同源克隆技术从谷子中分离获得一个CCT域基因的完整编码序列, 在对其进行生物信息学分析、组织特异性表达分析的基础上, 详细分析4个不同光温组合条件下该基因的表达模式, 揭示该基因参与谷子光温调控开花的可能机制, 为进一步深入阐明谷子光温互作调控开花机制奠定基础。
1 材料与方法
1.1 试验材料
选用对光周期较敏感的河北省谷子农家品种‘黄毛谷’。1.2 谷子材料种植及表型性状调查
采用盆栽方法, 于口径10 cm×10 cm的小方盆中装营养土, 每盆播8~10粒种子, 二叶期定苗, 每盆保留4株长势一致的小苗。8盆‘黄毛谷’从播种至三叶期均在自然条件下生长, 随后移入光照培养箱进行长日照(16 h光照/8 h黑暗)处理, 4盆在处理1周后采集幼嫩叶片用于总RNA的提取及SiCCT基因克隆, 另外4盆继续培养至抽穗用于基因组织特异性表达分析, 处理期间温度为25℃。120盆‘黄毛谷’从播种至三叶期均在12 h光照/12 h黑暗、25℃的条件下培养, 三叶期开始将40盆‘黄毛谷’转入长日照(15 h光照/9 h黑暗)、短日照(9 h光照/15 h黑暗) 2个培养箱, 每个处理20盆, 2次重复, 每个重复10盆, 2个光周期处理温度均设定为25℃, 用于SiCCT基因长、短日照条件的表达分析; 80盆移入短日照高温[SD (9 h光照/15 h黑暗), 27℃]、短日照低温[SD (16 h光照/8 h黑暗), 22℃]、长日照高温[LD (16 h光照/8 h黑暗), 27℃]、长日照低温[LD (16 h光照/8 h黑暗), 22℃] 4个培养箱, 每个处理20盆, 2次重复, 每个重复10盆, 调查抽穗期、株高、穗长及叶片数, 用于光温互作模式分析及SiCCT基因不同光温处理表达分析。1.3 谷子SiCCT基因的克隆及生物信息学分析
首先利用生物信息学方法从NCBI的GenBank数据库中检索到一条谷子CCT域基因mRNA序列(XM_004983603), 根据此序列利用软件Primer Premier 5.0设计1对特异引物(SiCCT-F: 5'-TGGG AGGAGGAAGAGAGGAG-3', SiCCT-R: 5'-CTTCTT CAGGTCCGTGGCTA-3'), 扩增范围包含基因的完整编码区域, 预期大小为1160 bp。用康为世纪的Ultrapure RNA Kit提取总RNA, 用TaKaRa公司的PrimeScript II 1st Strand cDNA Synthesis Kit反转录合成第1链cDNA, 以合成的cDNA为模板, 用特异引物扩增目的基因, 扩增体系含cDNA模板2 μL、2× GC buffer 10 μL、10 mmol L-1 dNTPs 0.4 μL、2 μmol L-1特异引物4 μL、Taq DNA聚合酶0.2 μL, 以ddH2O 补充至20 μL。PCR 扩增程序为95℃预变性3 min; 95℃变性30 s, 58℃退火30 s, 72℃延伸45 s, 共35个循环; 72℃延伸10 min, 4℃保存。将RT-PCR扩增产物纯化后连接到pBM16A克隆载体上, 再转化大肠杆菌DH5α感受态细胞, 挑选阳性菌液送生工生物工程(上海)股份有限公司测序。利用ProtParam、ExPASy、SOPMA软件分析SiCCT基因的理化性质、二级和三级结构以及保守结构域。利用DNAMAN5.0软件将SiCCT基因编码的氨基酸序列与NCBI下载的其他物种CCT域基因氨基酸序列进行多序列比对, 使用MEGA 7.0软件构建谷子与几个物种的系统进化树。
1.4 谷子SiCCT基因的表达分析
将自然条件下生长至三叶期的‘黄毛谷’幼苗转入长日照条件培养, 取营养生长期的叶片、根、茎节、叶鞘、茎结和生殖生长期的幼穗、穗颈作为试验材料, 每个样品取3个重复, 用剪刀迅速剪下放入2 mL离心管, 以液氮速冻, 用于SiCCT基因组织特异性表达分析; 25℃恒温条件下, 长日照、短日照处理的谷苗均从三叶期开始取样, 长日照取到十叶期结束, 短日照取到八叶期开始抽穗, 抽穗后7 d、15 d取样结束, 期间还对长至五至六叶期的谷苗, 从晚6时黑暗处理开始48 h内取样, 每隔3 h取一次, 每个时间点重复取样3次, 取完全展开的顶叶和第2叶片, 用于SiCCT基因长、短日照条件表达分析; 4个不同光温处理的谷苗均从三叶期开始取样直到抽穗, 取完全展开的顶叶和第2片叶, 取样时间为光照3 h后, 每个叶龄取1次样, 3个重复, 样品以液氮速冻, 用于SiCCT基因4个光温处理的表达分析。所有样品RNA提取及反转录过程同实验步骤1.3。根据SiCCT基因cDNA序列的编码区来设计实时荧光定量PCR引物, Q-F为5'-CGTGGACCCGAC GATGGAC-3'; Q-R为5'-GTCAGGCACCTTGGCG AATC-3'。参考文献用谷子Actin基因作为内参, 引物为SiActin-F: 5'-GGCAAACAGGGAGAAGATGA-3'和SiActin-R: 5'-GAGGTTGTCGGTAAGGTCACG-3', 由北京鼎国昌盛生物技术有限责任公司合成[31]。
将反转录好的cDNA作为Real-time PCR的模板, 用引物Q-F/Q-R扩增SiCCT基因的特异保守片段, 同时以引物SiActin-F和SiActin-R扩增作内参, 每个样品做3个重复, 扩增体系及循环程序参照宝生物公司TB Green Premix Ex Taq II (Tli RNaseH Plus)的试剂说明书, 首先配制10 μL扩增体系, 包含cDNA模板1 μL、TB Green Premix Ex Taq II (Tli RNaseH Plus) 5 μL、10 μmol L-1的Q-F和Q-R特异引物各1 μL、ddH2O 2 μL。使用Roche Light Cycler 96实时定量PCR仪, 采用两步法PCR程序, 扩增程序为95℃预变性30 s; 95℃变性30 s, 60℃退火30 s, 共40个循环。分析得到扩增曲线、溶解曲线, 并计算??CT值, 采用2-ΔΔCT计算方法进行相对表达量分析。
2 结果与分析
2.1 谷子光温互作模式
光周期对谷子的生长发育起关键作用, 高温条件下, 长日照处理的‘黄毛谷’比短日照处理抽穗时间推迟了60.3 d, 株高增加了30.4 cm, 叶片数增加了7.7片, 穗长增加了7.6 cm, 长日照相比短日照营养生长期明显延长; 低温条件下, 长日照处理的‘黄毛谷’抽穗时间比短日照处理推迟了21.4 d, 株高增加7.5 cm, 叶片数几乎一样, 穗长增加了4.7 cm, 说明温度的变化并不能改变长日照比短日照延长‘黄毛谷’营养生长期的总体趋势(图1和图2-a~d)。同时光周期也决定着温度的作用方向, 短日照条件下, 高温处理的‘黄毛谷’与低温处理相比株高减少了11.2 cm, 叶片数减少了2.0片, 抽穗期提前了11.3 d, 穗长减少了1.2 cm, 说明短日照条件下, 提高温度可以使‘黄毛谷’快速完成营养生长, 进入生殖生长期; 长日照条件下, 高温处理的‘黄毛谷’抽穗时间比低温处理延长了30 d, 株高增加了18.8 cm, 叶片数增加了5.6片, 穗长减少了2.1 cm, 说明长日照条件下, 提高温度使‘黄毛谷’营养生长期延长, 推迟进入生殖生长期, 长、短日照条件下温度的作用方向正好相反(图1, 图2-a~d)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同光温处理对‘黄毛谷’生长发育的影响
SD: 短日照; LD: 长日照。
Fig. 1Effect of different photo-thermal treatments on growth and development of ‘Huangmaogu’
SD: short day; LD: long day.
