 ,*, 赵晓曼, 徐珂, 王芯蕊, 张辰瑜, 东方阳河北科技师范学院农学与生物科技学院, 河北昌黎 066600
,*, 赵晓曼, 徐珂, 王芯蕊, 张辰瑜, 东方阳河北科技师范学院农学与生物科技学院, 河北昌黎 066600Anatomy and microscopic observation of Ricinus communis seed structure
GUO Xue-Min,*, ZHAO Xiao-Man, XU Ke, WANG Xin-Rui, ZHANG Chen-Yu, DONG-FANG YangCollege of Agronomy and Biotechnology, Heibei Normal University of Science & Technology, Changli 066600, Hebei, China通讯作者:
收稿日期:2019-09-23接受日期:2020-01-15网络出版日期:2020-06-12
| 基金资助: |
Received:2019-09-23Accepted:2020-01-15Online:2020-06-12
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (15210KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
郭学民, 赵晓曼, 徐珂, 王芯蕊, 张辰瑜, 东方阳. 蓖麻种子结构的解剖和显微观察[J]. 作物学报, 2020, 46(6): 914-923. doi:10.3724/SP.J.1006.2020.94141
GUO Xue-Min, ZHAO Xiao-Man, XU Ke, WANG Xin-Rui, ZHANG Chen-Yu, DONG-FANG Yang.
大戟科(Euphorbiaceae)的蓖麻(Ricinus communis L.)是重要的经济作物。蓖麻种子是一种植物学模式材料, 蓖麻油是全球特种化学工业的重要原料[1,2,3,4,5], 它是羟基化脂肪酸(hydroxylated fatty acid)的唯一商业来源。蓖麻种子是生物燃料的重要资源[6], 其提取物具有重要的抗病活性[7,8], 尤其是它所含蓖麻毒蛋白是最强的植物源毒素之一, 主要应用于生物农药领域和医药领域[9,10,11,12]。我国是蓖麻油的主要生产国和消费国, 加强蓖麻的基础研究, 对蓖麻资源的综合开发利用、提升国民经济水平具有十分重要的意义。
国外许多****观察了授粉后50 d的蓖麻种皮的结构[13]、种阜的结构[14]、萌发5 d的蓖麻种子子叶和胚乳的结构[15]、种子萌发后6~7 d胚乳从种皮侧到子叶侧超微结构的变化[16]以及在子叶衰老过程中叶绿体等细胞器的变化[17]。国内不少《植物学》教材中都有关于蓖麻种子的形态和胚乳细胞糊粉粒的扼要描述[18,19]。这些研究关注的是种子发育或萌发过程中某一时期某一部分的结构及其变化, 目前, 尚未见到完整成熟种子结构和维管束分布的报道。
本文利用石蜡切片法和显微照相技术, 较为系统地观察了成熟蓖麻种子的解剖学结构和维管束分布, 以期为从整体上认识种子的结构, 以及研究种子发育和萌发过程中物质运输的生理、生化和分子生物学提供基本资料。
1 材料与方法
1.1 材料
试验材料为从市面上购买的健康、饱满蓖麻种子。1.2 方法
1.2.1 种子形态观察 选择30粒左右成熟饱满、结构完整的蓖麻种子, 用蒸馏水浸泡12 h, 分别游离其中15粒种子各组成部分, 以观察其形态结构; 其余15粒在潮湿的培养皿中室温下进行萌发试验,以观察种孔的位置, 先后分别在OLYMPUS SZX16体视显微镜下观察、照相。1.2.2 种子解剖结构观察 取完整饱满成熟蓖麻种子10粒, 以蒸馏水浸泡12 h, 分离出外种皮、内种皮、种阜、胚乳和胚5个部分。
以FAA固定液分别固定种子背侧、侧面和腹侧外种皮, 24 h后, 以软化剂(过氧化氢:冰醋酸=1:2)软化2个月; 随后, 乙醇梯度脱水、二甲苯透明、浸蜡、换蜡、包埋、修块, 用ESM-100L大型滑走式切片机切片, 厚度14 μm, 经展片、烘片、脱蜡、染色、封片等, 制成永久切片。
同样, 以FAA固定内种皮、种阜、胚乳和胚, 经过上述步骤处理, 用KD-2258轮转切片机连续切片, 切片厚度8~12 μm, 制作切片。
分别用番红固绿对外种皮和内种皮染色, 以番红固绿与希夫试剂分别对种阜、胚乳、子叶和胚染色, 旨在显示不可溶性多糖(希夫试剂)和木质化、木栓化和角质化的细胞壁(番红固绿双重染色)。
在光学显微镜Olympus BX51下观察切片, 用DP72相机照相。选择代表性照片, 用Photoshop CS 6.0软件制作图版。
1.2.3 内种皮维管束分布的观察 选择成熟完整蓖麻种子, 去除外种皮, 将其余部分在蒸馏水中浸泡12 h, 从种阜端向上轻轻剥下相对完整的内种皮。用番红染色, 酸乙醇分色, 甘油封片, 在显微镜Olympus BX51下观察并照相记录。
2 结果与分析
2.1 种子形态
蓖麻种子呈扁卵形或扁椭圆形, 较平坦的一面为腹侧, 隆起一面为背侧, 它由种皮、胚乳和胚3部分构成(图1-A~E)。种子萌发时, 胚根从种阜基部、靠近种子背面的种孔突破种皮(图1-F), 种阜依然完整, 这表明种孔并未被种阜所覆盖。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1蓖麻种子的形态
A: 种子腹面观, 示外种皮的纹饰和种脊(箭头); B: 去外种皮的种子, 示内种皮维管束(箭头)的分布; C: 去种皮的种子, 示乳白色的胚乳; D: 与子叶平面垂直的正中纵切面, 示胚在种子中的位置; E: 游离的胚, 示子叶、胚轴和胚根; F: 去部分种皮、胚根突破种孔的萌发种子, 示种孔在种阜背侧, 并未被种阜覆盖。HI: 种脐; CA: 种阜; EN: 胚乳; TE: 种皮; EM: 胚; CO: 子叶; HY: 胚轴; RA: 胚根。Bar = 2 mm。
