 ,1,2,*, 沈雪芳,1,2,*
,1,2,*, 沈雪芳,1,2,*Genetic diversity and population structure analysis by SSR markers in waxy maize
LU Yuan1,2, AI Wei-Da3, HAN Qing1,2, WANG Yi-Fa1, LI Hong-Yang4, QU Yu-Ji5, SHI Biao,1,2,*, SHEN Xue-Fang,1,2,*通讯作者:
收稿日期:2018-01-15接受日期:2018-10-8网络出版日期:2018-11-03
| 基金资助: |
Received:2018-01-15Accepted:2018-10-8Online:2018-11-03
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (831KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
卢媛, 艾为大, 韩晴, 王义发, 李宏杨, 瞿玉玑, 施标, 沈雪芳. 糯玉米自交系SSR标记遗传多样性及群体遗传结构分析[J]. 作物学报, 2019, 45(2): 214-224. doi:10.3724/SP.J.1006.2019.83008
LU Yuan, AI Wei-Da, HAN Qing, WANG Yi-Fa, LI Hong-Yang, QU Yu-Ji, SHI Biao, SHEN Xue-Fang.
玉米(Zea mays L.)是全球三大重要作物之一, 糯玉米(Zea mays L. var. ceratina Kulesh)是玉米属的一个亚种。糯玉米富含支链淀粉、营养丰富、适口性良好等优良特性赋予其宝贵的价值和广泛的用途, 现已成为深受欢迎的食品和工业原料, 具有较高的经济价值。
中国是公认的糯玉米起源中心[1]。曾孟潜(1987)论证认为早在1760年前, 中国已种植糯玉米[2]。目前的研究普遍认为玉米起源于美洲, 野生玉米被驯化为普通玉米后传入中国, 其第9染色体短臂上的Wx基因发生隐性突变, 形成了一种籽粒干燥后胚乳呈角质不透明、无光泽的蜡质状突变体, 蒸煮后呈糯性, 因此, 被我国喜糯性食物的人们经人工选择保留下来, 称为糯玉米[3]。
虽然, 糯玉米起源于中国, 但是, 关于糯玉米种质资源方面的研究起步较晚, 对糯玉米种质资源的聚类分析主要集中在其遗传多样性。随着生物技术的快速发展, 糯玉米遗传多样性分析方法已突破了形态学、细胞学、同工酶标记方法的局限, 进入了分子生物学研究阶段。目前, 分子标记技术(RFLP、RAPD、SSR、SNP等)因其多态性高、稳定性好、操作简单等特点, 已被广泛应用于糯玉米的遗传多样性分析。刘丽君等[4]利用93对SSR标记研究了55份糯玉米自交系的遗传多样性, 基于UPGMA方法供试自交系分为4类, 划分结果基本符合品系的来源情况, 发现江苏沿江地区糯玉米种质资源主要由通系5群、衡白522群以及突变体材料组成。吴斌等[5]利用SSR标记基于UPGMA方法将36份西双版纳的小糯玉米地方品种聚为6个类群, 地方品种独立成群。Zheng等[6]利用SSR标记基于UPGMA方法将165份糯玉米材料聚类为3个类群。目前, 基于UPGMA方法对糯玉米种质的遗传多样性研究较多, 但是, 对基于模型的糯玉米群体遗传结构的分析还未见报道。本研究基于SSR标记对目前在育种中使用的87份糯玉米自交系进行了遗传多样性和群体遗传结构分析, 确定了糯玉米种质间的遗传背景差异, 为糯玉米优良品种选育和杂种优势模式构建提供了研究基础和理论依据。
1 材料与方法
1.1 试验材料
87份糯玉米自交系(N1~N87)和4份糯玉米杂交种(N88~N91)分别于2016年春、2016年秋和2017年春种植于上海市农业科学院庄行综合试验站(附表1), 行长4.0 m, 行距0.6 m, 株距0.25 m, 田间管理同一般生产田, 以上材料由上海市农业科学院玉米遗传育种课题组提供。Supplementary table 1
附表1
附表191份糯玉米种质名称及系谱来源
Supplementary table 1
| 编号 No. | 名称 Name | 系谱 Pedigree | 来源 Origin | |
|---|---|---|---|---|
| N1 | 16SW-122 | W97/玉美头601♀ W97/Yumeitou 601♀ | 上海 Shanghai | |
| N2 | W97 | 不详 Unknown | 上海 Shanghai | |
| N3 | W148 | 不详 Unknown | 上海Shanghai | |
| N4 | 16SW-292 | W75/W95 | 上海Shanghai | |
| N5 | 16SW-296 | (江南花糯二环系/白478)/W22 (Second cycle lines from Jiangnanhuanuo/Bai 478) /W22 | 江苏 Jiangsu | |
| N6 | 通系5 Tongxi 5 | 不详 Unknown | 江苏 Jiangsu | |
| N7 | 16SW-311 | 申W22/H54 Shen W22/H54 | 上海 Shanghai | |
| N8 | 16SW-312 | 京W♂/W100 JingW♂/W100 | 北京 Beijing | |
| N9 | 16SW-320 | W82/(W88/紫黑W1号) W82/(W88/Zihei W1) | 上海 Shanghai | |
| N10 | 16SW-321 | 申W22/(南汇黑/申W13) Shen W22/(Nanhuihei/Shen W13) | 上海 Shanghai | |
| N11 | 16SW-322 | W96/张红 W96 / Zhanghong | 甘肃 Gansu | |
| N12 | 16SW-324 | W75/[江南花糯二环系/(祝西/5003)] W75/[Second cycle lines from Jiangnanhuanuo/(Zhuxi/5003)] | 江苏 Jiangsu | |
| N13 | 16SW-327 | W75/玉美头601♀ W75/Yumeitou 601♀ | 广西 Guangxi | |
| N14 | 京W♂ Jing W♂ | 不详 Unknown | 北京 Beijing | |
| N15 | 16SW-333 | 申W48/张红 Shen W48/Zhanghong | 甘肃 Gansu | |
| N16 | W153 | 不详 Unknown | 上海 Shanghai | |
| N17 | W152 | 不详 Unknown | 上海 Shanghai | |
| N18 | W158 | 不详 Unknown | 上海 Shanghai | |
| N19 | 16SW-347 | W93/京W♂ W93/Jing W♂ | 北京 Beijing | |
| N20 | 16SW-349 | 申W22/H88 Shen W22/H88 | 上海 Shanghai | |
| N21 | 16SW-350 | 京W♂/H54 JingW♂/H54 | 北京 Beijing | |
| N22 | 16SW-363 | (江南花糯二环系/衡白522)/(江南花糯二环系/G478) (Second cycle lines from Jiangnanhuanuo/Hengbai 522)/(Second cycle lines from Jiangnanhuanuo/G478) | 江苏 Jiangsu | |
| N23 | 16SW-365 | 美玉8号二环系 Second cycle lines from Meiyu8 | 广西 Guangxi | |
| N24 | 16SW-367 | (通系5/昌7-2)/W88 (Tongxi 5/Chang 7-2)/W88 | 不详 Unknown | |
| N25 | 16SW-369 | [江南花糯二环系/(祝西/5003)]/(江南花糯二环系/衡白522) [Second cycle lines from Jiangnanhuanuo/(Zhuxi/5003)]/(Second cycle lines from Jiangnanhuanuo/Hengbai 522) | 江苏 Jiangsu | |
| N26 | 16SW-373 | 07东N-1/玉美头601♀ 07DongN-1/Yumeitou 601♀ | 广西 Guangxi | |
| N27 | 16SW-374 | W95/玉美头601♀ W95/Yumeitou 601♀ | 广西 Guangxi | |
| N28 | 16SW-424 | W93/西星黑糯1号二环系 W93/Second cycle lines from Xixingheinuo 1 | 山东 Shandong | |
| N29 | W116 | 不详 Unknown | 上海 Shanghai | |
| N30 | 16SW-426 | W88/申W22 W88/Shen W22 | 上海 Shanghai | |