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同光温处理间‘黄毛谷’4个性状的比较
a: 抽穗期; b: 株高; c: 穗长; d: 叶片数。SD: 短日照; LD: 长日照。
Fig. 2Comparison of four traits among different photo-thermal treatments
a: heading date; b: plant height; c: panicle length; d: leaf number. SD: short day; LD: long day.
短日照高温处理的谷子抽穗期最短, 缩短营养生长期的作用最强, 其次是短日照低温处理; 长日照低温、长日照高温2个处理的谷子抽穗期均比短日照低温、高温2个处理延长, 且长日照高温组合抽穗期最晚(图2-a)。
2.2 谷子SiCCT基因的克隆及生物信息学分析
提取长日照处理的‘黄毛谷’叶片总RNA, 经1%非变性琼脂糖凝胶电泳检测看到, 28S rRNA和18S rRNA条带清晰, 且前者的亮度大约是后者的1~2倍(图3)。经过超微量分光光度计检测, OD260/280均在1.97~2.10之间, 说明总RNA完整性比较好, 基本没有降解, 可以进行后续实验。将提取的总RNA反转录得到cDNA, 以此为模板使用基因特异性引物SiCCT-F、SiCCT-R进行RT-PCR扩增, 扩增产物经1%琼脂糖凝胶电泳检测得到一条清晰、明亮的特异性条带, 大小在1000~2000 bp之间, 与预期目的片段1160 bp相接近(图4)。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3‘黄毛谷’叶片总RNA电泳图
1, 2: 提取的两管RNA。
Fig. 3Electrophoregram of total RNA extracted from leaves of ‘Huangmaogu’
1, 2: two tubes of RNA extracted.
图4
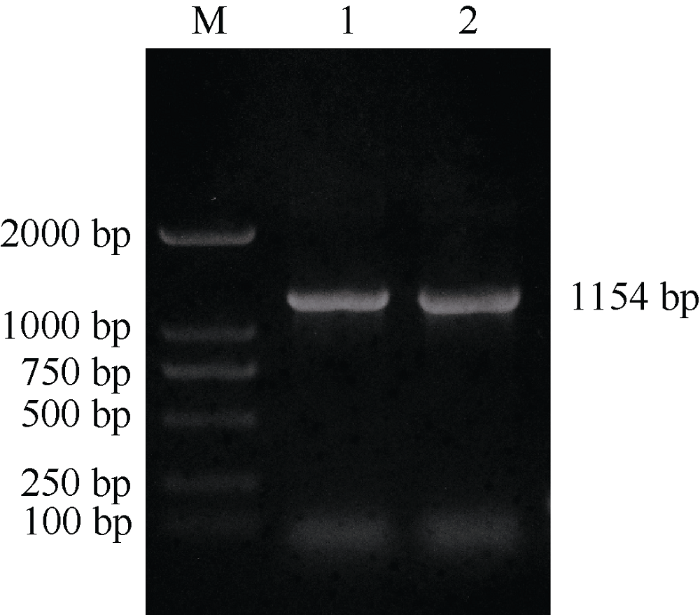 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4 SiCCT基因RT-PCR产物电泳图
M: marker DL2000; 1, 2: 两管RT-PCR产物。
Fig. 4Electrophoregram of RT-PCR products of SiCCT gene
M: marker DL2000; 1, 2: two tubes of RT-PCR products.
将RT-PCR扩增产物经纯化后连接到pBM16A克隆载体上, 再转化大肠杆菌DH5α感受态细胞,挑选阳性菌液送往生工生物工程(上海)股份有限公司进行测序, 获得大小为1154 bp的cDNA序列, 该序列的CDS全长861 bp, 编码286个氨基酸(附图1, 附图2)。分析SiCCT蛋白的结构域发现在215~257 aa之间存在1个保守的CCT结构域(附图3), 因此本研究所克隆的SiCCT基因属于CMF亚家族。
附图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附图1SiCCT基因cDNA序列
粗体部分为引物序列,下画线部分为起始密码子和终止密码子。
Supplementary Fig. 1cDNA sequences of SiCCT gene
The bolded parts are primer sequences, the underlined parts are initiation codon and termination codon.
附图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附图2SiCCT基因编码的氨基酸序列
Supplementary Fig. 2The deduced amino acid sequence of SiCCT gene
附图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附图3SiCCT基因编码蛋白质的保守结构域预测分析
Supplementary Fig. 3Prediction for the conserved domains of SiCCT protein
用SiCCT蛋白序列同源搜索NCBI数据库, 共检索到9条与SiCCT蛋白存在直系同源关系的序列, 这些序列来自水稻(XM_015757733.2)、高粱(XM_ 002464888.2)、玉米(XM_008660049.2)、节节麦(XP_ 020151451.1)、二穗短柄草(XM_003574283.3)、海枣树(XP_008796123.1)、油棕(XP_010933637)、野生糜(XP_025825012)、菠萝(XP_020093545) 9个物种。基于包括SiCCT蛋白在内的10个CCT结构域蛋白序列构建系统进化树, 10个物种可以聚为3个组, 谷子、高粱、玉米、水稻、节节麦、二穗短柄草聚在一个组, 亲缘关系较近, 它们同属于禾本科; 油棕、海枣树、菠萝聚为一组; 野生糜单独聚为一组(图5)。
图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5基于CCT域基因蛋白序列的系统发育树
Fig. 5Phylogenetic tree of CCT-motif genes based on protein sequences
2.3 谷子SiCCT基因的组织特异性表达分析
从图6可以看出, SiCCT基因在根、茎节、叶、茎结、叶鞘、穗和穗颈7个不同组织中都有表达, 但是在叶片中的相对表达量最高, 其次是穗和叶鞘, 在根部的相对表达量最低。图6
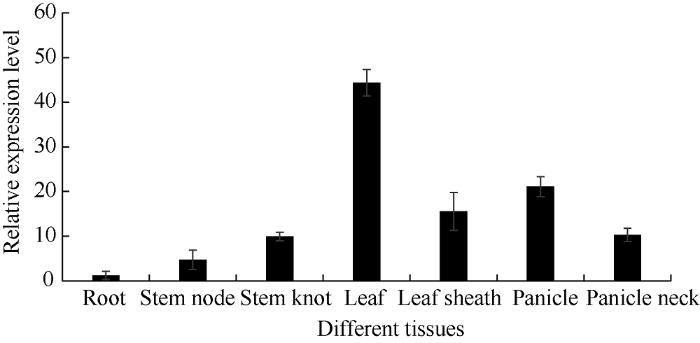 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6SiCCT基因在不同组织中的相对表达量
Fig. 6Relative expression of SiCCT in different tissues
2.4 不同光周期条件谷子SiCCT基因的昼夜表达分析
短日照条件下连续取样48 h, 以研究SiCCT基因在连续2 d中表达的变化情况, 发现SiCCT基因在‘黄毛谷’叶片中呈现24 h节律性表达, 黑暗条件0~3 h几乎不表达, 6 h之后表达量逐渐增高, 黑暗第12 h表达达到一个小峰, 随后下降, 在光照开始3 h后达到一个峰值, 然后迅速下降, 在24 h光照结束后表达量最低; 24 h之后的下一个循环周期同样SiCCT基因在黑暗6 h (30 h)的时候表达量开始缓慢上升, 在黑暗12 h (36 h)时达到一个峰值, 然后缓慢下降, 见光后开始上升, 当光照3 h (42 h)后再一次达到高峰, 随后又逐渐下降, 在光照快结束时(45 h)达到一个较低的水平。SiCCT基因的表达在1 d之内有2个峰, 不过白天表达峰值差不多是晚上峰值的2倍左右(图7-a)。图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7不同光周期条件SiCCT基因的昼夜表达
a: 短日照处理; b: 长日照处理。黑色条带表示黑暗时段, 白色条带表示光照时段。
Fig. 7Circadian expression of SiCCT gene under different photoperiod conditions
a: short-day treatment; b: long-day treatment. Black bars represent dark period, and white bars represent light period.