Fig. 1Shape of R. communis seed
A: ventral view of a seed, showing the ornamentation and raphe (arrow) of a seed; B: a seed without episperm, showing the distribution of vascular bundles (arrow) in endopleura; C: a seed without testa, showing the milky white endosperm; D: median longitudinal section perpendicular to the plane of cotyledon, showing the position of an embryo in a seed; E: a free embryo, showing cotyledon, hypocotyle and radicle; F: a germinating seed with partial testa and radicle breaking through the micropyle, showing the micropyle on the dorsal side of the caruncle and not covered by the caruncle. HI: Hilum; CA: Caruncle; EN: Endosperm; TE: Testa; EM: Embryo; CO: Cotyledon; HY: Hypocotyle; RA: Radicle. Bar = 2 mm.
2.2 种皮的解剖结构
蓖麻种皮包括外种皮(图2-A~E)、内种皮(图2- F~I; 图3)和种阜(图1-A; 图4-A~C)。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2蓖麻种皮的解剖结构
A: 种子背侧外种皮垂直于种子长轴横切面, 示长柱状表皮层、海绵组织层和栅栏组织层; B: 外种皮表皮细胞横切面, 示表皮细胞椭圆形形态; C: 种子侧面外种皮垂直于种子长轴横切面, 示长柱状表皮层、海绵组织层和栅栏组织层; D: 种子腹侧外种皮沿种脊延伸方向的横切面, 示柱状表皮层、海绵组织层和栅栏组织层以及位于海绵组织层的大型维管束; E: 种子腹侧外种皮垂直于种脊延伸方向的横切面, 示大型维管束; F: 内种皮分布示意图, 示在种阜端内种皮内层和外层围成的气室; G: 外种皮腹侧内侧体视显微镜图像, 示种阜端外种皮和内种皮外层不易分离, 以及种脊大型维管束在种阜相对一端与内种皮维管束索的连接点(箭头); H: 内种皮脉间区横切面, 示马氏层、扁平的海绵组织层和内珠被内层; I: 内种皮沿维管束延伸方向的横切面, 示维管束位于海绵组织层中。EP: 表皮层; SP: 海绵组织层; PP: 栅栏组织层; LB: 大型维管束; IE: 内种皮内层; OE: 内种皮外层; ML: 马氏层; IL: 内珠被内层; EB: 内种皮维管束。
Fig. 2Anatomical structure of R. communis testa
A: transection of episperm on the back of a seed perpendicular to the long axis, showing columnar epidermis, spongy parenchyma and palisade parenchyma; B: transection of epidermal cells from the episperm, showing epidermal cells in oval shape; C: transection of lateral episperm of a seed perpendicular to the long axis, showing columnar epidermis, spongy parenchyma and palisade parenchyma; D: transection of episperm at the ventral surface of a seed along the direction of raphe extension, showing columnar epidermis, spongy parenchyma, palisade parenchyma, and the large bundle in spongy parenchyma; E: transection of episperm at the ventral surface of a seed perpendicular to the direction of ridge extension, showing large vascular bundle; F: a schematic diagram of the distribution of endopleura, showing an air chamber surrounded by inner and outer layers of endopleura at caruncle end; G: stereomicroscopic image from the inner side of the ventral episperm, showing the nonseparable pattern of episperm and endopleura, and the junction point (arrow) between the large bundle of raphe and the bundle band of endopleura at the opposite end of the caruncle; H: transection of endopleura from the vein-islet, showing malpighian layer, flat spongy parenchyma and inner layer of inner integument; I: transection of endopleura along the direction of bundle extension, showing the vascular bundle located in spongy parenchyma. EP: epidermis; SP: spongy parenchyma; PP: palisade parenchyma; LB: large bundle; IE: inner layer of endopleura; OE: outer layer of endopleura; ML: malpighian layer; IL: inner layer of inner integument; EB: endopleura vascular bundle.
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3蓖麻内种皮维管束分布
A: 部分游离的内种皮, 示被染成红色的、代表维管束的木质部和维管束的分枝格局; B: 图A中方框a的放大图, 示与外种皮腹侧种脊大型维管束相连的维管束索, 分枝为3个较大的维管束; C: 图A中方框b的放大图, 示管胞末端侧壁相连(箭头); D: 图A中方框c的放大图, 示维管束分枝的格局; E: 图A中方框d的放大图, 示内种皮维管束末端的螺纹管胞(箭头)。BB: 维管束索。
Fig. 3Distribution of endopleura vascular bundles of R. communis
A: partial free endopleura, showing the reddish xylems representing vascular bundles and branching pattern of vascular bundles; B: detailed view of box a in figure A, showing that the vascular bundle band, connecting with large vascular bundle in the raphe from episperm at the ventral surface of a seed, branched into three larger vascular bundles; C: detailed view of box b in figure A, showing terminal side wall junction (arrow) of the tracheids; D: detailed view of box c in figure A, showing the pattern of branching of vascular bundles; E: detailed view of box d in figure A, showing the helical tracheid (arrow) at the end of vascular bundle of endopleura. BB: bundle band.
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4蓖麻种子种阜的形态与结构
A~C: 体视显微镜图片, 示种阜的形态; D~J: 横切面; K和L: 纵切面; D和H~J: 希夫试剂染色, 示细胞壁由糖类物质构成; E~G、K、L: 番红和固绿双重染色, 示细胞壁不角质层和含木质素。A: 种阜背面观, 示种阜中部的浅沟将种阜上部分为2个球形突起; B: 种阜腹面观, 示种阜和外种皮相连接; C: 种阜底面观, 示种阜管道在内种皮内层和外层之间的开口(箭头), 该开口靠近种子背侧, 以及白色的内种皮外层; D: 种阜上部横切面, 示2个球形突起; E: 种阜浅沟基部横切, 示未出现种阜管道; F: 种阜中部外侧横切面, 示外侧3层小型的细胞和内部大型的细胞; G: 种阜中部横切放大图, 示发达的纹孔(箭头); H: 种阜中部横切面, 示出现种阜管道; I: 图H中方框a放大图, 示Y字形种阜管道; J: 图H中方框b放大图, 示种阜中的维管束(箭头); K: 距种阜管道中轴一点距离的种阜纵切面, 示种阜管道(箭头)上端并未和浅沟基部联通; L: 沿种阜管道中轴的纵切面, 示种阜管道基部的开口(箭头)。EM: 外种皮; CC: 种阜管道。
Fig. 4Shape and structure of the caruncle of R. communis seed
