 ,*青海大学农林科学院 / 青海省春油菜遗传改良重点实验室 / 国家油菜改良中心青海分中心 / 农业部春油菜科学观测实验站 / 青海省春油菜研究开发中心, 青海西宁 810016
,*青海大学农林科学院 / 青海省春油菜遗传改良重点实验室 / 国家油菜改良中心青海分中心 / 农业部春油菜科学观测实验站 / 青海省春油菜研究开发中心, 青海西宁 810016Comparative analysis on flower bud differentiation of multilocular and bilocular traits in Brassica juncea
LI Xin, XIAO Lu, LI Lin-Fang, DU De-Zhi,*Academy of Agricultural and Forestry Sciences, Qinghai University / Key Laboratory of Qinghai Province for Spring Rapeseed Genetic Improvement / The Qinghai Research Branch of the National Rapeseed Genetic Improvement Center / Spring and Rape Scientific Observation Experimental Station of Ministry of Agriculture and Rural Affair / Qinghai Research and Development Center for Spring Rapeseed, Xining 810016, Qinghai, China通讯作者:
第一联系人:
收稿日期:2018-09-19接受日期:2019-01-12网络出版日期:2019-02-08
| 基金资助: |
Received:2018-09-19Accepted:2019-01-12Online:2019-02-08
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (9036KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
李新, 肖麓, 李麟芳, 杜德志. 芥菜型多室与二室油菜花芽分化过程的比较分析[J]. 作物学报, 2019, 45(5): 705-713. doi:10.3724/SP.J.1006.2019.84121
LI Xin, XIAO Lu, LI Lin-Fang, DU De-Zhi.
我国油菜栽培类型有甘蓝型油菜、芥菜型油菜和白菜型油菜。三类油菜中存在多室资源, 其中青海芥菜型多室油菜除抗裂角、黄籽等优良性状外, 还具有稳定遗传的多室(3~5果室)性状[1,2]。多室性状与油菜产量紧密联系, 众所周知, 油菜产量是由单位面积角果数、每角粒数和千粒重构成。普通芥菜型二室油菜的角果果室为2个, 多室油菜随着角果果室数目的增加, 其每角粒数显著高于相同遗传背景下二室油菜的每角粒数, 从而单株产量高于普通二室油菜[2,3,4,5,6,7]。
花芽分化是有花植物发育中最关键的阶段, 同时也是一个复杂的形态建成过程。油菜花芽分化是在油菜体内外因子的共同作用和相互协调下, 由量变到质变的过程, 花芽分化的迟早及分化数量也会影响油菜产量[8,9,10]。前人将白菜型油菜花芽分化划分为7~8个时期, 分化起始的叶片数为2~3片[11,12]; 将甘蓝型油菜花芽分化过程划分为8个时期15个阶段, 其开始分化的叶片数为9~11片[11,12,13]。相比白菜型及甘蓝型油菜, 对芥菜型油菜花芽分化研究未有报道。
多室油菜的解剖学研究主要包括对角果形态的观察以及假隔膜和心皮数目的统计[14,15,16]。吕泽文等[4]对甘肃芥菜型多室油菜角果解剖观察发现, 多室角果形态短粗, 含4个果瓣, 角果被2个平行排列的呈 “II” 型的假隔膜分隔为3个腔室。朱彦涛等[17]对甘蓝型多室角果研究发现, 多室角果呈三棱态, 角果皮三瓣, 被呈 “Y” 状的假隔膜分隔为三室。赵会彦[18]对青海芥菜型多室油菜角果解剖观察发现, 多室角果粗短, 整体呈圆筒形, 含4个果瓣, 两层假隔膜将多室角果分为3个腔室, 包括1个大腔室, 2个小腔室。