在长日照条件下, ‘黄毛谷’叶片中SiCCT基因的相对表达也呈现出24 h节律性, 黑暗表达量低, 光照表达量高, 光照开始后表达量迅速增加, 光照3 h后达到一个较高峰值, 随后快速下降, 光照结束后表达降到极低水平; 下一个循环周期与前面24 h表达模式相似, 同样开始光照后3 h表达达到峰值(图7-b)。与在短日照条件不同的是长日照条件 SiCCT基因只出现一个表达峰, 且是在光照3 h后出现, 而短日照条件出现2个表达峰, 一个是在黑暗12 h, 一个是在光照3 h。
2.5 不同光周期条件、不同生长期谷子SiCCT基因的表达分析
在短日照条件下, 从三叶期到七叶期‘黄毛谷’叶片中SiCCT基因表达随叶龄的增加而增加, 七叶期达峰值。八叶期谷子开始抽穗, SiCCT基因的相对表达量明显降低, 抽穗后7 d和抽穗后15 d SiCCT基因的表达也处于较低水平, 说明在谷子营养生长期SiCCT基因表达水平高, 而抽穗后的生殖生长期表达量始终较低(图8-a)。在长日照条件下, ‘黄毛谷’叶片中SiCCT基因从三叶期到十叶期相对表达量均随叶龄增加而增加, 八叶期达峰值, 九叶期、十叶期略下降, 但仍维持较高水平。因为长日照导致营养生长期延长, 十叶期谷子依然处于营养生长期, 没有抽穗, 说明无论短日照还是长日照, SiCCT基因在营养生长期表达量较高(图8-b)。图8
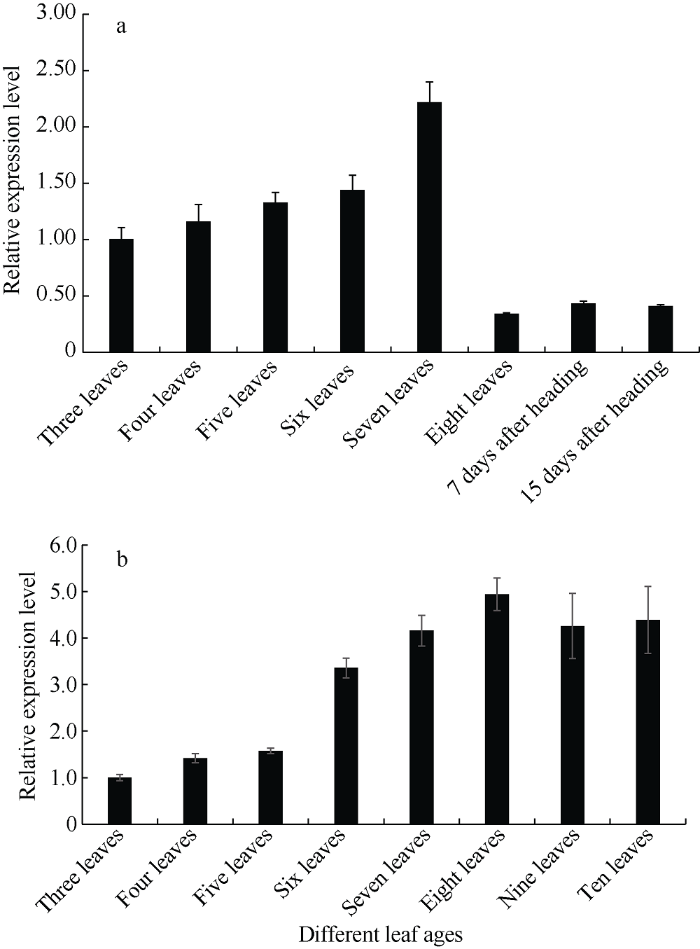 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8不同光周期条件SiCCT 基因在不同叶龄的表达水平
a: 短日照; b: 长日照。
Fig. 8Expression level of SiCCT gene at different leaf ages under different photoperiod conditions
a: short-day; b: long-day.
2.6 不同光温组合条件谷子SiCCT基因的表达分析
在高温条件下, 三到五叶期长日照处理的‘黄毛谷’叶片中SiCCT基因的表达总体低于短日照处理, 从六叶期开始, 长日照处理的SiCCT基因的表达水平明显高于短日照处理(图9-a); 在低温条件下, 三叶期到十叶期短日照处理的‘黄毛谷’叶片中SiCCT基因的表达水平均低于长日照处理(图9-b)。高温短日照处理‘黄毛谷’八叶期已抽穗, 而低温短日照处理‘黄毛谷’十叶期仍处于营养生长阶段, 说明短日照条件下低温对‘黄毛谷’营养生长具有延长作用; 在短日照条件下, 三叶期到八叶期营养生长阶段总体上低温处理SiCCT基因的表达水平高于高温处理, 只有五叶期高温处理SiCCT基因表达水平高于低温处理, 而八叶期后高温处理的‘黄毛谷’开始抽穗, 进入生殖生长期, SiCCT基因表达水平非常低, 低温处理的‘黄毛谷’仍处于营养生长期, 九叶期、十叶期SiCCT基因的表达水平仍比较高(图9-c); 长日照条件下, 除了三叶期高温处理和低温处理SiCCT基因表达水平接近外, 四叶期到十叶期高温处理‘黄毛谷’叶片中SiCCT基因的表达水平均高于低温处理, 与短日照条件表达模式相反(图9-d)。图9
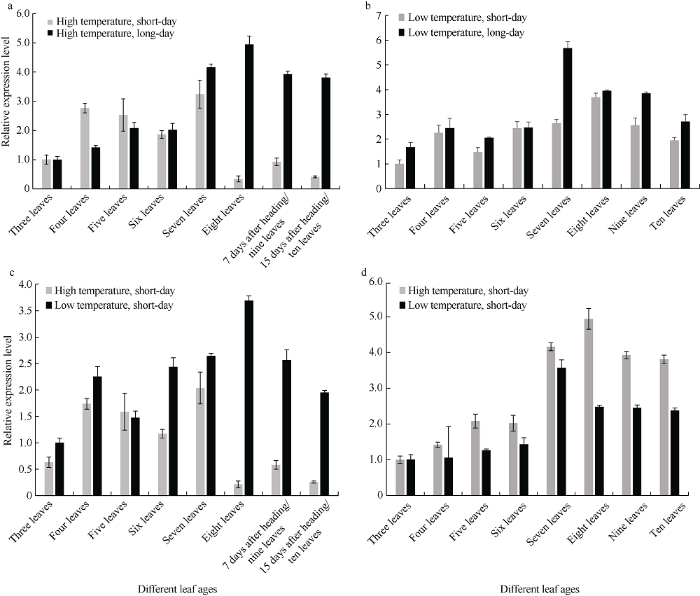 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9不同光温组合SiCCT基因的表达特点
a: 高温长日照、高温短日照; b: 低温长日照、低温短日照; c: 短日照高温、短日照低温; d: 长日照高温、长日照低温。
Fig. 9Expression feature of SiCCT gene under different photo-thermal combinations
a: high temperature and long-day, high temperature and short-day; b: low temperature and long-day, low temperature and short-day; c: short-day and high temperature, short-day and low temperature; d: long-day and high temperature, long-day and low temperature.