| N31 | 16SW-427 | W75/[(W01/申W48)/W01] W75/[(W01/Shen W48)/W01] | 上海 Shanghai | |
| N32 | 16SW-429 | W01/中糯1号OP W01/Zhongnuo 1 OP | 北京 Beijing | |
| N33 | 16SW-437 | 苏玉糯1号二环系 Second cycle lines from Suyunuo1 | 江苏 Jiangsu | |
| N34 | 16SW-438 | [通系5/(515/小黄糯)]/江南花糯二环系 [Tongxi 5/(515/Xiaohuangnuo)]/ Second cycle lines from Jiangnanhuanuo | 云南 Yunnan | |
| N35 | 16SW-439 | [通系5/(515/小黄糯)]/(申W13/通系5) [Tongxi 5/(515/ Xiaohuangnuo)]/(Shen W13/Tongxi 5) | 云南 Yunnan | |
| 编号 No. | 名称 Name | 系谱 Pedigree | 来源 Origin | |
| N36 | 16SW-441 | 申W22/小黄糯 Shen W22/ Xiaohuangnuo | 云南 Yunnan | |
| N37 | 16SW-442 | D049/杂1-4 D049/Za1-4 | 上海 Shanghai | |
| N38 | 16SW-444 | [通系5/(515/小黄糯)]/通系5 [Tongxi 5/(515/Xiaohuangnuo)]/Tongxi 5 | 云南 Yunnan | |
| N39 | 16SW-447 | 申W13/W27 Shen W13/W27 | 上海 Shanghai | |
| N40 | 16SW-449 | (W01/申W13)/申W22 (W01/Shen W13)/Shen W22 | 上海 Shanghai | |
| N41 | 16SW-450 | (申W48/紫黑1号)/[(申W48/W01)/申W48] (Shen W48/Zihei 1)/[(Shen W48/W01)/Shen W48] | 上海 Shanghai | |
| N42 | 16SW-451 | 文山红糯/申W22 Wenshanhongnuo/Shen W22 | 云南 Yunnan | |
| N43 | 16SW-452 | {[通系5/(515/小黄糯)]/通系5}/江南花糯二环系 {[Tongxi 5/(515/Xiaohuangnuo)]/Tongxi 5}/Second cycle lines from Jiangnanhuanuo | 云南 Yunnan | |
| N44 | 16SW-455 | (W01/申W48)/W01 (W01/Shen W48)/W01 | 上海 Shanghai | |
| N45 | 16SW-456 | (申W13/通系5)/{[通系5/(通系5/515)]/通系5} (Shen W13/Tongxi 5)/{[Tongxi 5/(Tongxi 5/515)]/Tongxi 5} | 上海 Shanghai | |
| N46 | 16SW-457 | (W01/申W13)/W01 (W01/Shen W13)/W01 | 上海 Shanghai | |
| N47 | 16SW-458 | 田白糯1号二环系 Second cycle lines from Tianbainuo1 | 广东 Guangdong | |
| N48 | 16SW-459 | (江南花糯二环系/白478)/申W48 (Second cycle lines from Jiangnanhuanuo /Bai 478)/Shen W48 | 江苏 Jiangsu | |
| N49 | 16SW-460 | 申W48/郑白糯6号二环系 Shen W48/Second cycle lines from Zhengbainuo6 | 河南 Henan | |
| N50 | 16SW-465 | [(七宝紫/申W22)/江南花糯二环系]/[江南花糯二环系/(祝西/5003)] [(Qibaozi/Shen W22)/Second cycle lines from Jiangnanhuanuo]/[Second cycle lines from Jiangnanhuanuo/(Zhuxi/5003)] | 上海 Shanghai | |
| N51 | 16SW-466 | {[通系5/(515/小黄糯)]/通系5}/[江南花糯二环系/(祝西/5003))] {[Tongxi 5/(515/Xiaohuangnuo)]/Tongxi 5}/[Second cycle lines from Jiangnanhuanuo/(Zhuxi/5003)] | 江苏 Jiangsu | |
| N52 | 16SW-468 | {{[通5/(515/小黄糯)]/通系5}/江南花糯二环系}/[江南花糯二环系/(祝西/5003)] {{[Tongxi 5/(515/Xiaohuangnuo)]/Tongxi5}/Second cycle lines from Jiangnanhuanuo}/[Second cycle lines from Jiangnanhuanuo/(Zhuxi/5003)] | 江苏 Jiangsu | |
| N53 | 16SW-470 | W88/W96 | 上海 Shanghai | |
| N54 | 16SW-471 | W88/[江南花糯二环系/(祝西/5003)] W88/[Second cycle lines from Jiangnanhuanuo/(Zhuxi/5003)] | 江苏 Jiangsu | |
| N55 | 16SW-475 | [W97/(七宝紫/申W22)]/{[通系5/(515/小黄糯)]/通系5} [W97/(Qibaozi/Shen W22)]/{[Tongxi 5/(515/Xiaohuangnuo)]/Tongxi 5} | 上海 Shanghai | |
| N56 | 16SW-476 | [(七宝紫/申W22)/江南花糯二环系]/{[通系5/(515/小黄糯)]/通系5} [(Qibaozi/Shen W22)/Second cycle lines from Jiangnanhuanuo]/{[Tongxi 5/(515/ Xiaohuangnuo)]/Tongxi 5} | 上海 Shanghai | |
| N57 | 16SW-478 | W01/京W♂ W01/JingW♂ | 北京 Beijing | |
| N58 | 16SW-480 | [通系5(488/5003)]/W88 [Tongxi 5(488/5003)]/W88 | 上海 Shanghai | |
| N59 | 16SW-481 | [(申W13/通系5)/昌7-2]/W88 [(Shen W13/Tongxi 5)/Chang 7-2]/W88 | 江苏 Jiangsu | |
| N60 | 16SW-484 | (文山红糯/申W22)/申W22 (Wenshanhongnuo/Shen W22)/Shen W22 | 云南 Yunnan | |
| N61 | 16SW-485 | W75/白478 W75/Bai 478 | 山东 Shandong | |
| N62 | 16SW-487 | 申W13/(江南花糯二环系/白478) Shen W13/(Second cycle lines from Jiangnanhuanuo/Bai 478) | 江苏 Jiangsu | |
| N63 | W108 | 不详 Unknown | 上海 Shanghai | |
| N64 | W107 | 不详 Unknown | 上海 Shanghai | |
| N65 | W01 | 不详 Unknown | 上海 Shanghai | |
| N66 | W75 | 不详 Unknown | 上海 Shanghai | |
| 编号 No. | 名称 Name | 系谱 Pedigree | 来源 Origin | |
| N67 | W151 | 不详 Unknown | 上海 Shanghai | |
| N68 | W114 | 不详 Unknown | 上海 Shanghai | |
| N69 | 张改♀ Zhanggai ♀ | 不详 Unknown | 上海 Shanghai | |
| N70 | wh-2 | 不详 Unknown | 上海 Shanghai | |
| N71 | 16HW-31 | (江南花糯二环系/白478)/W95 (Second cycle lines from Jiangnanhuanuo/Bai 478)/W95 | 山东 Shandong | |
| N72 | 16HW-46 | (W96/张红)/申W22 (W96/Zhanghong)/Shen W22 | 甘肃 Gansu | |
| N73 | 16AW-1001 | 引种大/W95 Yinzhongda/W95 | 上海 Shanghai | |