多室性状作为油菜中一优异性状, 其遗传特性及基因的定位研究有诸多报道[19,20,21,22,23,24,25,26,27], 但对多室角果的形态建成过程, 尤其对关键的花芽分化过程未有系统的研究报道。本研究观察花芽分化全过程, 清晰认识多室角果的形态建成过程, 分析二室和多室性状的分化差异, 了解多室性状与产量的相关性, 对制定合理的栽培措施及进行花期调控等具有指导意义, 可为农业育种工作中多室材料的提前选择带来便利。
1 材料与方法
1.1 试验材料与设计
选自青海省农林科学院春油菜研究所的青海芥菜型多室油菜和二室亲本“塔油2号”, 用于阐述二室及多室油菜花芽分化全过程。皆具黄籽、抗裂角等优异性状, 遗传性稳定, 前者较后者生育期晚。以多室油菜为轮回亲本, “塔油2号”为供体非轮回亲本连续回交3次, 构建多室与二室为1∶1的BC3F1分离群体, 从中随机选择二室单株自交, 随后在BC3F2群体中使用分子标记辅助选择纯合的二室单株和多室单株自交3代, 获得BC3F5群体。在相同遗传背景下, 比较纯合的二室及多室油菜花芽组织的分化差异。
试验于2016—2017年于青海省农林科学院试验地进行, 其海拔2225 m, 36.34°N, 101.49°E。小区面积31.2 m2 (15.6 m × 2.0 m), 每公顷密度为8380株。准备培养皿并放入清水浸湿的滤纸, 将种子放入培养皿内, 置光照培养架, 于22℃条件下共培养30 h (16 h光照, 8 h黑暗)后, 播种于试验地, 记录播种日期。
播种2 d后观察并记录出苗日期; 出苗4 d后, 待2片真叶展开后, 分别取二室及多室油菜生长锥于显微镜下观察, 记录分化状态; 2片真叶完全展开后, 每隔5 d, 取生长发育一致的新鲜植株3株, 依次对生长锥、花芽组织、雌蕊、柱头等取材, 记录每个分化时期的叶片数; 花芽分化进入中后期后, 分别取二室及多室油菜不同直径大小的花蕾(1、2、3和4 mm), 观察其柱头及子房内籽粒排列结构; 角果形成后, 解剖观察二室及多室角果内部结构。
1.2 采样和制片方法
将芥菜型油菜单株从试验地连根拔出, 用清水洗干净, 在体式显微镜下逐个解剖, 观察并记录花芽发育的动态。切取多室和二室油菜同一时期的生长锥、花芽组织、柱头及不同大小的花蕾。将各组织的一部分样品用FAA溶液固定, 做石蜡切片; 另一部分用2.5%的戊二醛溶液固定, 用于后期扫描电镜的样品制备。1.2.1 石蜡切片法 FAA固定液(固定24 h以上, 置于4℃保存)→酒精洗脱(70%洗脱2次, 85%洗脱1次, 95%洗脱1次, 100%洗脱2次, 每次1.5 h)→透明(1/2 V无水乙醇+1/2 V二甲苯混合液处理1次, 100%二甲苯洗脱1次, 每次1.5 h)→浸蜡(石蜡置于70℃恒温箱溶蜡, 将固定瓶内的处理液倒净, 用1/3 V二甲苯+2/3 V石蜡混合液浸蜡, 置于37℃恒温箱(3~4 d)→包埋→切片→脱蜡(二甲苯)→染色(置于番红染液2 h)→透明→封片及照相(Nikon DS-Fi1) [28]。
1.2.2 扫描电镜检测 2.5%戊二醛固定液(抽真空后置于4℃冰箱, 固定15 d以上)→漂洗(0.2 mol L-1缓冲液, 漂洗3次, 每次15 min)→1%锇酸固定(夏季1 h, 冬季2 h)→酒精脱水(30%、45%、60%、75%和90%梯度脱水, 每级5 min)→冷冻干燥→喷金→镜检照相(Jeol JSM-6610/LV) [29,30]。
1.2.3 体视镜下观察 解剖油菜角果使其内部种子排列及假隔膜清晰显现后观察(Nikon SWZ25)。
1.3 芥菜型二室及多室油菜花芽分化过程的比较
观察“塔油2号”(二室亲本)及青海芥菜型多室油菜(多室亲本)花芽分化全过程, 记录开始分化的时间、不同花芽组织的分化时间和各时期的叶片数。比较BC3F5群体中纯合二室油菜和纯合多室油菜的SAM、FM、心皮、子房腔、假隔膜、柱头等组织。对22组(二室和多室)SAM直径和22组(二室和多室) FM直径进行测微。
1.4 数据统计与分析
利用Microsoft Excel 2007和SPSS 16.0统计软件分析试验数据。用最小显著差法(LSD0.05)检验平均数。2 结果与分析
2.1 芥菜型二室油菜(亲本)花芽分化进程