3 讨论
光周期和温度是影响植物开花的2个重要环境因子, 而且它们对植物生长发育存在明显的互作效应。本研究发现不论高温还是低温, 长日照条件下的‘黄毛谷’比短日照都晚抽穗, 在短日照条件下, 高温促进‘黄毛谷’抽穗并开花, 低温使‘黄毛谷’持续进行营养生长而延迟抽穗; 长日照条件下, 高温却延迟了‘黄毛谷’的抽穗, 植株长期处于营养生长阶段, 而且大多只抽穗很少开花, 结籽很少, 低温能促进‘黄毛谷’进入生殖生长阶段, 提前抽穗并开花。研究结果表明短日照和高温、长日照和低温对谷子的生殖生长都具有促进作用, 而高温加剧了长日照对谷子生殖生长的抑制作用, 低温减轻了长日照对谷子生殖生长的抑制作用, 谷子和大豆具有相似的光温互作模式[5,6]。目前在水稻、玉米、高粱和甘蓝型油菜中已经克隆的CMF亚家族成员有Ghd7、NRR、OsCCT01、SbGHD7、ZmCCT9等基因, 这些基因多数受光周期调控, 对作物抽穗期开花期有重要影响[16,22,25,32-33], 而谷子CMF亚家族成员还未有报道, 其与水稻等作物受光周期调控的模式是否相同仍是未知。本研究首次从谷子中分离到一个CMF亚家族成员, 命名为SiCCT, 组织特异性表达分析表明该基因在叶片中表达量最高, 其次是幼穗, 与水稻Ghd7基因具有相似的组织表达特异性[16], 而多数受光信号调控的基因在叶片中的相对表达量较高, 因此推测谷子SiCCT基因受光周期调控。进一步分析发现SiCCT基因无论短日照还是长日照, 均表现昼夜节律性表达, 短日照条件光照期和黑暗期各有一个表达峰, 前者远高于后者; 长日照条件只有光照产生一个表达峰。水稻Ghd7基因无论长日照还是短日照仅在白天有一个表达峰[16], 高粱Ghd7的同源基因SbGHD7在长日照条件下有2个表达峰, 分别在清晨和夜晚产生[25], 说明谷子SiCCT基因与Ghd7、SbGHD7昼夜表达模式存在差异。然而谷子SiCCT基因与Ghd7、SbGHD7、ZmCCT9也具有相似的表达模式, 即无论长日照还是短日照, 整个营养生长期表达量都较高, 短日照条件谷子抽穗及抽穗后SiCCT基因表达量降到极低水平, 而长日照条件一直维持营养生长, SiCCT基因表达量维持在较高水平。因此推测SiCCT基因与Ghd7、SbGHD7、ZmCCT9具有相似的功能, 在长日照条件下高水平的表达抑制抽穗, 延长营养生长期。
本研究充分说明SiCCT基因和许多CCT域基因一样参与了光周期调控开花途径[34,35], 而光周期和温度对谷子生长发育有明显的互作效应, 因此SiCCT基因是否也受温度调控, 参与光周期途径和感温性途径的交互作用, 是需要弄清楚的下一个问题。通过对拟南芥光温互作机制研究发现, CO基因在光周期途径和感温性途径交互作用中非常关键, 短日照条件低温(16℃)增加了组蛋白H2A.Z在成花素FT位点的占领, 阻止了光敏色素互作因子4 (PIF4)与FT基因启动子的结合, 高温(27℃)则使H2A.Z与FT位点分离, 诱导CO蛋白在中午积累, 并与PIF4形成复合体, 结合FT启动子, 促进基因表达, 诱导开花; 而长日照条件低温使短营养期蛋白(SVP)与FT启动子结合抑制其表达, 温度增加使SVP在黄昏时活性增加, 同时也增加了夜晚CO的稳定性, 从而使FT基因表达模式改变[10]。本研究发现SiCCT基因在受光周期调控的同时也受温度调节, 长日照条件下, 低温处理的‘黄毛谷’叶片中SiCCT基因的相对表达量明显低于高温处理, 说明在长日照条件下高温促进SiCCT基因的表达, 而低温抑制SiCCT基因的表达, 这与水稻长日照条件下低温(23℃)处理促进Ghd7表达的结果相反[36]。而本研究发现高温增加了长日照对‘黄毛谷’生殖发育的抑制作用, 低温减轻了长日照对‘黄毛谷’生殖发育的抑制作用, 这与长日照条件下高温SiCCT基因表达量明显高于低温是相对应的, 更能说明SiCCT基因表达水平与‘黄毛谷’营养生长期存在正相关。无论高温还是低温, 长日照条件下SiCCT基因在不同叶期整体表达水平都高于短日照, 说明SiCCT基因总体上受光周期调控的作用要大于温度。本研究证明了SiCCT基因同时受光周期和温度调控, 而该基因如何与感温性途径的基因相互作用, 并且如何共同作用于谷子成花素基因, 从而实现光温互作对谷子的开花调控, 是后续的主要研究目标。
4 结论
光周期对谷子的发育起关键作用, 不论高温还是低温, 长日照条件下谷子营养生长期要比短日照延长; 温度的作用随光周期的不同而不同, 在短日照条件下, 高温缩短谷子营养生长期而低温促使营养生长期延长, 长日照条件下则相反, 高温延长谷子营养生长期而低温缩短谷子营养生长期; 谷子SiCCT基因在叶片中高表达, 长短日照均表现昼夜节律性表达特点, 整个营养生长期表达量相对较高; SiCCT基因主要受光周期调控, 同时也受温度调控, 其表达量与谷子营养生长期具有明显的正相关, 推测SiCCT基因通过参与光周期和温度互作调控机制来调节谷子抽穗期。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/j.tplants.2013.05.003URLPMID:23790253 [本文引用: 1]

Plants monitor changes in photoperiod and temperature to synchronize their flowering with seasonal changes to maximize fitness. In the Arabidopsis photoperiodic flowering pathway, the circadian clock-regulated components, such as FLAVIN-BINDING, KELCH REPEAT, F-BOX 1 and CONSTANS, both of which have light-controlled functions, are crucial to induce the day-length specific expression of the FLOWERING LOCUS T (FT) gene in leaves. Recent advances indicate that FT transcriptional regulation is central for integrating the information derived from other important internal and external factors, such as developmental age, amount of gibberellic acid, and the ambient temperature. In this review, we describe how these factors interactively regulate the expression of FT, the main component of florigen, in leaves.