| N74 | 16AW-1003 | 申W48/[(申W48/紫黑1号)/申W48] Shen W48/[(Shen W48/Zihei 1)/Shen W48] | 上海 Shanghai | |
| N75 | 16AW-1004 | [(申W48/W01)/申W48]/[(申W48/紫黑1号)/申W48] [(Shen W48/W01)/Shen W48]/[(Shen W48/Zihei 1)/Shen W48] | 上海 Shanghai | |
| N76 | 16AW-1005 | [(江南花糯二环系/白478)/郑白糯6号二环系]/京W♂ [(Second cycle lines from Jiangnanhuanuo/Bai 478)/Second cycle lines from Zhengbainuo6]/Jing W♂ | 河南 Henan | |
| N77 | 16AW-1006 | [(W48/5003)/W48]/中糯2号二环系 [(W48/5003)/W48]/Second cycle lines from Zhongnuo 2 | 北京 Beijing | |
| N78 | 查12at-1385 Cha 12at-1385 | 不详 Unknown | 上海 Shanghai | |
| N79 | 16AW-1008 | 张红/[江南花糯二环系/(祝西/5003)] Zhanghong/[Second cycle lines from Jiangnanhuanuo/(Zhuxi/5003)] | 江苏 Jiangsu | |
| N80 | 09N-5M | 不详 Unknown | 上海 Shanghai | |
| N81 | 09N-6M | 不详 Unknown | 上海 Shanghai | |
| N82 | 09N-S150P | 不详 Unknown | 上海 Shanghai | |
| N83 | 16AW-1015 | [(申W48/紫黑1号)/(申W48/W01)]/申W22 [(Shen W48/Zihei 1)/(Shen W48/W01)]/Shen W22 | 上海 Shanghai | |
| N84 | 16AW-1019 | 申W48/郑白糯6号二环系 Shen W48/Second cycle lines from Zhengbainuo 6 | 河南 Henan | |
| N85 | W-109 | 不详 Unknown | 上海 Shanghai | |
| N86 | 申W93 Shen W93 | 荆黑糯2号二环系 Second cycle lines from Jingheinuo 2 | 上海 Shanghai | |
| N87 | 黄W-♀ Huang W- ♀ | 不详 Unknown | 上海 Shanghai | |
| N88 | 16HW-LY5 | 16HW-31/申W48 16HW-31/Shen W48 | 上海 Shanghai | |
| N89 | 16HW-LY8 | 16HW-4/16HW-46 | 上海 Shanghai | |
| N90 | 沪紫黑糯2号 Huziheinuo 2 | 申W74/申W93 Shen W74/Shen W93 | 上海 Shanghai | |
| N91 | 五彩甜糯2号 Wucaitiannuo 2 | 申W93/wh-2 Shen W93/wh-2 | 上海 Shanghai | |
新窗口打开|下载CSV
1.2 玉米基因组DNA提取
苗期于田间采集以上91份糯玉米材料幼嫩叶片, 用液氮研磨后, 利用植物基因组DNA提取试剂盒以改进后的CTAB法提取DNA[7]。试剂盒购于北京庄盟国际生物基因科技有限公司。1.3 SSR引物筛选与PCR扩增
根据中华人民共和国农业行业标准(NY/T1432- 2014玉米品种鉴定技术规程SSR标记法)提供的40对核心引物序列和PCR扩增程序对供试的91份材料进行基因型鉴定[8,9]。此外, 根据玉米基因组数据库(Maize Genome Database, https://maizegdb.org/)公布的玉米SSR引物信息, 在玉米第1~第10染色体上取定位于不同染色体bin的SSR标记各10个鉴定基因型, 采用Lu等[10]的PCR扩增和产物鉴定程序。1.4 数据统计与分析
根据SSR扩增产物在电泳凝胶上的相对位置记录基因型, 在电泳凝胶上相同的位置有带记为“1”, 无带记为“0”, 带弱或在材料中无扩增条带的记为“9”。利用PowerMarker 3.25分析等位基因数、主等位基因频率(major allele frequency, MAF)、基因多样性(gene diversity, H)以及多态性信息含量(polymorphism information content, PIC)。MAF=检测位点最多基因型的个数/检测位点总基因型个数。$H=1-\sum{_{j=1}^{m}r_{j}^{2}}$, 其中m表示该位点上的等位基因数, rj为该位点上第j个等位基因在总居群中的平均频率。$\text{PIC}=1-\sum{_{j}^{i}p_{ij}^{2}}$, 式中p表示位点i的第j个等位变异出现的频率。利用NTSYS 2.10软件根据Nei等[11]的方法计算样品间遗传相似系数(genetic similarity, GS), $\text{GS}=2{{N}_{ij}}/({{N}_{i}}+{{N}_{j}})$, 其中Ni为i样品出现的谱带数, Nj为j样品出现的谱带数, Nij为i样品和j样品共有的谱带数。利用MEGA 7.0[12]中非加权配对算术平均法(unweighted pair-group method with arithmetic means, UPGMA)构建聚类分析树状图。使用Structure 2.3.4分析群体遗传结构, 群体数目(K)为1~10, 对每个K值模拟运算7次, 设不作数迭代(length of burn-in period)开始时的MCMC (markov chain monte carlo)为50,000次, 不作数迭代后的MCMC为100,000次, 计算Q值。当似然值随K值的增加而增大时, 利用ΔK的方法确定最适K值[13], $Delta K=m\left( \left[ \text{L}''(K) \right] \right)/S\left[ \text{L}'(K) \right],$其中$\text{ln}\ P(D)=L(K),$ $\text{L}'(K)=\text{L}(K)-\text{L}(K-1)$,$\left[ \text{L}''(K) \right]=[\text{L}'(K+1)-\text{L}'(K)]$。2 结果与分析
2.1 SSR标记分布及多态性分析
从140对SSR标记中共筛选出29对扩增稳定、条带差异明显的引物用于对87份糯玉米自交系的基因型分析, 共检测到180个等位基因, 变幅为4~20, 每个标记平均检测到6个等位基因(表1)。87份糯玉米自交系的平均基因多样性为0.619, 变幅为0.323~ 0.921。29对标记的PIC变幅为0.308~0.915, 平均为0.572; 其中, 20对标记的PIC ≥ 0.50 (69%), 属于高度多态性位点; 其余9对标记0.250 < PIC < 0.50 (31%), 属于中度多态性位点。综上结果说明这29对SSR标记多态性较高, 能较好地满足遗传多样性分析需求。Table 1
表1
表129对SSR标记的多态性参数
Table 1
| 编号 No. | 标记位点 Locus | Bin | 等位基因数 No. of alleles | 主等位基因频率 Major allele frequency | 基因多样性 Gene diversity | 多态性信息含量 Polymorphic information content |
|---|---|---|---|---|---|---|
| 1 | umc1147y4 | 1.07 | 4 | 0.54 | 0.575 | 0.496 |
| 2 | bnlg1671y17 | 1.10 | 4 | 0.60 | 0.579 | 0.531 |
| 3 | phi96100y1 | 2.00 | 4 | 0.67 | 0.504 | 0.458 |
| 4 | umc1823 | 2.02 | 4 | 0.45 | 0.640 | 0.571 |
| 5 | b1 | 2.03 | 6 | 0.39 | 0.743 | 0.705 |
| 6 | zpu1 | 2.05 | 4 | 0.40 | 0.663 | 0.599 |
| 7 | pbf1 | 2.05 | 4 | 0.80 | 0.333 | 0.308 |
| 8 | umc2253 | 2.05 | 4 | 0.59 | 0.554 | 0.483 |
| 9 | umc1875 | 2.06 | 4 | 0.53 | 0.570 | 0.484 |
| 10 | umc1536k9 | 2.07 | 4 | 0.78 | 0.358 | 0.320 |
| 11 | bnlg1520K1 | 2.09 | 7 | 0.39 | 0.722 | 0.675 |