芥菜型二室油菜自出苗开始, 约21 d左右, 平均叶片数为4.5 (4~5), 日平均温度为11.7℃时, 开始分化, 过程分为5个时期。(1)生长锥伸长期。生长锥作为油菜的茎顶端生长点, 是油菜生长过程中最为关键的组织。4月上旬催芽的油菜种, 播于西宁试验地。芽播2 d后开始出苗, 此时, 叶片为3片, 生长锥呈半圆锥形(图1-a: SAM), 不断伸长膨大, 四周被不断分化出来的的叶原基包围(图1-a: L), 无花原基出现。此阶段维持20 d左右。(2)花原基出现。生长锥伸长20 d后, 叶片数为4.5, 生长锥顶端周围依次长出1~6个花原基(图1-b: f1~f6)。此阶段持续6 d左右。花原基的出现, 预示着油菜花器官开始分化, 从营养生长进入生殖生长的阶段。(3)花萼原基分化。油菜为无限花序, 生长锥表面不断分化出花原基(图1-c~e; f)。4月下旬, 出苗27 d左右, 平均叶片数为5, 随着花原基的依次分化, 第一花原基伸长膨大且出现4片半新月形对称凸起, 该凸起为萼片原基(图1-f: S1~S4), 此时期油菜的分化已从茎顶端转移到花顶端, 而后只对花顶端观察研究, 以解析花器官的分化过程。此阶段持续5 d左右。(4)花瓣原基分化。5月初, 出苗33 d左右, 平均叶片数为5.5, 花萼原基不断伸长, 直至完全包裹花芽顶端(图1-g: S1~S4), 由于花芽顶端的伸长膨大, 其基部已出现花瓣原基, 小而不清晰(图1-h: 箭头所指), 结合石蜡切片纵切面观察发现, 花芽顶端侧基部出现圆形凸起, 该凸起为花瓣原基(图3-c: P)。此阶段维持5 d左右。(5)雌雄蕊形成。花瓣原基出现不久后, 花瓣原基基部腋间出现雄蕊(图1-h: St1~St6 )。随着雄蕊的不断发育, 心皮即将分化, 5月中旬, 45 d左右, 平均叶片数7片时, 花顶端中心生长锥向内凹陷, 呈唇形态, 形成两块对称的心皮(图1-h: C1, C2), 随着心皮的不断发育, 中下部发育成子房腔(图1-i: OV), 上部不断伸长形成花柱(图1-i~k), 花柱顶端的凹陷逐渐呈闭合状态, 最终形成柱头(图1-l: 箭头所指)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1芥菜型二室油菜花芽分化进程图
a: 生长锥伸长, 茎顶端分生组织(SAM)、叶原基(L); 图b: 花原基出现, 茎顶端分生组织(SAM)、花原基(f1~f6); 图c~图e: 花原基不断分化膨大, 茎顶端分生组织(SAM)、花原基(f)、花顶端分生组织(FM)、萼片原基(S); 图f: 花萼原基分化, 花顶端分生组织(FM)、萼片原基(S1~S4); 图g: 花萼(S1~S4), 包裹花顶端; 图h: 花瓣原基分化, 花瓣原基(P)、雄蕊原基(St1~St6)和心皮原基(C1, C2)分化; 图i~图k: 子房腔(OV)的形成及花柱伸长; 图l: 柱头(箭头所指)形成。
Fig. 1Flowering bud differentiation of bilocular B. juncea
Fig. a: growth cone elongation, showing shoot apical meristem (SAM) and leaf primordial (L); Fig. b: flower primordia appear, showing shoot apical meristem (SAM) and flower primordia (f1-f6); Figs. c-e: continuously differentiated and expanded flower primordia, showing shoot apical meristem (SAM), flower primordia (f), flower meristem (FM) and sepal primordia (S); Fig. f: sepal primordia differentiation, showing flower meristem (FM) and sepal primordia (S1-S4); Fig. g: the top of flower wrapped by sepal primordia (S1-S4); Fig. h: petal primordia differentiation, showing petal primordia (P), stamen primordia (St1-St6) and carpel primordia (C1, C2); Figs. i-k: ovary(OV) formation and stigma elongation; Fig. l: the stigma formation (indicated by the arrow).