URLPMID:16839183 [本文引用: 1]
The transition to flowering is an important event in the plant life cycle and is modulated by several environmental factors including photoperiod, light quality, vernalization, and growth temperature, as well as biotic and abiotic stresses. In contrast to light and vernalization, little is known about the pathways that mediate the responses to other environmental variables. A mild increase in growth temperature, from 23 degrees C to 27 degrees C, is equally efficient in inducing flowering of Arabidopsis plants grown in 8-h short days as is transfer to 16-h long days. There is extensive natural variation in this response, and we identify strains with contrasting thermal reaction norms. Exploiting this natural variation, we show that FLOWERING LOCUS C potently suppresses thermal induction, and that the closely related floral repressor FLOWERING LOCUS M is a major-effect quantitative trait locus modulating thermosensitivity. Thermal induction does not require the photoperiod effector CONSTANS, acts upstream of the floral integrator FLOWERING LOCUS T, and depends on the hormone gibberellin. Analysis of mutants defective in salicylic acid biosynthesis suggests that thermal induction is independent of previously identified stress-signaling pathways. Microarray analyses confirm that the genomic responses to floral induction by photoperiod and temperature differ. Furthermore, we report that gene products that participate in RNA splicing are specifically affected by thermal induction. Above a critical threshold, even small changes in temperature can act as cues for the induction of flowering. This response has a genetic basis that is distinct from the known genetic pathways of floral transition, and appears to correlate with changes in RNA processing.
DOI:10.1104/pp.111.192013URL [本文引用: 1]
High temperature influences plant development and can reduce crop yields. We examined how ambient temperature influences reproductive development in the temperate cereals wheat (Triticum aestivum) and barley (Hordeum vulgare). High temperature resulted in rapid progression through reproductive development in long days, but inhibited early stages of reproductive development in short days. Activation of the long-day flowering response pathway through day-lengthinsensitive alleles of the PHOTOPERIOD1 gene, which result in high FLOWERING LOCUS T-like1 transcript levels, did not allow rapid early reproductive development at high temperature in short days. Furthermore, high temperature did not increase transcript levels of FLOWERING LOCUS T-like genes. These data suggest that genes or pathways other than the longday response pathway mediate developmental responses to high temperature in cereals. Transcriptome analyses suggested a possible role for vernalization-responsive genes in the developmental response to high temperature. The MADS-box floral repressor HvODDSOC2 is expressed at elevated levels at high temperature in short days, and might contribute to the inhibition of early reproductive development under these conditions. FLOWERING PROMOTING FACTOR1-like, RNase-S-like genes, and VER2-like genes were also identified as candidates for high-temperature-responsive developmental regulators. Overall, these data suggest that rising temperatures might elicit different developmental responses in cereal crops at different latitudes or times of year, due to the interaction between temperature and day length. Additionally, we suggest that different developmental regulators might mediate the response to high temperature in cereals compared to Arabidopsis (Arabidopsis thaliana).
DOI:10.2135/cropsci2001.413721xURL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1093/jxb/eru416URLPMID:25326628 [本文引用: 1]
The timing of flowering is a crucial decision in the life cycle of plants since favourable conditions are needed to maximize reproductive success and, hence, the survival of the species. It is therefore not surprising that plants constantly monitor endogenous and environmental signals, such as day length (photoperiod) and temperature, to adjust the timing of the floral transition. Temperature in particular has been shown to have a tremendous effect on the timing of flowering: the effect of prolonged periods of cold, called the vernalization response, has been extensively studied and the underlying epigenetic mechanisms are reasonably well understood in Arabidopsis thaliana. In contrast, the effect of moderate changes in ambient growth temperature on the progression of flowering, the thermosensory pathway, is only starting to be understood on the molecular level. Several genes and molecular mechanisms underlying the thermosensory pathway have already been identified and characterized in detail. At a time when global temperature is rising due to climate change, this knowledge will be pivotal to ensure crop production in the future.
DOI:10.1111/nph.13883URLPMID:26856528 [本文引用: 2]
Day length and ambient temperature are major stimuli controlling flowering time. To understand flowering mechanisms in more natural conditions, we explored the effect of daily light and temperature changes on Arabidopsis thaliana. Seedlings were exposed to different day/night temperature and day-length treatments to assess expression changes in flowering genes. Cooler temperature treatments increased CONSTANS (CO) transcript levels at night. Night-time CO induction was diminished in flowering bhlh (fbh)-quadruple mutants. FLOWERING LOCUS T (FT) transcript levels were reduced at dusk, but increased at the end of cooler nights. The dusk suppression, which was alleviated in short vegetative phase (svp) mutants, occurred particularly in younger seedlings, whereas the increase during the night continued over 2 wk. Cooler temperature treatments altered the levels of FLOWERING LOCUS M-beta (FLM-beta) and FLM-delta splice variants. FT levels correlated strongly with flowering time across treatments. Day/night temperature changes modulate photoperiodic flowering by changing FT accumulation patterns. Cooler night-time temperatures enhance FLOWERING BHLH (FBH)-dependent induction of CO and consequently increase CO protein. When plants are young, cooler temperatures suppress FT at dusk through SHORT VEGETATIVE PHASE (SVP) function, perhaps to suppress precocious flowering. Our results suggest day length and diurnal temperature changes combine to modulate FT and flowering time.
DOI:10.1093/jxb/erm042URLPMID:17420173 [本文引用: 1]
The control of flowering is central to reproductive success in plants, and has a major impact on grain yield in crop species. The global importance of temperate cereal crops such as wheat and barley has meant emphasis has long been placed on understanding the genetics of flowering in order to enhance yield. Leads gained from the dissection of the molecular genetics of model species have combined with comparative genetic approaches, recently resulting in the isolation of the first flowering time genes in wheat and barley. This paper reviews the genetics and genes involved in cereal flowering pathways and the current understanding of how two of the principal genes, Vrn and Ppd, have been involved in domestication and adaptation to local environments, and the implications for future breeding programmes are discussed.
DOI:10.1105/tpc.104.029504URLPMID:15705949 [本文引用: 1]
Environmental time cues, such as photocycles (light/dark) and thermocycles (warm/cold), synchronize (entrain) endogenous biological clocks to local time. Although much is known about entrainment of the Arabidopsis thaliana clock to photocycles, the determinants of thermoperception and entrainment to thermocycles are not known. The Arabidopsis PSEUDO-RESPONSE REGULATOR (PRR) genes, including the clock component TIMING OF CAB EXPRESSION 1/PRR1, are related to bacterial, fungal, and plant response regulators but lack the conserved Asp that is normally phosphorylated by an upstream sensory kinase. Here, we show that two PRR family members, PRR7 and PRR9, are partially redundant; single prr7-3 or prr9-1 mutants exhibit modest period lengthening, but the prr7-3 prr9-1 double mutant shows dramatic and more than additive period lengthening in the light and becomes arrhythmic in constant darkness. The prr7-3 prr9-1 mutant fails both to maintain an oscillation after entrainment to thermocycles and to reset its clock in response to cold pulses and thus represents an important mutant strongly affected in temperature entrainment in higher plants. We conclude that PRR7 and PRR9 are critical components of a temperature-sensitive circadian system. PRR7 and PRR9 could function in temperature and light input pathways or they could represent elements of an oscillator necessary for the clock to respond to temperature signals.