| 12 | umc2105k3 | 3.00 | 4 | 0.75 | 0.412 | 0.379 |
| 13 | umc1746 | 3.00 | 4 | 0.76 | 0.402 | 0.375 |
| 14 | umc2257 | 3.01 | 4 | 0.53 | 0.590 | 0.515 |
| 15 | umc1495 | 3.04 | 13 | 0.26 | 0.843 | 0.826 |
| 16 | umc2264 | 3.04 | 7 | 0.34 | 0.769 | 0.734 |
| 17 | phi053k2 | 3.05 | 8 | 0.43 | 0.734 | 0.699 |
| 18 | phys2 | 3.05 | 20 | 0.13 | 0.921 | 0.915 |
| 19 | umc1027 | 3.06 | 5 | 0.82 | 0.323 | 0.308 |
| 20 | umc1489y3 | 3.07 | 7 | 0.46 | 0.668 | 0.611 |
| 21 | mtl1 | 4.01 | 4 | 0.45 | 0.650 | 0.580 |
| 22 | bnlg490y4 | 4.04 | 4 | 0.57 | 0.591 | 0.536 |
| 23 | umc1705w1 | 5.03 | 4 | 0.52 | 0.620 | 0.553 |
| 24 | bnlg2305k4 | 5.07 | 5 | 0.51 | 0.631 | 0.569 |
| 25 | bnlg161k8 | 6.00 | 8 | 0.34 | 0.786 | 0.758 |
| 26 | bnlg1702k1 | 6.05 | 8 | 0.55 | 0.629 | 0.588 |
| 27 | phi065k9 | 9.03 | 5 | 0.48 | 0.599 | 0.519 |
| 28 | umc1492y13 | 9.04 | 6 | 0.53 | 0.647 | 0.602 |
| 29 | umc1432y6 | 10.02 | 15 | 0.20 | 0.889 | 0.879 |
| 平均值 Mean | 6 | 0.51 | 0.619 | 0.572 |
新窗口打开|下载CSV
2.2 遗传多样性分析
糯玉米单交种“沪紫黑糯2号”与亲本申W93间的GS为0.80; “五彩甜糯2号”与亲本申W93和wh-2间的GS分别为0.67和0.64; 16HW-LY5与其亲本16HW-31间的GS为0.63; 16HW-LY8与其亲本16HW-46间的GS为0.68。综上结果可见, 供试单交种与其亲本间的GS变幅为0.63~0.80, 亲缘关系较近, GS结果与系谱一致, 说明本研究对糯玉米种质间遗传多样性分析结果合理可靠。16SW-292和16AW-363之间的GS最小(0.49), 16SW-466和16SW-468之间的GS最大(0.93)。根据系谱信息16SW-292和16AW-363选育于不同亲本, 亲缘关系较远; 16SW-466和16SW-468选自于相同亲本的不同组合, 亲缘关系很近(表1)。87份糯玉米自交系间两两计算共获得3741个GS, 变幅为0.49~0.93, 平均为0.66。由图1可见, 87份糯玉米自交系间的GS呈单峰分布, 85.40%的自交系间GS在0.60~0.80之间, 91.29%的自交系间GS ≤ 0.75, 说明本研究供试糯玉米自交系间遗传基础较丰富, 适于进行遗传多样性分析。
图1
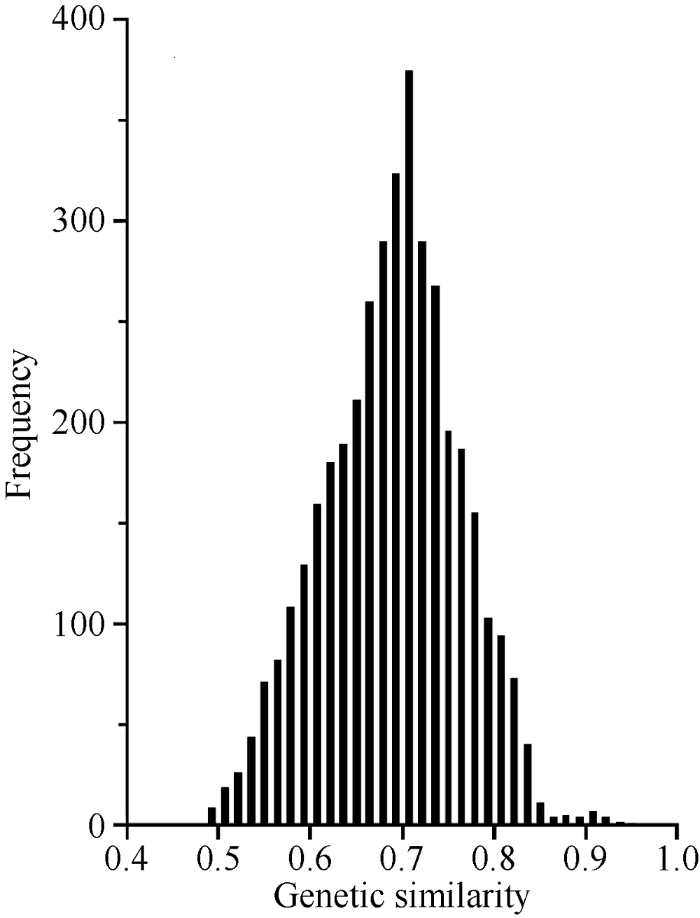 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1糯玉米自交系间GS频次分布图
Fig. 1Frequency distributions of GS between waxy maize inbred lines
2.3 聚类分析
聚类树状图表明87份糯玉米自交系在GS为0.64处可划分为四大类(图2)。图2
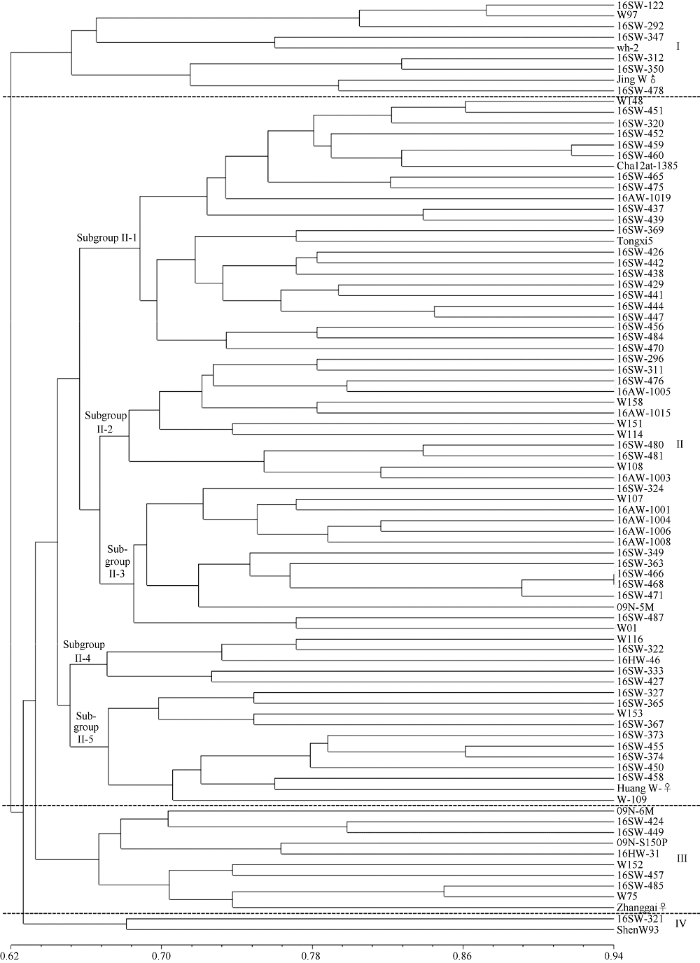 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图287份糯玉米自交系基于SSR标记的聚类分析图
Fig. 2Cluster analysis of 87 waxy maize inbred lines by SSR markers
类群I包括wh-2、京W♂和W97等来自北京和上海的共9份糯玉米自交系, 自交系间GS为0.59~0.87, 平均为0.69。根据系谱信息发现, 自交系京W♂、以及从以京W♂为亲本组配的杂交种中选育的二环系均被聚类于类群I, 京W♂与类群I内其他糯玉米自交系间的平均GS为0.69, 推测类群I材料与京W♂亲缘关系较近。
类群Ⅱ包括通系5、W158、09N-5M和16SW-365等66份糯玉米自交系, 材料间的GS为0.49~0.93, 平均为0.68。其中, 16SW-451和16SW-468间的GS最小, 为0.49; 16SW-466和16SW-468间的GS最大, 为0.93。此外, 类群II在GS为0.67处又可划分为5个亚类。亚群II-1包含通系5、W148和16SW-437等24份自交系, 它们之间的GS平均为0.72。其中, 包括糯玉米自交系通系5、以及以通系5为亲本杂交后选育的16SW-456、16SW-438、16SW-444等6份自交系, 通系5与亚群II-1内其他糯玉米自交系间的平均GS为0.69, 推测亚群II-1自交系与通系5亲缘关系较近。亚群II-2包括W151、W158、16SW-311等12份糯玉米自交系, 它们之间的GS平均为0.71; 其中除了来自河南的自交系16AW-1005外, 其余自交系均来源于上海和江苏。亚群II-3包括16SW-466、W-107、09N-5M等14份糯玉米自交系, 它们之间的GS平均为0.73; 其中除了来自河南的自交系16AW-1006外, 其余自交系均来源于上海和江苏。亚群II-4包括来自甘肃和上海的W116、16SW-322、16SW-46、16SW-333、16SW-427共5份糯玉米自交系, 它们之间的GS平均为0.65; 亚群II-4包括全部来自甘肃的自交系16SW-322、16SW-333和16HW-46, 它们与II-4亚群内来自上海的自交系W116和16SW-427间的平均GS分别为0.64、0.65和0.64, 推测W116和16SW-427与甘肃地区种质亲缘关系较近。亚群II-5包括黄W-♀、W153、16SW-365、16SW-373等来自广西、上海和广东的共11份糯玉米自交系, 它们之间的GS平均为0.71; 其中来自广西的16SW-327、16SW-365、16SW-373和16SW-374共4份糯玉米自交系, 它们与亚群II-5内其他自交系间的GS分别为0.67、0.73、0.74和0.72, 推测亚群II-5的自交系与广西糯玉米种质亲缘关系较近。
类群III包括09N-6M、W75、W152等10份糯玉米自交系, 它们之间的GS为0.57~0.85, 平均为0.70。类群III包括全部来自山东的自交系16SW-424、16SW-485和16HW-31, 它们与类群III其他自交系间的GS分别为0.75、0.75和0.71, 推测类群III自交系与山东地区种质亲缘关系较近。
来源于上海的糯玉米自交系16SW-321和申W93与其他材料间的GS平均分别为0.63和0.64, 遗传差异较大, 因此, 16SW-321和申W93被单独聚类为类群IV。
2.4 群体遗传结构分析
利用Structure软件分析发现, 在1~10范围内随着K值的增大ln P(D)值也随之增大(附图1)。因此, 依据ΔK的方法进一步确定类群数, 发现在K = 4时ΔK值出现显著的峰值(图3-A), 据此推断87份糯玉米自交系可划分为4个类群, 分别包含24、25、19和19份糯玉米自交系(图3-B)。附图1
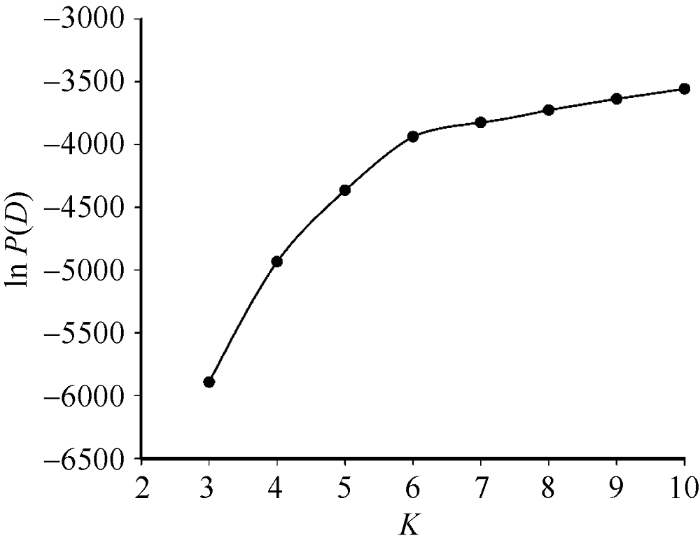 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附图1ln P(D)随K值的变化
Supplementary fig. 1The line chart of ln P(D) value changing with K-value
图3
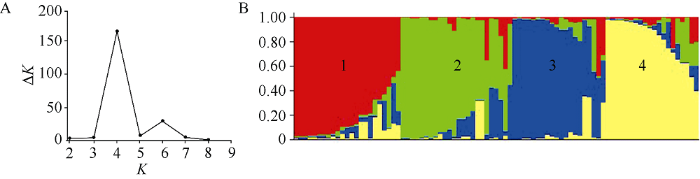 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图387份糯玉米自交系的群体遗传分析图
A: ΔK随K值的变化趋势; B: K = 4时, 以Structure运算获得的87份糯玉米自交系的群体遗传结构。
Fig. 3Population genetic analysis of 87 waxy maize inbred lines
A: the line chart of ΔK changing with K-value; B: the population structure of 87 waxy maize inbred lines estimated by Structure (K = 4).