2.2 芥菜型多室油菜(亲本)花芽分化进程
芥菜型多室油菜自出苗开始, 约22 d左右, 日平均温度为11.7℃, 平均叶片数为5时, 开始分化, 过程分为5个时期。(1)生长锥伸长期。自出苗2 d, 叶片数为3时, 生长锥呈圆锥形(图2-a: SAM), 不断伸长膨大, 四周被叶原基包围(图2-a: L), 此阶段持续20 d左右。(2)花原基出现。生长锥持续伸长膨大, 约22 d后, 生长锥侧表面长出花原基, 并随着第一花原基的不断膨大, 第二、第三、第四等花原基依次分化出来(图2-b: F1~F6)。该阶段叶片数为4.5, 持续8 d左右。(3)花萼原基分化。随着花原基的依次分化, 花原基数量不断增多(图2-c~e: F)。平均叶片数为5时, 第一花原基伸长且膨大, 侧基部出现4片半新月形对称凸起, 该凸起为萼片原基(图2-f: S1~S4)。该阶段持续5~7 d左右。随着第一花原基上花萼原基的分化, 多室油菜的分化从茎顶端转移到花顶端, 象征着营养生长结束, 生殖生长开始。 (4)花瓣原基分化。自出苗38 d左右, 平均叶片数为6时, 花萼原基不断伸长, 直至完全包裹花芽顶端(图2-g: S1~S4), 通过石蜡纵切面切片观察到, 花芽顶端侧基部出现对称性圆形凸起, 该凸起为花瓣原基(图3-f: P), 结合扫描电镜观察, 可见刚开始分化的花瓣原基(图2-h: P)。该阶段持续4 d左右。(5)雌雄蕊形成。花瓣原基基部腋间出现雄蕊(图2-h: St1~St6), 其后, 心皮随之分化, 5月中旬, 52 d左右, 平均叶片数为7.5时, 花芽顶端发生结构性变化, 花芽中心生长锥向内凹陷成“X”状, 形成4个心皮(图2-h: C1~C4), 并将花芽中心生长锥分隔为4个区域。随着心皮不断分化, 下部发育成子房腔(图2-i~k: OV), 上部伸长后形成花柱并逐渐形成柱头(图2-l: 箭头所指)。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2芥菜型多室油菜花芽分化进程图
a: 生长锥伸长, 茎顶端分生组织(SAM)、叶原基(L); 图b: 花原基出现, 茎顶端分生组织(SAM)、花原基(F1~F6); 图c~e: 花原基不断分化膨大, 茎顶端分生组织(SAM)、花原基(F)、花顶端分生组织(FM)、萼片原基(S); 图f: 花萼原基分化, 花顶端分生组织(FM)、萼片原基(S1~S4); 图g: 花萼(S1~S4), 包裹花顶端; 图h: 花瓣原基分化, 花瓣原基(P)、雄蕊原基(St1~St6)和心皮原基(C1~C4)分化; 图i~k: 子房腔(OV)的形成及花柱伸长; 图k~l: 柱头(箭头所指)形成。
Fig. 2Flowering bud differentiation of multilocular B. juncea
Fig. a: growth cone elongation, showing shoot apical meristem (SAM) and leaf primordial (L); Fig. b: flower primordia appear, showing shoot apical meristem (SAM) and flower primordia (F1-F6); Figs. c-e: continuously differentiated and expanded flower primordial, showing shoot apical meristem (SAM), flower primordia (F), flower meristem (FM) and sepal primordia (S); Fig. f: sepal primordia differentiation, showing flower meristem (FM) and sepal primordia (S1-S4); Fig. g: the top of flower wrapped by sepal primordia (S1-S4); Fig. h: petal primordia differentiation, showing petal primordia (P), stamen primordia (St1-St6) and carpel primordia (C1-C4); Figs. i-k: ovary (OV) formation and stigma elongation; Fig. l: stigma formation (indicated by the arrow).