DOI:1000-2421(2010)06-0669-08URL [本文引用: 1]
通过分析4个完成测序和注释的植物基因组,系统地分离鉴定了97个水稻、玉米、高粱和拟南芥的CCT结构域基因,并对相应蛋白质的结构和基因之间的系统演化关系进行了分析。结果表明:CCT结构域基因的蛋白质结构和特性在不同物种之间具有广泛的变异;不同基因组中CCT结构域基因通过染色体复制扩展了基因家族成员。根据其CCT结构域分为4组,其中1个亚组集中了绝大部分的禾本科CCT结构域基因,该亚组基因可能特异参与了禾本科作物开花时间的调控。基因家族成员的扩展,蛋白质结构的改变以及基因表达模式的变异共同导致了CCT结构域基因家族成员在物种间和物种内的功能分化。
DOI:1000-2421(2010)06-0669-08URL [本文引用: 1]
通过分析4个完成测序和注释的植物基因组,系统地分离鉴定了97个水稻、玉米、高粱和拟南芥的CCT结构域基因,并对相应蛋白质的结构和基因之间的系统演化关系进行了分析。结果表明:CCT结构域基因的蛋白质结构和特性在不同物种之间具有广泛的变异;不同基因组中CCT结构域基因通过染色体复制扩展了基因家族成员。根据其CCT结构域分为4组,其中1个亚组集中了绝大部分的禾本科CCT结构域基因,该亚组基因可能特异参与了禾本科作物开花时间的调控。基因家族成员的扩展,蛋白质结构的改变以及基因表达模式的变异共同导致了CCT结构域基因家族成员在物种间和物种内的功能分化。
DOI:10.1105/tpc.12.12.2473URLPMID:11148291 [本文引用: 1]
A major quantitative trait locus (QTL) controlling response to photoperiod, Hd1, was identified by means of a map-based cloning strategy. High-resolution mapping using 1505 segregants enabled us to define a genomic region of approximately 12 kb as a candidate for Hd1. Further analysis revealed that the Hd1 QTL corresponds to a gene that is a homolog of CONSTANS in Arabidopsis. Sequencing analysis revealed a 43-bp deletion in the first exon of the photoperiod sensitivity 1 (se1) mutant HS66 and a 433-bp insertion in the intron in mutant HS110. Se1 is allelic to the Hd1 QTL, as determined by analysis of two se1 mutants, HS66 and HS110. Genetic complementation analysis proved the function of the candidate gene. The amount of Hd1 mRNA was not greatly affected by a change in length of the photoperiod. We suggest that Hd1 functions in the promotion of heading under short-day conditions and in inhibition under long-day conditions.
DOI:10.1038/nature01549URLPMID:12700762 [本文引用: 1]
The photoperiodic control of flowering is one of the important developmental processes of plants because it is directly related to successful reproduction. Although the molecular genetic analysis of Arabidopsis thaliana, a long-day (LD) plant, has provided models to explain the control of flowering time in this species, very little is known about its molecular mechanisms for short-day (SD) plants. Here we show how the photoperiodic control of flowering is regulated in rice, a SD plant. Overexpression of OsGI, an orthologue of the Arabidopsis GIGANTEA (GI) gene in transgenic rice, caused late flowering under both SD and LD conditions. Expression of the rice orthologue of the Arabidopsis CONSTANS (CO) gene was increased in the transgenic rice, whereas expression of the rice orthologue of FLOWERING LOCUS T (FT) was suppressed. Our results indicate that three key regulatory genes for the photoperiodic control of flowering are conserved between Arabidopsis, a LD plant, and rice, a SD plant, but regulation of the FT gene by CO was reversed, resulting in the suppression of flowering in rice under LD conditions.
DOI:10.1111/j.1365-313X.2011.04839.xURL [本文引用: 4]
Variation in photoperiod response is a major factor determining plant development and the agronomic performance of crops. The genetic control of photoperiodic flowering has been elucidated in the model plant Arabidopsis, and many of the identified genes are structurally conserved in the grasses. In this study, HvCO1, the closest barley ortholog of the key photoperiod response gene CONSTANS in Arabidopsis, was over-expressed in the spring barley Golden Promise. Over-expression of HvCO1 accelerated time to flowering in long- and short-day conditions and caused up-regulation of HvFT1 mRNA under long-day conditions. However, the transgenic plants retained a response to photoperiod, suggesting the presence of photoperiod response factors acting downstream of HvCO1 transcription. Analysis of a population segregating for HvCO1 over-expression and natural genetic variation at Ppd-H1 demonstrated that Ppd-H1 acts downstream of HvCO1 transcription on HvFT1 expression and flowering. Furthermore, variation at Ppd-H1 did not affect diurnal expression of HvCO1 or HvCO2. Over-expression of HvCO1 increased transcription of the spring allele of Vrn-H1 in long- and short-day conditions, while genetic variation at Ppd-H1 did not affect Vrn-H1 expression. Over-expression of HvCO1 and natural genetic variation at Ppd-H1 accelerated inflorescence development and stem elongation. Thus, HvCO1 probably induces flowering by activating HvFT1 whilst Ppd-H1 regulates HvFT1 independently of HvCO1 mRNA, and all three genes also appear to have a strong effect in promoting inflorescence development.
DOI:10.1186/1471-2229-14-1URLPMID:24387633 [本文引用: 1]
BACKGROUND: The characteristics of pollen tube growth are not constant, but display distinct patterns of growth within the different tissues of the pistil. In the stigma, the growth rate is slow and autotrophic, whereas in the style, it is rapid and heterotrophic. Very little is known about the interactions between these distinct maternal tissues and the traversing pollen tube and the role of this interaction on the observed metabolism. In this work we characterise pollen tube growth in the apple flower and look for differences in glycoprotein epitope localization between two different maternal tissues, the stigma and the style. RESULTS: While immunocytochemically-detected arabinogalactan proteins were present at high levels in the stigma, they were not detected in the transmitting tissue of the style, where extensins were abundant. Whereas extensins remained at high levels in unpollinated pistils, they were no longer present in the style following pollen tube passage. Similarily, while abundant in unpollinated styles, insoluble polysaccharides such as beta-glucans, were depleted in pollinated pistils. CONCLUSIONS: The switch from autotropic to heterotrophic pollen tube growth correlates spatially with a change of glycoprotein epitopes between the stigma and the style. The depletion of extensins and polysaccharides following pollen tube passage in the style suggest a possible contribution to the acceleration of heterotrophic pollen tube growth, which would imply an active contribution of female tissues on prezygotic male-female crosstalk.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1073/pnas.1718058115URLPMID:29279404 [本文引用: 1]
From its tropical origin in southwestern Mexico, maize spread over a wide latitudinal cline in the Americas. This feat defies the rule that crops are inhibited from spreading easily across latitudes. How the widespread latitudinal adaptation of maize was accomplished is largely unknown. Through positional cloning and association mapping, we resolved a flowering-time quantitative trait locus to a Harbinger-like transposable element positioned 57 kb upstream of a CCT transcription factor (ZmCCT9). The Harbinger-like element acts in cis to repress ZmCCT9 expression to promote flowering under long days. Knockout of ZmCCT9 by CRISPR/Cas9 causes early flowering under long days. ZmCCT9 is diurnally regulated and negatively regulates the expression of the florigen ZCN8, thereby resulting in late flowering under long days. Population genetics analyses revealed that the Harbinger-like transposon insertion at ZmCCT9 and the CACTA-like transposon insertion at another CCT paralog, ZmCCT10, arose sequentially following domestication and were targeted by selection for maize adaptation to higher latitudes. Our findings help explain how the dynamic maize genome with abundant transposon activity enabled maize to adapt over 90 degrees of latitude during the pre-Columbian era.