利用自交系Q值可以分析不同类群在各自交系中所占成分比例。根据最大Q值分布, 87份糯玉米自交系中有15份Q值< 0.6, 占自交系总数的17.24%, 然而Q > 0.8和0.9的自交系分别占64.37%和47.13%, 说明该群体大部分材料的血缘关系较单一。
3 讨论
3.1 糯玉米种质遗传多样性分析
一直以来, 搜集种质资源、拓宽遗传基础、对种质资源开展科学的鉴定与评价是选育优良品种的基础, 在品种改良中始终占据重要地位[14,15]。本研究利用29对SSR标记对87份目前育种中常用的糯玉米自交系进行遗传多样性分析, GS取值范围为0.49~0.93, 平均为0.66, 在GS为0.64处将供试群体划分为四大类。类群I包含9份糯玉米自交系, 其中包括京W♂、以及所有以京W♂为亲本选育的4份自交系。京W♂与类群I内其他糯玉米自交系间的平均GS为0.69, 亲缘关系较近, 推测16SW-122、W97、wh-2和16SW-292可能是北京种质京W♂在上海的改良自交系。类群Ⅱ包含66份自交系, 包括“苏玉糯1号”的二环系16SW-437, 以“江南花糯”二环系为亲本选育的自交系16SW-296、16SW-324、16SW-363、16SW-369、16SW-438、16SW-452、16SW-459、16SW-465、16SW-466、16SW-468、16SW-471、16SW-476、16SW-487、16AW-1005、16AW-1008共15份材料, “美玉8号”的二环系16SW-365, “田白糯1号”的二环系16SW-458。“美玉8号”的母本是“苏玉糯1号”的二环系, 父本是衡白522与广西桂林农家种白糯玉米杂交选育的二环系[16]; “田白糯1号”母本是“苏玉糯2号”与“中糯1号”杂交选育的二环系, 父本选自广西河池农家种[17]; “苏玉糯2号”与“苏玉糯1号”的双亲都含有通系5和衡白522, 因此亲缘关系较近[18]。根据系谱信息, “美玉8号”、“田白糯1号”和“苏玉糯1号”的亲缘关系较近, 可见本研究结果与系谱信息一致。“苏玉糯1号”是由通系5与衡白522杂交育成的国审糯玉米品种, 具有高产、稳产性、品质较好、多抗的特点, 用其选育的二环系已在育种中大量使用, 组配出了“郑白糯4号”、“南农紫玉糯1号”、“山农202”等一批优质糯玉米品种[19]。“江南花糯”是由“苏玉糯1号”的二环系与农家种组配的杂交种, 具有产量高、肉质厚、糯性强、香味浓、外观美的特点[20]。类群II中16SW-460、16AW-1019和16AW-1005是以“郑白糯6号”的二环系为亲本选育的自交系。国审糯玉米品种“郑白糯6号”表现中矮秆、抗倒、抗病、产量高、结实性好、穗匀、品质好[21]。此外, 通系5和以通系5为亲本选育的12份自交系也被聚类于类群II。通系5农艺性状优良、糯性好、配合力高, 通系5或衍生品种已被国内多个育种单位作为种质资源广泛应用, 育成了28个优良的鲜食糯玉米品种。根据系谱信息可见, 类群II的自交系系谱来源品质优异, 推测类群II的自交系可作为骨干亲本用于今后的优质糯玉米选育。
类群IV只包含16SW-321和申W93两份材料, 其中申W93是“荆黑糯2号”的二环系。“荆黑糯2号”来源于湖北,据此推测类群Ⅳ种质与湖北糯玉米种质亲缘关系较近。
我国是公认的糯玉米起源中心, 据统计我国79.55%的糯玉米种质分布于西南地区(云南、广西、贵州、四川)[22]。本研究供试群体中包括来自北京、甘肃、河南、山东、江苏、上海、广西、广东和云南的糯玉米种质, 结合系谱信息和聚类结果可见, 大部分材料的亲缘关系与其地理来源相关; 而少部分材料推测是由于不同区域品种间进行了频繁的基因交流和重组, 遗传基础组成丰富, 失去了明显的区域特征, 导致聚类结果与地理来源信息不符。
综上结果说明目前我国糯玉米育种工作仍面临着亲本遗传基础狭窄的困境, 亟须加强对不同地区糯玉米种质资源的引进和创制, 为选育具有突破性的优质糯玉米新品种提供物质基础。
3.2 遗传相似系数聚类和Structure模型聚类结果比较
目前, 生产中应用的糯玉米自交系多来源于优质杂交种或中间群体, 很多材料已无系谱可查或遗传基础复杂, 无法利用系谱追踪法研究它们的亲缘关系。分子标记技术不受环境和季节限制、标记覆盖全基因组、多态性高、成本较低, 已成为遗传多样性分析最常用的技术。种质聚类方法有多种, 包括遗传相似系数法、遗传距离法、模型聚类法, 选择不同的参数会导致分析结果较大差异[23]。本研究通过GS将87份糯玉米自交系划分为4个类群, 分别包括9、66、10和2份材料; 而基于Structure模型的群体遗传结构分析也将这些材料分为4个居群, 分别包括24、25、19和19份材料。可见利用UPGMA方法和Structure模型聚类法划分的类群数一致, 说明本研究对群体类群的划分结果合理可靠。基于遗传相似系数的聚类分析是根据材料间亲缘关系的远近, 类群划分时选择的GS水平受人为主观判定影响; 而群体遗传结构分析以亚群是否达到Hardy-Weinberg平衡建立的数学模型, 通过计算材料相应的Q值分析其组成成分来进行聚类划分。刘秀云等[24]通过UPGMA聚类分析将255个枣品种分为15个亚类; 同时, 利用群体结构分析也将这些材料划分为15个群体, 与聚类分析结果基本一致。王晋等[25]认为Structure软件的群体遗传结构分析可以避免人为因素对类群划分的影响, 进而对供试材料的群体结构进行校正。赵旭等[26]认为Structure的模型分析根据ΔK峰值确定亚群间遗传分化不明显的复杂群体的类群数[13,27], 模型聚类方法可以将遗传距离聚类中系谱关系不明确的自交系划分到相应类群, Structure的模型聚类法更有利于研究玉米自交系的遗传关系, 将类群划分更细致; 而结合遗传相似系数和Structure模型聚类法能更好地确定自交系间的亲缘关系。本研究中虽然利用遗传相似系数聚类和Structure模型聚类划分的类群数相同, 但是各类群的种质组成差异较大, 推测这也是由于所利用的遗传相似系数聚类和Structure模型聚类计算方法不同导致的。因此, 进行种质聚类分析时, 根据种质已知信息, 选择合适的参数和算法对于准确评价群体间遗传多样性及种质的精细聚类分析具有重要意义。
4 结论
基于遗传相似系数, 通过UPGMA方法将87份糯玉米自交系划分为4个类群, 利用Structure软件进行群体遗传结构分析也将该群体聚类为4个居群, 但各类群包含的种质不同。本研究认为目前我们育种中使用的糯玉米自交系遗传变异较单一, 亟须引进和创制遗传基础丰富的种质资源。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.3969/j.issn.1005-0906.2006.03.013URL [本文引用: 1]

糯玉米起源于我国,我国糯玉米种质资源丰富,糯玉米育种起步较晚,但发展迅速.分析了我国糯玉米育种现状,提出了几点育种研究对策:育种工作者应将种质资源工作放在战略位置;合理制定育种目标,确定合适的选育方向;加强新技术应用;育种与成果转化并重,调动育种积极性.
DOI:10.3969/j.issn.1005-0906.2006.03.013URL [本文引用: 1]
糯玉米起源于我国,我国糯玉米种质资源丰富,糯玉米育种起步较晚,但发展迅速.分析了我国糯玉米育种现状,提出了几点育种研究对策:育种工作者应将种质资源工作放在战略位置;合理制定育种目标,确定合适的选育方向;加强新技术应用;育种与成果转化并重,调动育种积极性.
DOI:10.3969/j.issn.1008-2239.2010.03.024URL [本文引用: 1]
分析了糯玉米的起源,探讨了当前研究现状,阐述了生产发展概况。
DOI:10.3969/j.issn.1008-2239.2010.03.024URL [本文引用: 1]
分析了糯玉米的起源,探讨了当前研究现状,阐述了生产发展概况。
URL [本文引用: 1]
现代作物一般经历驯化和随后的育种选择两次人工选择过程。玉米驯化起源于美洲,中国糯玉米是在美洲驯化玉米基础上经近代育种选择而成,因此是研究玉米和糯玉米遗传改良育种分子选择机制的独特材料。本论文调查了325份糯玉米的waxy(wx)基因型及主要基因型的地理分布,然后分别对源自独立起源的wx基因型群体做了遗传多样性调查和淀粉代谢途径的6个关键基因(wx,sh1,sh2,su1,ea1,bt2)的育种选择的统计检验,获得如下主要结果: wx基因型调查发现,325份糯玉米材料中315份(96.6%)属于wx-D7(D7)或wx-D10(D10)突变基因型(其中55份(88.8%)商业杂交种...
URL [本文引用: 1]
现代作物一般经历驯化和随后的育种选择两次人工选择过程。玉米驯化起源于美洲,中国糯玉米是在美洲驯化玉米基础上经近代育种选择而成,因此是研究玉米和糯玉米遗传改良育种分子选择机制的独特材料。本论文调查了325份糯玉米的waxy(wx)基因型及主要基因型的地理分布,然后分别对源自独立起源的wx基因型群体做了遗传多样性调查和淀粉代谢途径的6个关键基因(wx,sh1,sh2,su1,ea1,bt2)的育种选择的统计检验,获得如下主要结果: wx基因型调查发现,325份糯玉米材料中315份(96.6%)属于wx-D7(D7)或wx-D10(D10)突变基因型(其中55份(88.8%)商业杂交种...