观察发现, 在西宁试验田环境下, 二室油菜自出苗开始, 约21 d左右, 平均叶片数为4.5 (4~5)时, 花芽开始分化且过程分为5个时期; 多室油菜自出苗开始, 约22 d, 平均叶片数为5时, 花芽开始分化且整个分化过程与二室油菜的分化相比会推迟7 d左右(表1), 可以清晰看到多室角果在形态建成初期时的分化方式。
Table 1
表1
表1大田环境二室和多室油菜花芽组织的分化时间
Table 1
| 类型 Type | 生长锥 Growth cone | 花原基 Flower primordium | 萼片原基 Sepal primordium | 花瓣原基 Petal primordium | 雄蕊原基 Stamen primordium | 心皮 Carpel |
|---|---|---|---|---|---|---|
| 二室Bilocular | 2 | 21 | 27 | 33 | 40 | 45 |
| 多室Multilocular | 2 | 22 | 31 | 38 | 51 | 52 |
新窗口打开|下载CSV
2.3 多室与二室油菜花芽分化比较
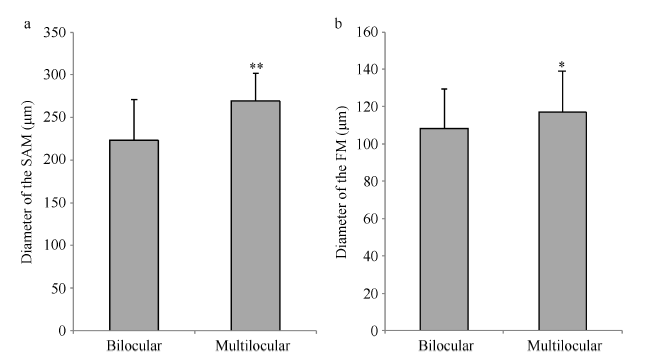
了解亲本间花芽分化全过程后, 观察比较BC3F5群体中同一发育阶段的纯合二室单株和多室单株的茎顶端分生组织(SAM)、花顶端分生组织(FM)、心皮、子房腔、假隔膜和柱头等组织, 发现在这几个组织中均有分化差异。(1)通过对22组生长锥顶端石蜡切片结合纵切面测微比较发现, 二室油菜SAM直径均值为223.07 μm (图3-a, b), 多室油菜SAM直径均值为269.50 μm (图3-d, e), 后者显著大于前者(图4-a)。因此, 二室油菜和多室油菜出现分化差异的最早组织是SAM, 该差异一直维持到花芽原基的出现。(2)随着生长锥基部不断分化出的花原基伸长膨大, 且第一花芽原基顶端完全被萼片原基包裹, 石蜡切片结合纵切面测微比较发现, 二室油菜FM直径均值为108.10 μm (图3-c), 多室油菜FM直径均值为119.53 μm (图3-f), 多室FM的直径大小显著大于二室FM的直径大小(图4-b), 类似于SAM的分化差异。因此, 可认为多室角果的出现与茎顶端分生组织和花顶端分生组织的膨大分化联系紧密。(3)心皮分化时期, 花芽顶端分生组织发生结构性变化。二室花芽顶端向内凹陷成“I”状裂口, 形成2块心皮(图3-g: C1, C2), 将花芽中心生长锥一分为二, 随后, 二室对半心皮将花芽中心生长锥分隔成2个大小基本相等的腔室(图3-h: lc), 中间是一列假隔膜(图3-n), 两侧各着生一列籽粒(图3-i: O), 最终形成含2个心皮和2列籽粒的二室角果(图3-m); 而多室花芽顶端向内凹陷成“X”状裂口, 形成4个心皮(图3-j: C1~C4), 将花芽中心生长锥分隔为4个区域, 随之逐渐形成相对应的4个子房腔室(图3-k: lc), 并伴有2个平行式的隔膜而着生4列种子(图3-l, 图3-q), 最终形成含有4个心皮和4列籽粒的四室角果(图3-p)。(4)花柱不断伸长后其顶端形成柱头, 柱头顶端呈现 “I” 及 “X” 状凹陷(图3-o, r), 这种结构发育稳定, 呼应了与子房腔内的结构, 因此, 可作为一种指示性状用以早期区分是二室油菜还是多室油菜。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3多室与二室花芽分化比较
图a~b: 二室茎顶端分生组织; 图c: 二室花顶端分生组织; 图d~e: 多室茎顶端分生组织; 图f: 多室花顶端分生组织; 图g: 二室心皮分化(C1, C2); 图h: 二室子房腔(lc); 图i: 二室籽粒着生(O); 图j: 多室心皮分化(C1~C4); 图k: 多室子房腔(lc); 图l: 多室籽粒着生(O); 图m~n: 二室角果内部结构; 图o: 二室柱头; 图p~q: 多室角果内籽粒排列结构; 图r: 多室柱头; P: 花瓣原基; St: 雄蕊原基; C: 心皮; lc: 心室; O: 胚珠; M: 中间区域; L: 横向区域。