A-C: stereomicrograph, showing the shape of the caruncle; D-J: transection; K and L: longitudinal section; D and H-J: images stained with Schiff’s reagent, showing cell walls composed of carbohydrates; E-G, K, L: images with double staining of saffron and fixed green, showing cell walls without lignin and cuticle. A: view on the back of the caruncle, showing the shallow ditch in the middle part of the caruncle, which divided its upper part into two spherical protrusions; B: ventral view of the caruncle, showing the tight combination of caruncle and episperm; C: view on the bottom of the caruncle, showing an opening (arrow) of caruncle channel between the inner and outer layers of the endopleura and near the dorsal surface of the seed, and the outer layer of the white endopleura; D: transection of the upper part of the caruncle, showing two spherical protrusions; E: transection of shallow ditch base of the caruncle, showing no caruncle channel; F: transection of the middle lateral side of the caruncle, showing three layers of small and cuticle-free cells on the outside and large parenchyma cells inside; G: detailed view of transection of the middle part of the caruncle, showing developed pits (arrow); H: transection of middle part of the caruncle, showing the caruncle channel; I: detailed view of box a in figure H, showing Y-shaped caruncle channel; J: detailed view of box b in figure H, showing vascular bundle in the caruncle; K: longitudinal section of the caruncle a short distance from the central axis of the caruncle channel, showing the upper end of the caruncle channel (arrow) which did not connected with the base of the shallow ditch; L: longitudinal section along the central axis of the caruncle channel, showing the opening (arrow) at the base of the caruncle channel. EM: episperm; CC: caruncle channel.
2.2.1 外种皮解剖结构 外种皮表面光滑, 有黑色和黄褐色的纹饰; 种脊位于种子腹侧中央, 与种子长轴平行, 并未向外突出, 它是倒生胚珠的珠柄与珠被愈合处留于外种皮上的痕迹(图1-A)。
种子各部分外种皮的结构基本相同, 由外至内分别为长柱状表皮层、海绵组织层和栅栏组织层(图2-A~E)。外种皮腹侧种脊海绵组织层贯穿有大型维管束(图2-D, E), 始于种脐, 止于种阜相对一端(相当于胚珠的合点端)的外种皮内侧与内种皮维管束索的连接点(图2-G), 而在外种皮背侧和侧面, 均未发现维管束分布(图2-A, C)。
2.2.2 内种皮解剖结构 内种皮包裹着胚乳, 是半透明膜质结构。在种阜端由内种皮内层和内种皮外层围成1个气室, 类似于鸡蛋的卵壳膜在一端所形成的气室(图2-F, G; 图4-C; 图7-A, B)。内种皮由5~6层细胞组成, 由外到内依次为马氏层(Malpighian layer, ML)、海绵组织层和内珠被内层, 其中马氏层为1层细胞, 海绵组织层由3~4扁平的薄壁细胞组成, 其中有维管束穿过, 而内珠被内层为1层薄壁细胞(图2-H, I)。
内种皮维管束分布很有规律。在未被染色时, 维管束呈黄褐色, 主要分布在合点端绝大部分面积的内种皮中, 而在种阜端内皮层中, 未见维管束分布(图1-B)。被番红染色后, 内种皮维管束木质部呈红色(图3-A), 其中与外种皮腹侧种脊大型维管束相连的维管束索, 分枝为3个较大的维管束(图3-B), 然后, 从合点端开始不断分枝变细, 终止于内种皮内层和外层结合处附近(图3-B~E), 这也进一步验证了上述在种阜端的内种皮内层和外层部分无维管束分布的结果(图3-A, E)。外种皮种脊中的大型维管束和内种皮的各级维管束, 共同构成种皮维管系统。
2.2.3 种阜的解剖结构 种阜是外种皮在种子一端延伸形成膨大的浅棕色海绵状隆起物(图1-A; 图4-A~C; 图7-A, B)。从种阜上部看, 种阜是由2个浅棕色半球状的结构相互连接构成的(图4-A, B)。种阜由薄壁细胞构成, 外侧3层细胞比内部细胞小, 细胞壁上有发达的纹孔, 纹孔来源于增厚的果胶质(图4-D~G)。在种阜中部横切面腹侧, 有2个维管束斜向分布, 构成种阜维管系统, 其中管胞也被希夫试剂染成红色(图4-H, J); 稍靠种阜背侧中部有一个种阜管道, 呈“丫”字形(图4-H, I)。在种阜纵切面上, 种阜管道外端为盲端, 内端开口于内种皮的气室(图4-C, K, L; 图7-A), 该管道壁细胞较小, 沿管道呈径向排列(图4-H, I)。从染色情况看, 种阜细胞壁由不可溶性糖(纤维素和半纤维素, 具亲水性)和果胶构成(图4-D~L)。