DOI:10.3969/j.issn.1000-4440.2011.04.006URL [本文引用: 1]
为研究江苏沿江地区糯玉米种质的遗传多样性以及江苏省糯玉米的遗传改良和杂种优势利用提供参考,利用93对SSR标记研究55份糯玉米自交系和30份糯玉米单交种的遗传多样性,并利用UPGMA方法对所有自交系材料进行系统聚类。结果表明在自交系中筛选出88对多态性较好的引物,共扩增出350个差异片断,每对引物检测出2~9个差异片段,平均为3.98个;SSR标记的多态性信息量值在0.137与0.832之间,平均为0.520。在单交种中筛选出87对多态性好的引物,扩增出311个差异片断,每对引物可检测出2~9个差异片断,平均3.57个,SSR标记的多态性信息量值在0.064与0.839之间,平均为0.475。利用UPGMA聚类分析方法将所有供试自交系分为4类,划分结果基本符合品系的来源情况,江苏沿江地区糯玉米种质资源主要由通系5群、衡白522群以及突变体材料组成,多样性较为丰富,可为糯玉米遗传改良提供一定的遗传基础。
DOI:10.3969/j.issn.1000-4440.2011.04.006URL [本文引用: 1]
为研究江苏沿江地区糯玉米种质的遗传多样性以及江苏省糯玉米的遗传改良和杂种优势利用提供参考,利用93对SSR标记研究55份糯玉米自交系和30份糯玉米单交种的遗传多样性,并利用UPGMA方法对所有自交系材料进行系统聚类。结果表明在自交系中筛选出88对多态性较好的引物,共扩增出350个差异片断,每对引物检测出2~9个差异片段,平均为3.98个;SSR标记的多态性信息量值在0.137与0.832之间,平均为0.520。在单交种中筛选出87对多态性好的引物,扩增出311个差异片断,每对引物可检测出2~9个差异片断,平均3.57个,SSR标记的多态性信息量值在0.064与0.839之间,平均为0.475。利用UPGMA聚类分析方法将所有供试自交系分为4类,划分结果基本符合品系的来源情况,江苏沿江地区糯玉米种质资源主要由通系5群、衡白522群以及突变体材料组成,多样性较为丰富,可为糯玉米遗传改良提供一定的遗传基础。
URL [本文引用: 1]
利用SSR标记对西双版纳的36个小糯玉米地方品种和2个对照品种进行遗传多样性研究。从800余对SSR引物中筛选出100对多态性好、稳定性高的SSR引物。结果表明,这100对SSR引物在36个糯玉米地方品种中共检测出353个等位基因,每对引物检测出2~8个,平均为3.53个,平均多态性息量(PIC)为0.53,平均标记索引系数(MI)为2.00。聚类分析表明,糯玉米地方品种聚为6个类群,与对照品种相比,地方品种独立成群。
URL [本文引用: 1]
利用SSR标记对西双版纳的36个小糯玉米地方品种和2个对照品种进行遗传多样性研究。从800余对SSR引物中筛选出100对多态性好、稳定性高的SSR引物。结果表明,这100对SSR引物在36个糯玉米地方品种中共检测出353个等位基因,每对引物检测出2~8个,平均为3.53个,平均多态性息量(PIC)为0.53,平均标记索引系数(MI)为2.00。聚类分析表明,糯玉米地方品种聚为6个类群,与对照品种相比,地方品种独立成群。
.
DOI:10.1371/journal.pone.0066606URLPMID:3688585 [本文引用: 1]
Waxy maize (Zea mays L. var. certaina Kulesh), with many excellent characters in terms of starch composition and economic value, has grown in China for a long history and its production has increased dramatically in recent decades. However, the evolution and origin of waxy maize still remains unclear. We studied the genetic diversity of Chinese waxy maize including typical landraces and inbred lines by SSR analysis and the results showed a wide genetic diversity in the Chinese waxy maize germplasm. We analyzed the origin and evolution of waxy maize by sequencing 108 samples, and downloading 52 sequences from GenBank for the waxy locus in a number of accessions from genus Zea. A sharp reduction of nucleotide diversity and significant neutrality tests (Tajima's D and Fu and Li's F*) were observed at the waxy locus in Chinese waxy maize but not in nonglutinous maize. Phylogenetic analysis indicated that Chinese waxy maize originated from the cultivated flint maize and most of the modern waxy maize inbred lines showed a distinct independent origin and evolution process compared with the germplasm from Southwest China. The results indicated that an agronomic trait can be quickly improved to meet production demand by selection.
DOI:10.1093/nar/8.19.4321URL [本文引用: 1]
DOI:10.1016/B978-0-12-385851-1.00015-9URL [本文引用: 1]
Abstract A core set of 60 SSRs was selected and modified using 231 Chinese and USA maize (Zea mays L.) inbred lines from more than 2000 SSRs for DNA fingerprinting analysis. All 60 SSR markers met the following criteria: (1) amplification of a single locus; (2) distinct amplification products; (3) adequate intervals between adjacent alleles; (4) suitable PCR fragment size; (5) reasonable discrimination power (DP); and (6) even distribution across the maize genome. Furthermore, the 60 SSR primers were re-designed to adjust the PCR product size. Together with the application of four different fluorescent dyes, a high-throughput 10-plex capillary electrophoresis platform was explored. The 60 core SSR markers were further divided into three groups (20 SSRs per group) according to peak morphology and DP value. Groups I, II and III were used in DNA fingerprinting analysis as a basic core, an expanded core and a candidate core set respectively. The allele number per locus varied from three to 22 with an average of 8.95; the average number of alleles per group I, II and III was a respective 7.35, 7.8 and 11.4. The DP values ranged from 0.366 to 0.913, with an average of 0.718 among all loci; the average group DP values were 0.697, 0.718 and 0.737 for groups I, II and III respectively; and the cumulative values of discrimination power (CDP) approached 1 for all groups. Cluster analysis results using 60 selected loci divided the Chinese inbred lines into six groups, including Luda Red Cob, P, Improved Reid, Tang-si-ping-tou, Waxy and Lancaster. The USA inbred lines were segregated into four groups, including SSS, Lancaster, Iodent and Oh43/Oh07Mid mixed.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
.
DOI:10.1086/282771URL [本文引用: 1]
DOI:10.1093/molbev/msw054URLPMID:27004904 [本文引用: 1]