Fig. 3Comparison of flower bud differentiation between bilocular B. juncea and multilocular B. juncea
Figs. a-b: the shoot apical meristem of the bilocular rapeseed; Fig. c: the flower meristem of the bilocular rapeseed; Figs. d-e: the shoot apical meristem of the multilocular rapeseed ; Fig. f: the Flower meristem of multilocular rapeseed; Fig. g: the stage of carpel differentiation of the bilocular rapeseed (C1, C2); Fig. h: the ovary formation of the bilocular rapeseed ; Fig. i: seed arrangement structure of the bilocular rapeseed (O); Fig. j: the stage of carpel differentiation of the multilocular rapeseed (C1-C4); Fig. k: the ovary formation of the multilocular rapeseed; Fig. l: seed arrangement structure of the multilocular rapeseed (O); Figs. m-n: internal structure of the bilocular rapeseed; Fig. o: the stigma of the bilocular rapeseed; Figs. p-q: internal structure of the multilocular rapeseed; Fig. r: the stigma of the multilocular rapeseed; P: petal primordia; St: stamen primordia; C: carpel; lc: locule; O: ovule; M: medial region; L: lateral region.
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4二室与多室SAM及FM的比较
**表示在0.01概率水平上显著相关; *表示在0.05概率水平上显著相关。
Fig. 4Comparison of SAM and FM between multilocular plants and bilocular plants
** Significant correlation at the 0.01 probability level; * Significant correlation at the 0.05 probability level.
3 讨论
众所周知, 芥菜型多室油菜具有诸多优良性状, 尤其是多室性状, 与油菜产量正相关[31,32,33]。花芽分化是有花植物发育中最关键的阶段, 油菜花芽分化的迟早影响其产量及性状产生。而目前多室油菜的研究主要集中在遗传特性及相关基因的定位克隆, 对多室油菜发育中花芽分化过程研究未有报道, 前人对甘蓝型和白菜型油菜花芽分化的研究结果丰富且年代久远, 对芥菜型油菜花芽分化研究未有报道。本研究划分了油菜三大栽培种之一的芥菜型油菜花芽分化的全过程, 记录了各时期的平均叶片数及进入分化的时间, 相比多数品种的甘蓝型及白菜型油菜, 青海芥菜型多室油菜进入花芽分化的时期在白菜型油菜之后、甘蓝型油菜之前, 对芥菜型油菜分化规律的研究具有指导意义。