2.3 胚乳和胚的解剖结构
胚乳呈白色, 光滑饱满, 占种子体积的绝大部分, 以不同厚度包裹着胚(图1-C; 图7-A, B)。胚由子叶、胚芽、胚轴和胚根组成(图1-D; 图7-A, B)。子叶2片, 紧贴胚乳, 薄而大, 主脉纹明显, 侧脉分布不对称(图1-E; 图7-B)。2片子叶基部之间是胚芽(图5-I; 图6-C, F)。胚根呈光滑圆锥形, 胚轴连接胚芽与胚根(图1-E; 图6-C; 图7-A, B)。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5希夫试剂染色的蓖麻胚的解剖结构
A: 子叶中部横切面, 示2片子叶被胚乳包围的格局; B: 图A的局部放大图, 示胚乳细胞中大量含有拟晶体和磷酸盐球形体的糊粉粒; C: 图A中胚乳边缘的局部放大图, 示外侧9~11层小型胚乳细胞和内侧大型的胚乳细胞, 且均含有糊粉粒; D: 图(A)中子叶中段横切面, 示含有糊粉粒的小型的子叶细胞、大型的胚乳细胞以及子叶和胚乳之间的粘液层(箭头); E: 胚根横切面, 示基本分生组织和原形成层; F: 图E的局部放大图, 示基本分生组织和原形成层细胞缺少糊粉粒; G: 胚轴横切面, 示皮层、原形成层和维管柱; H: 图G的局部放大图, 示皮层、原形成层和维管柱细胞缺少糊粉粒; I: 胚芽横切面, 示2个胚芽突起被包围在2片子叶主脉之间; J: 图I的局部放大图, 示胚芽细胞缺少糊粉粒; K: 距图I所在平面上面一点距离的子叶基部横切面, 示2片子叶主脉结构; L: 图K的局部放大图, 示主脉细胞缺少糊粉粒。图中圆圈代表外韧维管束。GM: 基本分生组织; PR: 原形成层; CX: 皮层; PL: 胚芽; VC: 维管柱。
Fig. 5Anatomical structure of R. communis embryos stained with periodic acid Schiff (PAS)
A: transection of middle cotyledon, showing the pattern of two cotyledons surrounded by endosperm; B: local enlargement of figure A, showing plenty of aleurone grains containing crystalloids and phosphate spheres in endosperm cells; C: local enlargement of endosperm margin in figure A, showing the outer 9-11 layers of small endosperm cells and the inner large ones containing aleurone grains; D: transection of the middle cotyledon in Figure A, showing small cotyledon cells and large endosperm ones, which contained aleurone grains, and the mucilaginous layer between the cotyledon and endosperm (arrow). E: transection of the radicle, showing ground meristem and procambia; F: local enlargement of figure E, showing ground meristem and procambia cells lacking aleurone grains; G: transection of the hypocotyle, showing cortex, procambia and vascular cylomnder; H: local enlargement of figure G, showing cortex, procambia and vascular cylomnder cells lacking aleurone grains; I: transection of plumule, showing two plumule protuberances surrounded by two main veins of cotyledons; J: local enlargement of figure I, showing plumule cells lacking aleurone grains; K: transection a short distance above the level in figure I in cotyledon base, showing anatomical structure of main veins of two cotyledons; L: cells in main veins lacking aleurone grains. The circles in the figure represented collateral vascular bundles. GM: ground meristem; PR: procambia; CX: cortex; PL: plumule; VC: vascular cylinder.