Abstract. We present the latest version of the Molecular Evolutionary Genetics Analysis (M ega ) software, which contains many sophisticated methods and tools
DOI:10.1111/mec.2005.14.issue-8URL [本文引用: 2]
DOI:10.3969/j.issn.1672-1810.2003.03.009URL [本文引用: 1]
本文对 2 0个玉米优异地方品种资源主要农艺性状进行了研究 ,运用灰色系统理论的方法 ,分析了资源的综合性状及各性状间的关系。研究结果表明 ,株高影响穗粗 ,穗位高影响穗长 ,株高和穗位均低的资源不会有较高的单株粒重 ;叶片数对资源的筛选、利用有一定的影响 ,对穗长、穗粗、单株粒重有显著影响 ;利用生育期较短的资源可能会选出单株粒重较高的自交系和杂交种。据此结果结合育种目标评价 ,筛选出白头霜、高油玉米、洛阳 85、白鹤、二夏、品综 2号、本地黄玉米、金皇后8个玉米地方品种 ,综合农艺性状好 ,具有较高的利用价值。
DOI:10.3969/j.issn.1672-1810.2003.03.009URL [本文引用: 1]
本文对 2 0个玉米优异地方品种资源主要农艺性状进行了研究 ,运用灰色系统理论的方法 ,分析了资源的综合性状及各性状间的关系。研究结果表明 ,株高影响穗粗 ,穗位高影响穗长 ,株高和穗位均低的资源不会有较高的单株粒重 ;叶片数对资源的筛选、利用有一定的影响 ,对穗长、穗粗、单株粒重有显著影响 ;利用生育期较短的资源可能会选出单株粒重较高的自交系和杂交种。据此结果结合育种目标评价 ,筛选出白头霜、高油玉米、洛阳 85、白鹤、二夏、品综 2号、本地黄玉米、金皇后8个玉米地方品种 ,综合农艺性状好 ,具有较高的利用价值。
URL [本文引用: 1]
正玉米起源于中南美洲,是天然的异花授粉作物,它雌雄同株异花且异位,主要靠风媒传粉,自然群体通常是杂合体,遗传基础丰富,类型多样。玉米传入我国栽培已经有近500年
URL [本文引用: 1]
正玉米起源于中南美洲,是天然的异花授粉作物,它雌雄同株异花且异位,主要靠风媒传粉,自然群体通常是杂合体,遗传基础丰富,类型多样。玉米传入我国栽培已经有近500年
DOI:10.3969/j.issn.1001-7283.2005.05.033URL [本文引用: 1]
加甜糯玉米是介于糯玉米和甜玉米之间的一种新类型特种玉米,在这一新类型特种玉米育种中,北京市率先审定推广了由海南绿川种苗有限公司选育的“都市丽人”加甜糯玉米;湖南、北京、上海也审定推广了由我们选育的美玉(加甜糯)3号。为了进一步提高加甜糯玉米的产量,增进品质,使
DOI:10.3969/j.issn.1001-7283.2005.05.033URL [本文引用: 1]
加甜糯玉米是介于糯玉米和甜玉米之间的一种新类型特种玉米,在这一新类型特种玉米育种中,北京市率先审定推广了由海南绿川种苗有限公司选育的“都市丽人”加甜糯玉米;湖南、北京、上海也审定推广了由我们选育的美玉(加甜糯)3号。为了进一步提高加甜糯玉米的产量,增进品质,使
DOI:10.3969/j.issn.1004-874X.2006.12.006URL [本文引用: 1]
田白糯1号是广州市田园农业科 技研究中心以TMN03-7-5-3-2-1-6作母本、以TMN16-2-4-6-1-7-3作父本选育而成的糯玉米新组合。该组合利用“温带血缘×亚 热带血缘”杂优模式,实现了优良性状互补,早熟、矮秆、抗病、优质、皮薄,糯香和适口性较好,穗型美观,籽粒较大,商品性好。
DOI:10.3969/j.issn.1004-874X.2006.12.006URL [本文引用: 1]
田白糯1号是广州市田园农业科 技研究中心以TMN03-7-5-3-2-1-6作母本、以TMN16-2-4-6-1-7-3作父本选育而成的糯玉米新组合。该组合利用“温带血缘×亚 热带血缘”杂优模式,实现了优良性状互补,早熟、矮秆、抗病、优质、皮薄,糯香和适口性较好,穗型美观,籽粒较大,商品性好。
DOI:10.3969/j.issn.1672-755X.2006.01.018URL [本文引用: 1]
苏玉糯2号是江苏沿江地区农科所以自选系361为母本,自选系366为父本,于1994年组配成的糯玉米单交种,各级试验示范结果表明:苏玉糯2号比目前东南区和黄准海区生产上大面积种植的苏玉(糯)1号鲜果穗、干子粒产量各增产5%~10%以上,采收期早4~5 d,具有早熟,高产,优质的特点。
DOI:10.3969/j.issn.1672-755X.2006.01.018URL [本文引用: 1]
苏玉糯2号是江苏沿江地区农科所以自选系361为母本,自选系366为父本,于1994年组配成的糯玉米单交种,各级试验示范结果表明:苏玉糯2号比目前东南区和黄准海区生产上大面积种植的苏玉(糯)1号鲜果穗、干子粒产量各增产5%~10%以上,采收期早4~5 d,具有早熟,高产,优质的特点。
DOI:10.3969/j.issn.1007-5739.2012.12.026URL [本文引用: 1]
苏玉糯1号是国内首个国家级审定的糯玉米品种,具有优质、高产、 多抗等优良特性。作为发展特色农业和开发名特优产品,该品种在农村产业结构调整中发挥了巨大的作用,取得良好的经济效益和社会效益。在其带动下,一大批育 种部门及个人纷纷开展糯玉米育种工作.促进了我国糯玉米事业的发展。介绍苏玉糯1号的特性及其作为种质在生产上的应用情况,以促进苏玉糯1号的应用。
DOI:10.3969/j.issn.1007-5739.2012.12.026URL [本文引用: 1]
苏玉糯1号是国内首个国家级审定的糯玉米品种,具有优质、高产、 多抗等优良特性。作为发展特色农业和开发名特优产品,该品种在农村产业结构调整中发挥了巨大的作用,取得良好的经济效益和社会效益。在其带动下,一大批育 种部门及个人纷纷开展糯玉米育种工作.促进了我国糯玉米事业的发展。介绍苏玉糯1号的特性及其作为种质在生产上的应用情况,以促进苏玉糯1号的应用。
URL [本文引用: 1]
“江 南花糯”系江苏省农业科学院粮食作物研究所最新育成的糯质鲜食玉米杂交新组合。该组合肉质厚、糯性强、香味浓、色泽鲜艳、甜度高 ,且不易与普通玉米混淆 ,现已在江苏大面积推广 ,并在浙江、上海、安徽、河南、重庆、四川、湖北、湖南、广东等地示范应用。1 亲本来源及
URL [本文引用: 1]
“江 南花糯”系江苏省农业科学院粮食作物研究所最新育成的糯质鲜食玉米杂交新组合。该组合肉质厚、糯性强、香味浓、色泽鲜艳、甜度高 ,且不易与普通玉米混淆 ,现已在江苏大面积推广 ,并在浙江、上海、安徽、河南、重庆、四川、湖北、湖南、广东等地示范应用。1 亲本来源及
DOI:10.3969/j.issn.1000-6400.2007.04.011URL [本文引用: 1]
郑白糯6号是河南省农业科学院粮食作物研究所选育的糯玉米单交种。其母本郑白糯06是以 “98W-2”为基础材料.连续自交选育而成。该系中早熟.矮秆.抗病、抗倒。籽粒大。叶片略宽。品质好。配合力高。父本郑白糯01是以“意大利黑玉米× 掖478”为基础材料.连续自交三代后选白粒再与掖478回交一次.而后又选白粒连续自交选育而成。该系中熟。矮秆、抗倒。高抗茎腐病。株型好。结实性 好。配合力高。
DOI:10.3969/j.issn.1000-6400.2007.04.011URL [本文引用: 1]
郑白糯6号是河南省农业科学院粮食作物研究所选育的糯玉米单交种。其母本郑白糯06是以 “98W-2”为基础材料.连续自交选育而成。该系中早熟.矮秆.抗病、抗倒。籽粒大。叶片略宽。品质好。配合力高。父本郑白糯01是以“意大利黑玉米× 掖478”为基础材料.连续自交三代后选白粒再与掖478回交一次.而后又选白粒连续自交选育而成。该系中熟。矮秆、抗倒。高抗茎腐病。株型好。结实性 好。配合力高。
URL [本文引用: 1]
糯玉米起源于中国,是玉米属玉米种的一个亚种.糯玉米经数百年的 变异,在株高、粒色、产量、抗性、品质等方面都形成了较大差异,是中国宝贵的玉米种质资源.糯玉米具有甜、粘、香的特点,风味优于普通玉米,其可溶性糖高 于普通玉米而低于甜玉米,更适合当前发达国家人们对清淡口味的追求.可以预料,随着人们生活水平的提高,以及对糯玉米营养特点的认识,用糯玉米作为原料加 工的产品,一定会以独特的风味加入到人们日益丰富的食谱中去.因此,深入研究糯玉米的遗传多样性,对糯玉米的新品种选育具有重要的现实和理论价值.该研究 利用玉米核DNA SSR和叶绿体DNA SSR...
URL [本文引用: 1]
糯玉米起源于中国,是玉米属玉米种的一个亚种.糯玉米经数百年的 变异,在株高、粒色、产量、抗性、品质等方面都形成了较大差异,是中国宝贵的玉米种质资源.糯玉米具有甜、粘、香的特点,风味优于普通玉米,其可溶性糖高 于普通玉米而低于甜玉米,更适合当前发达国家人们对清淡口味的追求.可以预料,随着人们生活水平的提高,以及对糯玉米营养特点的认识,用糯玉米作为原料加 工的产品,一定会以独特的风味加入到人们日益丰富的食谱中去.因此,深入研究糯玉米的遗传多样性,对糯玉米的新品种选育具有重要的现实和理论价值.该研究 利用玉米核DNA SSR和叶绿体DNA SSR...
DOI:10.3321/j.issn:0366-6964.2004.01.007URL [本文引用: 1]
选用5个多态性较好的微卫星标记,检测了山东省仅存的5个地方鸡种:寿光鸡、日照麻鸡、莱芜黑鸡、济宁百日鸡、鲁西斗鸡,以及一个外来鸡种--安卡黄鸡和一个外省地方鸡种--广西黄鸡共7个鸡种的遗传多样性.根据测试结果计算了每个等位基因的频率,并以基因频率为基础计算了Nei氏标准遗传距离(Ds)和DA遗传距离,发现日照麻鸡与济宁百日鸡的距离最近,而鲁西斗鸡与其他6个鸡种距离都较远.根据两种遗传距离分别进行了NJ法和UPGMA法聚类,得到4个聚类图.结果表明:DA遗传距离的UPGMA聚类图比较可靠.
DOI:10.3321/j.issn:0366-6964.2004.01.007URL [本文引用: 1]
选用5个多态性较好的微卫星标记,检测了山东省仅存的5个地方鸡种:寿光鸡、日照麻鸡、莱芜黑鸡、济宁百日鸡、鲁西斗鸡,以及一个外来鸡种--安卡黄鸡和一个外省地方鸡种--广西黄鸡共7个鸡种的遗传多样性.根据测试结果计算了每个等位基因的频率,并以基因频率为基础计算了Nei氏标准遗传距离(Ds)和DA遗传距离,发现日照麻鸡与济宁百日鸡的距离最近,而鲁西斗鸡与其他6个鸡种距离都较远.根据两种遗传距离分别进行了NJ法和UPGMA法聚类,得到4个聚类图.结果表明:DA遗传距离的UPGMA聚类图比较可靠.