DOI:10.1007/s00122-017-2978-1URLPMID:28916922 [本文引用: 2]
KEY MESSAGE: The elite ZmCCT haplotypes which have no transposable element in the promoter could enhance maize resistance to Gibberella stalk rot and improve yield-related traits, while having no or mild impact on flowering time. Therefore, they are expected to have great value in future maize breeding programs. A CCT domain-containing gene, ZmCCT, is involved in both photoperiod response and stalk rot resistance in maize. At least 15 haplotypes are present at the ZmCCT locus in maize germplasm, whereas only three of them are found in Chinese commercial maize hybrids. Here, we evaluated ZmCCT haplotypes for their potential application in corn breeding. Nine resistant ZmCCT haplotypes that have no CACTA-like transposable element in the promoter were introduced into seven elite maize inbred lines by marker-assisted backcrossing. The resultant 63 converted lines had 0.7-5.1 Mb of resistant ZmCCT donor segments with over 90% recovery rates. All converted lines tested exhibited enhanced resistance to maize stalk rot but varied in photoperiod sensitivity. There was a close correlation between the hybrids and their parental lines with respect to both resistance performance and photoperiod sensitivity. Furthermore, in a given hybrid A5302/83B28, resistant ZmCCT haplotype could largely improve yield-related traits, such as ear length and 100-kernel weight, resulting in enhanced grain yield. Of nine resistant ZmCCT haplotypes, haplotype H5 exhibited excellent performance for both flowering time and stalk rot resistance and is thus expected to have potential value in future maize breeding programs.
DOI:10.1073/pnas.1213962110URLPMID:23388640 [本文引用: 1]
Flowering time (i.e., heading date in crops) is an important ecological trait that determines growing seasons and regional adaptability of plants to specific natural environments. Rice (Oryza sativa L.) is a short-day plant that originated in the tropics. Increasing evidence suggests that the northward expansion of cultivated rice was accompanied by human selection of the heading date under noninductive long-day (LD) conditions. We report here the molecular cloning and characterization of DTH2 (for Days to heading on chromosome 2), a minor-effect quantitative trait locus that promotes heading under LD conditions. We show that DTH2 encodes a CONSTANS-like protein that promotes heading by inducing the florigen genes Heading date 3a and RICE FLOWERING LOCUS T 1, and it acts independently of the known floral integrators Heading date 1 and Early heading date 1. Moreover, association analysis and transgenic experiments identified two functional nucleotide polymorphisms in DTH2 that correlated with early heading and increased reproductive fitness under natural LD conditions in northern Asia. Our combined population genetics and network analyses suggest that DTH2 likely represents a target of human selection for adaptation to LD conditions during rice domestication and/or improvement, demonstrating an important role of minor-effect quantitative trait loci in crop adaptation and breeding.
[本文引用: 1]
[本文引用: 1]
DOI:10.1093/jxb/erw344URLPMID:27638689 [本文引用: 3]
CONSTANS (CO)-like genes have been intensively investigated for their roles in the regulation of photoperiodic flowering, but very limited information has been reported on their functions in other biological processes. Here, we found that a CO-like gene, Ghd2 (Grain number, plant height, and heading date2), which can increase the yield potential under normal growth condition just like its homologue Ghd7, is involved in the regulation of leaf senescence and drought resistance. Ghd2 is expressed mainly in the rice (Oryza sativa) leaf with the highest level detected at the grain-filling stage, and it is down-regulated by drought stress conditions. Overexpression of Ghd2 resulted in significantly reduced drought resistance, while its knockout mutant showed the opposite phenotype. The earlier senescence symptoms and the transcript up-regulation of many senescence-associated genes (SAGs) in Ghd2-overexpressing transgenic rice plants under drought stress conditions indicate that Ghd2 plays essential roles in accelerating drought-induced leaf senescence in rice. Moreover, developmental and dark-induced leaf senescence was accelerated in the Ghd2-overexpressing rice and delayed in the ghd2 mutant. Several SAGs were confirmed to be regulated by Ghd2 using a transient expression system in rice protoplasts. Ghd2 interacted with several regulatory proteins, including OsARID3, OsPURalpha, and three 14-3-3 proteins. OsARID3 and OsPURalpha showed expression patterns similar to Ghd2 in rice leaves, with the highest levels at the grain-filling stage, whereas OsARID3 and the 14-3-3 genes responded differently to drought stress conditions. These results indicate that Ghd2 functions as a regulator by integrating environmental signals with the senescence process into a developmental programme through interaction with different proteins.
DOI:10.1038/nbt.2196URLPMID:22580951 [本文引用: 1]
We generated a high-quality reference genome sequence for foxtail millet (Setaria italica). The approximately 400-Mb assembly covers approximately 80% of the genome and >95% of the gene space. The assembly was anchored to a 992-locus genetic map and was annotated by comparison with >1.3 million expressed sequence tag reads. We produced more than 580 million RNA-Seq reads to facilitate expression analyses. We also sequenced Setaria viridis, the ancestral wild relative of S. italica, and identified regions of differential single-nucleotide polymorphism density, distribution of transposable elements, small RNA content, chromosomal rearrangement and segregation distortion. The genus Setaria includes natural and cultivated species that demonstrate a wide capacity for adaptation. The genetic basis of this adaptation was investigated by comparing five sequenced grass genomes. We also used the diploid Setaria genome to evaluate the ongoing genome assembly of a related polyploid, switchgrass (Panicum virgatum).
DOI:10.1038/nbt.2195URLPMID:22580950 [本文引用: 1]
Foxtail millet (Setaria italica), a member of the Poaceae grass family, is an important food and fodder crop in arid regions and has potential for use as a C(4) biofuel. It is a model system for other biofuel grasses, including switchgrass and pearl millet. We produced a draft genome ( approximately 423 Mb) anchored onto nine chromosomes and annotated 38,801 genes. Key chromosome reshuffling events were detected through collinearity identification between foxtail millet, rice and sorghum including two reshuffling events fusing rice chromosomes 7 and 9, 3 and 10 to foxtail millet chromosomes 2 and 9, respectively, that occurred after the divergence of foxtail millet and rice, and a single reshuffling event fusing rice chromosome 5 and 12 to foxtail millet chromosome 3 that occurred after the divergence of millet and sorghum. Rearrangements in the C(4) photosynthesis pathway were also identified.