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6番红固绿双重染色的蓖麻胚的解剖结构
A和B: 种子中部的横切面; C~F: 垂直于子叶所在平面的纵切面。A: 子叶中段横切, 示2片子叶分离、子叶与胚乳通过粘液层(箭头)相互连接的格局, 以及大型胚乳细胞和小型子叶细胞均含糊粉粒; B: 胚乳横切面, 示含大量糊粉粒的胚乳细胞; C: 去大部分子叶的胚的纵切面, 示部分子叶、胚芽、胚轴和胚根; D: 图C中胚根纵切面, 示原表皮、基本分生组织和原形成层的分化格局; E: 图C中胚轴的纵切面, 示表皮、皮层和维管柱的分化格局; F: 胚芽和部分子叶的纵切面, 示胚芽和联系胚轴与子叶的维管束(箭头)。
Fig. 6Anatomical structure of R. communis embryos stained with saffron and green fixation
A and B: the transverse section of the middle part of the seed; C-F: longitudinal section perpendicular to the plane of the cotyledon. A: transection of the middle cotyledon, showing the patterns of cotyledon separation, cotyledon-endosperm interconnection through the mucilaginous layer (arrows), large endosperm cells and small cotyledon ones with aleurone grains; B: transection of the endosperm, showing endosperm cells containing large amounts of aleurone grains; C: longitudinal section of embryos with most cotyledons removed, showing partial cotyledon, plumule, hypocotyle and radicle; D: longitudinal section of the radicle in figure C, showing differentiation pattern of primary epidermis, ground meristem and procambium; E: longitudinal section of the hypocotyle in figure C, showing differentiation pattern of epidermis, cortex and vascular cylomnder; F: longitudinal section of plumule and partial cotyledon, showing the plumule and vascular bundles (arrows) connecting the hypocotyle and cotyledon.
图7
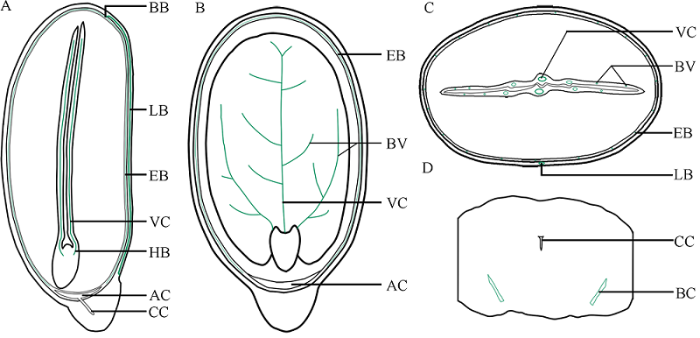 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7蓖麻种子维管束分布示意图
A: 与子叶面垂直的正中纵切面, 示内、外种皮维管束的关系及来源于胚轴的子叶主脉维管束; B: 与子叶面平行的正中纵切面, 示内种皮维管束和子叶维管束空间上的分离; C: 与种子长轴垂直的正中横切面, 示外种皮、内种皮和子叶维管束的空间关系; D: 种阜中部局部横切面, 示种阜维管束。VC: 子叶主脉维管束; HB: 胚轴维管束; AC: 气室; BV: 子叶分枝维管束; BC: 种阜维管束。
Fig. 7Diagram showing the distribution of vascular bundles in R. communis seed
A: median longitudinal section perpendicular to the plane of cotyledon, showing the relationship between vascular bundles of endosperm and episperm and main vascular bundles of cotyledons originating from hypocotyle; B: median longitudinal section parallel to the plane of cotyledon, showing spatial separation of vascular bundles in endopleura and cotyledon; C: median transverse section perpendicular to the long axis of seed, showing spatial relationship of vascular bundles in episperm, endopleura and cotyledon; D: local transverse section of middle part of caruncle, showing bundles in caruncle. VC: the vascular bundles in the main vein of cotyledons; HB: hypocotyle bundles; AC: air chamber; BV: branched vascular bundles in cotyledons; BC: bundles in the caruncle.