DOI:10.3864/j.issn.0578-1752.2016.14.011URL [本文引用: 1]
【目的】枣原产中国,种质资源丰富。对来自中国22个省区不同用途的255个枣品种进行SSR分析,揭示这些不同产地来源的枣种质资源之间的亲缘关系和群体遗传结构,为枣种质资源的科学管理和分子标记辅助育种提供参考。【方法】利用改良CTAB提取供试枣种质的基因组DNA,以前期枣基因组测序挖掘出的SSR引物为基础,进行高效率引物筛选,并利用SSR分子标记技术对255份枣种质资源的基因组DNA进行PCR扩增,然后利用8%聚丙烯酰胺凝胶电泳分离,银染后显色。根据条带有无统计数据,计算出多态性位点百分率(PIC),用NTSYS软件进行UPGMA聚类分析;利用Structure软件分析群体遗传结构,计算出最适群体数目,构建遗传结构图。【结果】从64对SSR引物中筛选出23对高效率SSR引物,在供试材料中共检测出117个多态性位点,各引物扩增的多态性位点数为2—10条,每对引物平均扩增多态位点数为5.09个,PIC值变幅为0.359—0.727,平均为0.548,这些多态性引物可应用到其他枣种质资源的研究中;建立了只需1—2个标记就可鉴别出来的部分枣品种的SSR指纹,可用于这些品种的快速分子鉴定;255个枣品种的聚类分析将所有枣品种分为15个亚类,包括4个大类和11个小类,不同品种间的相似系数范围0.71—1.00,其中北京花生枣单独聚为一类,与其他枣品种关系较远;‘奉节鸡蛋枣’和‘溆浦鸡蛋枣’、‘陕西奶枣’和‘天津大马牙枣’的相似系数均为1.00;结合聚类图、供试品种的用途和原产地分析,不同枣品种间的亲缘关系与品种原产地有一定相关性,但和品种用途没有显著相关性。群体结构分析中,通过绘制K与ΔK的关系图,K=15时,ΔK最大,据此将255个枣品种也同样划分为15个群体,与聚类分析结果基本一致;进一步分析表明,各群体中大部分品种血缘关系比较单一,较少品种含有其他类群的遗传成分。总体看,山西或陕西的枣品种出现在绝大部分居群中,说明这两个省的资源在不同群体间的基因交流中发挥了重要作用;南方栽培区域中来自湖南的枣品种形成了相对独立的居群,可能是其起源相对单一,且在长期栽培过程中和其他产地枣品种间基因交流较少所致。上述结果表明,供试枣品种中与来源区域相关性明显的品种由相同地域内枣品种演化而来,而另一部分与来源产地相关性不明显的品种则是由不同区域间品种经过频繁的基因交流和重组选育而来,融合了不同区域品种的特点,从而没有了明显的区域特征。【结论】不同地理环境在枣品种的群体进化中发挥了较重要的作用,影响了不同产地间枣种质资源的遗传结构组成。
DOI:10.3864/j.issn.0578-1752.2016.14.011URL [本文引用: 1]
【目的】枣原产中国,种质资源丰富。对来自中国22个省区不同用途的255个枣品种进行SSR分析,揭示这些不同产地来源的枣种质资源之间的亲缘关系和群体遗传结构,为枣种质资源的科学管理和分子标记辅助育种提供参考。【方法】利用改良CTAB提取供试枣种质的基因组DNA,以前期枣基因组测序挖掘出的SSR引物为基础,进行高效率引物筛选,并利用SSR分子标记技术对255份枣种质资源的基因组DNA进行PCR扩增,然后利用8%聚丙烯酰胺凝胶电泳分离,银染后显色。根据条带有无统计数据,计算出多态性位点百分率(PIC),用NTSYS软件进行UPGMA聚类分析;利用Structure软件分析群体遗传结构,计算出最适群体数目,构建遗传结构图。【结果】从64对SSR引物中筛选出23对高效率SSR引物,在供试材料中共检测出117个多态性位点,各引物扩增的多态性位点数为2—10条,每对引物平均扩增多态位点数为5.09个,PIC值变幅为0.359—0.727,平均为0.548,这些多态性引物可应用到其他枣种质资源的研究中;建立了只需1—2个标记就可鉴别出来的部分枣品种的SSR指纹,可用于这些品种的快速分子鉴定;255个枣品种的聚类分析将所有枣品种分为15个亚类,包括4个大类和11个小类,不同品种间的相似系数范围0.71—1.00,其中北京花生枣单独聚为一类,与其他枣品种关系较远;‘奉节鸡蛋枣’和‘溆浦鸡蛋枣’、‘陕西奶枣’和‘天津大马牙枣’的相似系数均为1.00;结合聚类图、供试品种的用途和原产地分析,不同枣品种间的亲缘关系与品种原产地有一定相关性,但和品种用途没有显著相关性。群体结构分析中,通过绘制K与ΔK的关系图,K=15时,ΔK最大,据此将255个枣品种也同样划分为15个群体,与聚类分析结果基本一致;进一步分析表明,各群体中大部分品种血缘关系比较单一,较少品种含有其他类群的遗传成分。总体看,山西或陕西的枣品种出现在绝大部分居群中,说明这两个省的资源在不同群体间的基因交流中发挥了重要作用;南方栽培区域中来自湖南的枣品种形成了相对独立的居群,可能是其起源相对单一,且在长期栽培过程中和其他产地枣品种间基因交流较少所致。上述结果表明,供试枣品种中与来源区域相关性明显的品种由相同地域内枣品种演化而来,而另一部分与来源产地相关性不明显的品种则是由不同区域间品种经过频繁的基因交流和重组选育而来,融合了不同区域品种的特点,从而没有了明显的区域特征。【结论】不同地理环境在枣品种的群体进化中发挥了较重要的作用,影响了不同产地间枣种质资源的遗传结构组成。
DOI:10.11869/j.issn.100-8551.2014.02.0177URL [本文引用: 1]
为了确定大麦亲本材料间的亲缘关系,为后期关联分析奠定基础,本 研究利用71个SSR标记对不同来源的99份大麦材料进行了遗传多样性及群体遗传结构分析.结果表明,71个SSR标记共检测到184个等位变异,变幅为 2-5个,平均每个标记有2.6个.Shannon指数变幅为0.0565 ~1.2241,平均值为0.6086.标记多态性信息含量(PIC)的变幅介于0.0200 ~0.6633之间,平均值为0.3729.聚类结果表明,试验品种的遗传相似系数(GS)的变异范围为0.5109 ~ 0.9511,平均值为0.7202.GS值在0.7100水平上分为5大类,分别包括5、4、3、6和79份材料;主坐标分析将材料分为5个亚群,分别 包括5、10、12、24和26份材料.群体遗传结构分析将材料分为5个亚群,分别包含24、18、6、22、29份材料.
DOI:10.11869/j.issn.100-8551.2014.02.0177URL [本文引用: 1]
为了确定大麦亲本材料间的亲缘关系,为后期关联分析奠定基础,本 研究利用71个SSR标记对不同来源的99份大麦材料进行了遗传多样性及群体遗传结构分析.结果表明,71个SSR标记共检测到184个等位变异,变幅为 2-5个,平均每个标记有2.6个.Shannon指数变幅为0.0565 ~1.2241,平均值为0.6086.标记多态性信息含量(PIC)的变幅介于0.0200 ~0.6633之间,平均值为0.3729.聚类结果表明,试验品种的遗传相似系数(GS)的变异范围为0.5109 ~ 0.9511,平均值为0.7202.GS值在0.7100水平上分为5大类,分别包括5、4、3、6和79份材料;主坐标分析将材料分为5个亚群,分别 包括5、10、12、24和26份材料.群体遗传结构分析将材料分为5个亚群,分别包含24、18、6、22、29份材料.
DOI:10.11869/hnxb.2013.12.1828URL [本文引用: 1]
This study was carried out to analyze the genetic diversity of maize inbred lines and to study genetic structure of germplasm population in order to provide the theoretical basis for preparing dominant hybrid seed. Simple sequence repeat (SSR) analysis of the 70 loci distributed uniformly throughout the maize genome was used to detect genetic variation among 96 maize inbred lines and 6 testers. The population genetic structure was assessed by using Structure2.3.1 software. Setting subsets(K) for 1-9,using 螖K(k) to determine the most suitable value of K, and then to generate the Q-matrix. Seventy loci identified 213 polymorphic bands. The average number of allele per SSR locus was 3.610 with a range from 2 to 7. The polymorphism information content(PIC) for the SSR loci varied from0.216 to 0.846 with an average of 0.614. Genetic similarities among the materials in this study ranged from 0.282 to 0.882 with an average of 0.670. The inbred lines could be classified into 6 clusters by UPGMA (unweighted pair group method arithmetic average) which could be simplified into SS (LRC,PA,Reid) and NSS(PB,SPT, Lancaster) heterotic groups. Assessment of population structure showed that the most appropriate K=7, namely, there were 7 sub-groups in the target population. The defined population analysis of inbred lines provided the basis for selecting new hybridization combinations.
DOI:10.11869/hnxb.2013.12.1828URL [本文引用: 1]
This study was carried out to analyze the genetic diversity of maize inbred lines and to study genetic structure of germplasm population in order to provide the theoretical basis for preparing dominant hybrid seed. Simple sequence repeat (SSR) analysis of the 70 loci distributed uniformly throughout the maize genome was used to detect genetic variation among 96 maize inbred lines and 6 testers. The population genetic structure was assessed by using Structure2.3.1 software. Setting subsets(K) for 1-9,using 螖K(k) to determine the most suitable value of K, and then to generate the Q-matrix. Seventy loci identified 213 polymorphic bands. The average number of allele per SSR locus was 3.610 with a range from 2 to 7. The polymorphism information content(PIC) for the SSR loci varied from0.216 to 0.846 with an average of 0.614. Genetic similarities among the materials in this study ranged from 0.282 to 0.882 with an average of 0.670. The inbred lines could be classified into 6 clusters by UPGMA (unweighted pair group method arithmetic average) which could be simplified into SS (LRC,PA,Reid) and NSS(PB,SPT, Lancaster) heterotic groups. Assessment of population structure showed that the most appropriate K=7, namely, there were 7 sub-groups in the target population. The defined population analysis of inbred lines provided the basis for selecting new hybridization combinations.
.
DOI:10.1007/s00122-009-1108-0URLPMID:19639296 [本文引用: 1]
The highland region or Northwestern Argentina (NWA) is one of the southernmost areas of native maize cultivation and constitutes an expansion of the peruvian Andes sphere of influence. To examine the genetic diversity and racial affiliations of the landraces cultivated in this area, 18 microsatellite markers were used to characterize 147 individuals from 6 maize races representative of traditional materials. For the whole data set, a total of 184 alleles were found, with an average of 10.2 alleles per locus. The average gene diversity was 0.571. The observed patterns of genetic differentiation suggest that historical association is probably the main factor in shaping population structure for the landraces studied here. In agreement with morphological and cytogenetic data, Bayesian analysis of NWA landraces revealed the occurrence of three main gene pools. Assessment of racial affiliations using a combined dataset including previous data on American landraces showed a clear relationship between one of these gene pools and typical Andean races, whereas the remaining two gene pools exhibited a closer association to Caribbean accessions and native germplasm from the United States, respectively. These results highlight the importance of integrating regional genetic studies if a deeper understanding of maize diversification and dispersal is to be achieved.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}