一直以来, 多室油菜的遗传特性是****们研究的关键, 近年来, 随着遗传特性的稳定, 对多室油菜进行了诸多角果解剖学研究, 试从角果内籽粒的排列结构及心皮数目的差异来解释和区分多室油菜, 本研究发现, 多室与二室油菜最早期的性状差异是SAM和FM直径的差异, 这种差异与顶端分生组织的膨大联系紧密。油菜的心皮起源于花顶端分生组织(FM), 而花顶端分生组织FM又始于茎顶端分生组织(SAM)的侧基部, 由此可知, 油菜一系列的器官组织发育是通过SAM中心区域内干细胞的不断分裂实现的, 在维持油菜的营养生长中, 茎端分生组织持续产生新的细胞, 来补充内因细胞分化而迁移出的细胞[34,35,36]。这种细胞分裂必须维持动态平衡, 保证茎端分生组织行使正常的功能, 这种动态平衡的实现需要一系列关键基因的精确调控, 目前在拟南芥及番茄中报道了很多与此相关的基因[37,38,39,40]。多室性状与SAM、FM的分化有关, 为此, 我们可在模式植物及相关植物中找到与分生组织相关联的基因, 了解多室性状在不同物种中的表现形式, 来解释多室性状的形成机制。
雌蕊的形成始于花顶端分生组织的不断分化, 中下部形成子房腔及心皮, 上部形成花柱及柱头, 普通油菜的子房腔, 其花芽生长锥顶端不断向内凹陷成嘴唇形, 形成2个心皮, 假隔膜从凹陷缝隙处分化后将子房腔分隔成2个腔室, 形成具2个心室的普通油菜, 而这种结构形态延伸发育到柱头表面。多室油菜的雌蕊分化类似于普通二室油菜, 子房腔向内凹陷成“X”状, 形成4个心皮, 且柱头上也有该形态, 将柱头分成4个区域。因此, 柱头上特殊的形态也可代表子房腔内的心室数、种子的排列数, 而柱头作为早期肉眼可观察的油菜组织, 可以直接区分二室或多室角果, 便于农业育种工作中多室性状的提前选择。
温度和光照影响花芽分化各阶段花芽数量, 以及花芽分化过程中一些生理生化指标, 影响其产量结构[8]。不同温度环境下, 油菜花芽分化的时间长短不一, 性状表现也存在差异。油菜单株中出现二室和多室角果的嵌合型现象与其角果的假隔膜及心皮的分化有关, 而假隔膜和心皮的分化决定了子房结构, 不同温度及光照环境影响花芽分化的生理指标及分化时间的长短, 尤其温度变化对嵌合型角果类型的出现有很大影响, 继而有必要进一步深入探究不同温度环境和光照时长对多室油菜性状表现的影响。
4 结论
青海芥菜型多室油菜自出苗开始, 约22 d左右, 平均叶片数为5时, 开始花芽分化且经历5个时期。二室与多室性状差异始于茎顶端分生组织(SAM)的直径差异, 多室大于二室; 而花顶端分生组织(FM)也表现出一致的差异。心皮分化时期, 多室花芽中心生长锥向内凹陷成“X”状, 形成具四心皮分隔的4块区域, 而二室花芽中心生长锥向内凹陷成“I”状, 形成二心皮的对半区域。二室的凹陷对应一列隔膜将子房内腔分隔成 2个大小基本相等的腔室, 含2列籽粒。多室中, “X”的凹陷对应其4列假隔膜将子房分隔成4个腔室, 含四列籽粒。二室柱头顶端形成 “I” 凹陷, 多室柱头顶端形成“X”状凹陷。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
Magsci [本文引用: 1]

<p>花芽分化是有花植物发育中最为关键的阶段,同时也是一个复杂的形态建成过程。这一过程是在植物体内外因子的共同作用和相互协调下完成的。了解植物花芽分化的机理对于制定合理的栽培措施、进行花期调控等具有重要意义。本文就植物花芽分化过程中的形态结构、环境因素、植物体自身因素以及生长调节剂对花芽分化的影响及植物花芽分化的调节机制做了一个概述,并对花芽分化过程中基因对成花的作用做了简要探讨。</p>
Magsci [本文引用: 1]
<p>花芽分化是有花植物发育中最为关键的阶段,同时也是一个复杂的形态建成过程。这一过程是在植物体内外因子的共同作用和相互协调下完成的。了解植物花芽分化的机理对于制定合理的栽培措施、进行花期调控等具有重要意义。本文就植物花芽分化过程中的形态结构、环境因素、植物体自身因素以及生长调节剂对花芽分化的影响及植物花芽分化的调节机制做了一个概述,并对花芽分化过程中基因对成花的作用做了简要探讨。</p>
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}