胚乳由表皮和胚乳组织构成, 其中未发现维管束。胚乳细胞都含有大量的含有拟晶体和磷酸盐球形体的糊粉粒(图5-A~D), 具有贮藏营养物质的功能。胚乳细胞的细胞壁由果胶、纤维素和半纤维素等亲水性物质构成(图5; 图6-A, B)。
从胚、胚乳及其各部分的连接情况看, 胚轴和胚根在空间上与胚乳分离(图5-E, G, I~K), 而2片子叶近轴面相互分离, 但其远轴面分别通过非细胞结构的粘液层与胚乳连接在一起(图5-A, D, 图6-A)。
从胚维管束看, 胚根维管束尚未分化(图5-E, F; 图6-C, D), 而胚轴已分化出8个外韧维管束(图5-G, H), 它们通过2片子叶主脉基部的8个维管束(图5-I~K; 图6-C, F; 图7-A), 与子叶各级侧脉维管束(图1-E; 图5-A; 图7-B)相连接, 构成胚维管系统。其中, 每片子叶主脉基部的4个维管束在延伸过程中由粗变细(图5-K, L), 数量由4个减少到1个。
从胚细胞所含糊粉粒看, 子叶细胞中亦含有大量糊粉粒, 但胚根(图5-E, F)、胚轴(图6-C, E)和胚芽(图6-I, J)细胞中很鲜见。
从细胞壁组成看, 胚细胞的细胞壁也由果胶、纤维素和半纤维素等亲水性物质构成(图5; 图6)。
3 讨论
本研究发现, 蓖麻种子的种孔并未被种阜覆盖, 种阜端的内种皮具一气室, 种阜管道内端开口于该气室; 2片子叶近轴面分离, 远轴面则通过粘液层与胚乳相连接; 胚乳与子叶的细胞均含糊粉粒, 而在胚芽、胚轴和胚根细胞中未发现; 胚轴和胚根与胚乳相分离; 种皮维管系统、种阜维管系统和胚维管系统在空间上相对独立、功能上相互联系。3.1 种皮
种孔是由胚珠顶端的珠孔发育而成的孔道, 为种子萌发时吸收水分和胚根伸出的部位。有的《植物学》教材[18,19]认为种孔被种阜所覆盖, 只有剥去种阜才能看到, 这与本研究的结果不一致。事实上, 如果种阜覆盖住种孔, 意味着种阜将成为胚根伸出种皮的阻碍, 那么, 只有种阜被破坏或胚根经过弯曲才能突破种皮, 这样, 将削弱种阜在种子萌发中的吸水功能, 或影响胚根迅速扎入土壤, 这不符合植物结构与功能相适应的观点。因此, 蓖麻种子的种孔不可能被种阜所覆盖。本研究中种皮结构与Bianchini和Pacini[13]结果相比较, 不一致之处是, (1)外种皮表皮细胞明显径向伸长, 因此推测授粉50 d后可能是表皮细胞径向生长的重要时期; (2)外种皮表皮角质层和种脊大型维管束木质部并未被番红染为红色, 该现象也在合欢种皮解剖结构观察中发现过[20], 可能是因为种皮角质层成分特殊和管状分子未木质化的缘故, 这还有待于进一步验证; (3)内种皮细胞不是2层, 而是5~6层。另外, 马炜梁[19]将蓖麻种子种皮分为外种皮、中种皮和内种皮, 与本实验结果、Bianchini和Pacini[13]的结果以及国内其他《植物学》教材的描述均不一致。一方面, 我们为此认真研究了马炜梁蓖麻种皮图像的标注, 认为这可能是因为, 他误将外种皮的海绵组织层和栅栏组织层当作“中种皮”的缘故; 另一方面, 我们认为蓖麻种皮的发育还有待于进一步深入研究。特别是, 本研究发现, 内种皮维管组织发达, 在种阜端形成气室, 气室与种阜管道内口相连通, 这为蓖麻种子的水分运输提供了重要解剖学基础。
3.2 种阜
“elaiosome”是Sernander[21]提出的一个生态术语, 用来表示蚂蚁散布的所有肉质和可食用的植物附属物(传播体)。Bhojwani和Bhantnagar[22]在研究大戟科植物“elaiosome”的吸水功能时, 将外珠被增生形成的、具有吸收作用的一类“elaiosome”称为“carubcle”, 即种阜。所以, 《植物生物学词典》[23] 认为, 种阜是大戟科和无患子科植物受精后, 在珠柄处经细胞分裂与增大而形成的一种垫状结构。种阜的主要功能有两方面: (1)生态功能: 蚁媒传播(myrmecochory), 许多蚂蚁被种阜中的脂类、蛋白质、淀粉和维生素所吸引, 将种子携带到其巢穴, 使种子离开母体[3]; (2)生理功能: ①诱导休眠, 有的植物种子在去除种阜后才能迅速萌发, 但蓖麻种阜不具此功能[24]; ②促进种子脱水, 授粉40 d后, 种阜可将胚乳和胚细胞的水分转运到外界, 有利于种子寿命延长; ③促进种子吸水, 在种子萌发期间, 种阜从土壤吸收水分, 可暂时贮存, 或将其转移到种子的其余部分[13]。