DOI:10.1105/tpc.110.075309URLPMID:20693355 [本文引用: 1]
C(4) photosynthesis drives productivity in several major food crops and bioenergy grasses, including maize (Zea mays), sugarcane (Saccharum officinarum), sorghum (Sorghum bicolor), Miscanthus x giganteus, and switchgrass (Panicum virgatum). Gains in productivity associated with C(4) photosynthesis include improved water and nitrogen use efficiencies. Thus, engineering C(4) traits into C(3) crops is an attractive target for crop improvement. However, the lack of a small, rapid cycling genetic model system to study C(4) photosynthesis has limited progress in dissecting the regulatory networks underlying the C(4) syndrome. Setaria viridis is a member of the Panicoideae clade and is a close relative of several major feed, fuel, and bioenergy grasses. It is a true diploid with a relatively small genome of ~510 Mb. Its short stature, simple growth requirements, and rapid life cycle will greatly facilitate genetic studies of the C(4) grasses. Importantly, S. viridis uses an NADP-malic enzyme subtype C(4) photosynthetic system to fix carbon and therefore is a potentially powerful model system for dissecting C(4) photosynthesis. Here, we summarize some of the recent advances that promise greatly to accelerate the use of S. viridis as a genetic system. These include our recent successful efforts at regenerating plants from seed callus, establishing a transient transformation system, and developing stable transformation.
DOI:10.3109/07388551.2012.716809URL [本文引用: 1]
Foxtail millet is one of the oldest domesticated diploid C-4 Panicoid crops having a comparatively small genome size of approximately 515 Mb, short life cycle, and inbreeding nature. Its two species, Setaria italica (domesticated) and Setaria viridis (wild progenitor), have characteristics that classify them as excellent model systems to examine several aspects of architectural, evolutionary, and physiological importance in Panicoid grasses especially the biofuel crops such as switchgrass and napiergrass. Foxtail millet is a staple crop used extensively for food and fodder in parts of Asia and Africa. In its long history of cultivation, it has been adapted to arid and semi-arid areas of Asia, North Africa, South and North America. Foxtail millet has one of the largest collections of cultivated as well as wild-type germplasm rich with phenotypic variations and hence provides prospects for association mapping and allele-mining of elite and novel variants to be incorporated in crop improvement programs. Most of the foxtail millet accessions can be primarily abiotic stress tolerant particularly to drought and salinity, and therefore exploiting these agronomic traits can enhance its efficacy in marker-aided breeding as well as in genetic engineering for abiotic stress tolerance. In addition, the release of draft genome sequence of foxtail millet would be useful to the researchers worldwide in not only discerning the molecular basis of biomass production in biofuel crops and the methods to improve it, but also for the introgression of beneficial agronomically important characteristics in foxtail millet as well as in related Panicoid bioenergy grasses.
DOI:10.3724/SP.J.1006.2019.84128URL [本文引用: 1]
本研究连续2年在短日照(海南)、中等日照(河南)、长日照(吉林) 3个不同光周期环境调查160份谷子资源的抽穗期、株高、叶片数、穗长、穗粗、穗码数、码粒数、穗重、穗粒重、千粒重10个主要性状, 利用SPSS 19.0软件进行多因素方差分析及多重比较, 揭示光周期对谷子各性状的影响, 评价160份谷子资源对光周期的敏感性。结果表明, 3个不同光周期环境间谷子株高、叶片数、穗长、穗码数、抽穗期、穗粗、穗重、穗粒重、码粒数9个性状均表现出极显著差异(P<0.01), 千粒重表现出显著差异(P<0.05)。株高、叶片数、穗长、穗码数、抽穗期5个性状随着日照的延长呈现出递增趋势。品种对谷子的10个性状有极显著影响(P<0.01), 年份对千粒重以外的9个性状有极显著影响(P<0.01)。光周期与品种互作效应对10个性状均有极显著影响(P<0.01), 光周期与年份互作效应对抽穗期以外的9个性状有极显著影响(P<0.01), 年份与品种互作效应对株高、叶片数、穗长、穗粗、穗重、穗粒重、穗码数、码粒数8个性状有极显著影响(P<0.01)。160份谷子资源中对光周期表现中、低敏感性的材料没有明显的地域特性, 而对光周期表现强敏感性的材料主要是来自春谷区的农家品种。筛选出光周期极不敏感材料小早谷和极端敏感材料呼和浩特大毛谷、然谷、红钙谷、茄谷、二白谷等, 为选育谷子光周期钝感品种及开展光周期敏感性形成机理研究奠定了基础。
DOI:10.3724/SP.J.1006.2019.84128URL [本文引用: 1]
本研究连续2年在短日照(海南)、中等日照(河南)、长日照(吉林) 3个不同光周期环境调查160份谷子资源的抽穗期、株高、叶片数、穗长、穗粗、穗码数、码粒数、穗重、穗粒重、千粒重10个主要性状, 利用SPSS 19.0软件进行多因素方差分析及多重比较, 揭示光周期对谷子各性状的影响, 评价160份谷子资源对光周期的敏感性。结果表明, 3个不同光周期环境间谷子株高、叶片数、穗长、穗码数、抽穗期、穗粗、穗重、穗粒重、码粒数9个性状均表现出极显著差异(P<0.01), 千粒重表现出显著差异(P<0.05)。株高、叶片数、穗长、穗码数、抽穗期5个性状随着日照的延长呈现出递增趋势。品种对谷子的10个性状有极显著影响(P<0.01), 年份对千粒重以外的9个性状有极显著影响(P<0.01)。光周期与品种互作效应对10个性状均有极显著影响(P<0.01), 光周期与年份互作效应对抽穗期以外的9个性状有极显著影响(P<0.01), 年份与品种互作效应对株高、叶片数、穗长、穗粗、穗重、穗粒重、穗码数、码粒数8个性状有极显著影响(P<0.01)。160份谷子资源中对光周期表现中、低敏感性的材料没有明显的地域特性, 而对光周期表现强敏感性的材料主要是来自春谷区的农家品种。筛选出光周期极不敏感材料小早谷和极端敏感材料呼和浩特大毛谷、然谷、红钙谷、茄谷、二白谷等, 为选育谷子光周期钝感品种及开展光周期敏感性形成机理研究奠定了基础。
[本文引用: 1]
[本文引用: 1]
DOI:10.1093/mp/sss157URLPMID:23253602 [本文引用: 1]
DOI:10.1038/srep07663URLPMID:25563494 [本文引用: 1]
CCT domain-containing genes generally control flowering in plants. Currently, only six of the 41 CCT family genes have been confirmed to control flowering in rice. To efficiently identify more heading date-related genes from the CCT family, we compared the positions of heading date QTLs and CCT genes and found that 25 CCT family genes were located in the QTL regions. Association mapping showed that a total of 19 CCT family genes were associated with the heading date. Five of the seven associated genes within QTL regions and two of four associated genes outside of the QTL regions were confirmed to regulate heading date by transformation. None of the seven non-associated genes outside of the QTL regions regulates heading date. Obviously, combination of candidate gene-based association mapping with linkage analysis could improve the identification of functional genes. Three novel CCT family genes, including one non-associated (OsCCT01) and two associated genes (OsCCT11 and OsCCT19) regulated the heading date. The overexpression of OsCCT01 delayed flowering through suppressing the expression of Ehd1, Hd3a and RFT1 under both long day and short day conditions. Potential functions in regulating heading date of some untested CCT family genes were discussed.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}