本研究结果与Lisci等[14]报道基本一致, 但种阜维管系统和种阜管道是首次报道(图4-F, G, K, L)。蓖麻种阜适于种子成熟时干燥脱水以及发育与萌发过程中吸水的结构可能包括: (1)细胞壁组分, 种阜主要由细胞壁果胶增厚的薄壁细胞构成, 细胞壁组分是果胶、纤维素和半纤维素, 均属于亲水性物质; (2)纹孔, 种阜细胞壁纹孔发达, 有利于胞间水分流动; (3)维管束, 种阜维管束有利于水分在种阜内的长距离输导; (4)种阜管道, 它可能是种子萌发时, 水分和空气通过种阜管道的毛细作用和扩散作用进出种子的通道。
3.3 维管系统
在种子发育期间, 水分、无机盐和有机物等物质通过种皮维管系统, 从果实胎座被输送到胚乳和胚, 供胚乳和胚发育和贮藏; 种子萌发时, 水分亦可沿此路径进入种子, 使其吸胀而萌发(图7-A~C)。在蓖麻种子成熟过程中脱水和萌发过程中吸水时, 可通过种阜维管系统进行较长距离的水分输导(图7-D)。在种子萌发、形成幼苗的过程中, 子叶作为中介, 也可将胚乳贮藏的营养物质通过子叶各级分枝维管束、主脉维管束和胚轴维管束运输到胚的各个部分来利用; 子叶变绿能进行光合作用、制造有机物后, 也通过此途径将营养物质输导至胚根、胚轴和幼苗的其他部分来利用。当种子萌发、胚根具有吸水能力后, 水分和无机盐则沿着相反的路径运动(图7-A~C)。
3.4 胚乳与胚
被子植物胚乳是指胚囊中2个极核受精后发育形成的三倍体特殊营养结构, 贮藏有丰富的营养物质, 这些营养物质在种子萌发中由胚长成幼苗时被吸收利用, 或在胚的发生中立即被吸收消耗[22]。蓖麻成熟种子中, 胚乳细胞均含丰富的糊粉粒(图5-A, B; 图6-B)和圆球体(脂质体)[16]。子叶是一种侧生在胚轴上的、片状或线形的、特殊的吸收器官, 其功能是吸收和转移异养幼苗所需的大量物质。本研究中子叶的观察结果(图1-E; 图5-A; 图7-A~C)与Kriedemann和Beevers[15]的发现一致。从结构上看, 首先, 蓖麻胚乳和子叶与胚芽、胚轴和胚根细胞中糊粉粒分布具有显著差异(图5;图6), 这一方面反映其功能的不同, 另一方面反映其功能在时空上是分离的, 即胚乳和子叶在种子发育和萌发时分别承担营养贮藏、转化和转运功能, 而胚芽、胚轴和胚根则在种子萌发时进行生长和发育。其次, 2片子叶远轴面分别通过非细胞结构的粘液层与胚乳大面积相连接(图5-A, D; 图6-A), 这扩大了子叶和胚乳之间物质转运的面积, 有助于种子发育和萌发时物质的快速转运和贮藏。
从功能上看, 蓖麻胚乳与胚密切相关[15,25]。蔗糖在胚乳中合成, 通过自由空间(粘液层), 与质子协同转运(Cotransport of sucrose and protons)至子叶, 被子叶吸收, 成为子叶、胚轴、胚根和嫩枝生长和发育的碳源, 直到绿化的子叶可以进行同化物的生产和输送[26]。通常从胚乳进入子叶的糖分有80%以上被传递到胚轴[15]。可见, 在蓖麻种子萌发过程中, 糖的异生作用、转运和利用在空间上是明显分离的, 而且是单向运输。
蓖麻种子胚轴和胚根在空间上与胚乳分离(图5-E, K, I), 这种格局可消除种子萌发过程中, 胚轴向上伸长和胚根向下扎入土壤时, 胚乳对二者的生长发育可能造成的阻力, 有利于种子在合适的条件下迅速萌发。
4 结论
采用石蜡切片和显微照相技术全方位观察了成熟蓖麻种子种皮(包括外种皮、内种皮和种阜)、胚乳和胚的结构特征和维管系统的构成, 为全面掌握蓖麻成熟种子的结构和研究种子发育和萌发过程中的物质运输提供了结构证据。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
.
[本文引用: 4]
[本文引用: 2]
[本文引用: 4]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/BF00387548URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}