 ,1,**, 胡丹3,**, 聂蛟1, 黄科慧1, 张玉珂1, 张园莉1, 佘恒志1, 方小梅1,2, 阮仁武1,2, 易泽林,1,2,*
,1,**, 胡丹3,**, 聂蛟1, 黄科慧1, 张玉珂1, 张园莉1, 佘恒志1, 方小梅1,2, 阮仁武1,2, 易泽林,1,2,*Genetic Analysis of Plant Height and Stem Diameter in Common Buckwheat
LI Ying-Shuang,1,**, HU Dan3,**, NIE Jiao1, HUANG Ke-Hui1, ZHANG Yu-Ke1, ZHANG Yuan-Li1, SHE Heng-Zhi1, FANG Xiao-Mei1,2, RUAN Ren-Wu1,2, YI Ze-Lin,1,2,*通讯作者:
第一联系人:
收稿日期:2017-11-9接受日期:2018-04-11网络出版日期:2018-06-11
| 基金资助: |
Received:2017-11-9Accepted:2018-04-11Online:2018-06-11
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (587KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
李英双, 胡丹, 聂蛟, 黄科慧, 张玉珂, 张园莉, 佘恒志, 方小梅, 阮仁武, 易泽林. 甜荞株高和茎粗的遗传分析[J]. 作物学报, 2018, 44(8): 1185-1195. doi:10.3724/SP.J.1006.2018.01185
LI Ying-Shuang, HU Dan, NIE Jiao, HUANG Ke-Hui, ZHANG Yu-Ke, ZHANG Yuan-Li, SHE Heng-Zhi, FANG Xiao-Mei, RUAN Ren-Wu, YI Ze-Lin.
甜荞(Fagopyrum esculentum M.)起源于中国, 具有生育期短、适应性强特点, 几个世纪以来一直是受欢迎的谷物和饲料作物[1]。甜荞营养元素全面, 含有较高的蛋白质、天然抗氧化剂、类黄酮、矿物质、维生素和膳食纤维[2,3], 具有抗过敏、利尿、镇咳、降血脂、降血压、防癌抗癌等功效, 是食药兼用型粮食作物[4,5,6,7], 已成为21世纪人类的绿色食品之一[8,9]。甜荞在生长发育过程中受到风雨等外界自然条件的影响后容易发生茎秆倒伏和弯曲, 是当前甜荞生产中存在的一个致命因素, 其轻者导致减产重者绝收, 极大地制约了甜荞产量及农民种植的积极性[10,11]。因此, 研究荞麦茎秆抗倒伏规律对实现荞麦高产、稳产具有重要意义。
株高和茎粗是评定作物抗倒伏能力的重要指标。在一定株高范围内, 植株越高, 倒伏率越高, 同时, 茎秆的节间长度越短, 基部茎秆越粗, 植株越不易倒伏[12,13,14,15,16]。目前主要通过调控种植密度[17]、烯效唑拌种[18]、茎秆解剖结构以及木质素合成研究甜荞倒伏[19,20], 对甜荞株高、茎粗的遗传分析尚未见相关报道。盖钧镒等[21]提出适合植物遗传分析的1对主基因+多基因和2对主基因+多基因混合模型的单世代和联合多世代的分析方法, 已经广泛应用于不同作物不同性状的遗传分析, 如水稻种子抗老化[22]、棉花高品质纤维性状[23]、玉米种子休眠[24]、小麦雄性不育[25]、大豆的根系性状[26]、花生产量[27]、油菜株高和抗倒性[28,29]、油菜花色[30]等。本文初步分析甜荞株高和茎粗的遗传效应, 为深入解析株高和茎粗的遗传机制奠定基础, 也为甜荞抗倒伏遗传改良与生产提供依据。
1 材料与方法
1.1 试验设计
亲本材料分别为高秆健壮品种酉荞2号(P1)和矮秆纤细品种乌克兰大粒荞(P2), 前者为本课题组育成的品种, 后者由重庆市荞麦产业体系提供。试验材料种植在西南大学歇马科研基地(10°3′53″~29°39′10″N, 106°18′14″~106°56′53″E), 试验田土壤为沙壤土。2014年秋季, 在隔离区内配制两亲本的正、反交组合, 初花期鉴定长短柱头并去除异类型花, 酉荞2号群体去除短花柱植株, 留长花柱植株, 乌克兰群体去除长花柱植株, 留短花柱植株, 成熟后分别收获两品种植株上的种子, 分别为正、反交F1代。2015年春季, F1隔离种植正交F1代不同植株间杂交, 反交F1代不同植株间杂交, 分别收获两群体的全部种子, 获得正、反交F2代。正、反交F1代与双亲各自回交, 在母本上收获的种子分别为正、反交B1和B2代。
2015年秋季, 将正、反交组合的6个世代分别播种, 行长3.00 m, 行距0.33 m, 株距0.20 m。P1、P2、F1种植4行, B1、B2和F2种植10行, 周围种植3行保护行进行隔离。
成熟期在田间随机选取一定数量单株(表1), 用直尺测量茎秆基部至植株顶端的距离作为株高, 用游标卡尺测量茎秆最粗的部位作为茎粗。
Table 1
表1
表1正、反交组合6个世代的株高与茎粗
Table 1
| 亲本组合 Parent combination | 世代 Generation | 株数 No. of plants | 株高 Plant height | 茎粗 Stem diameter | ||
|---|---|---|---|---|---|---|
| 平均值±标准差 Mean±SD (cm) | 变异系数 CV (%) | 平均值±标准差 Mean±SD (mm) | 变异系数 CV (%) | |||
| 亲本 Parent | P1 | 25 | 106.08±8.05 | 7.59 | 6.10±0.94 | 6.49 |
| P2 | 25 | 75.32±6.01 | 7.98 | 4.80±0.59 | 8.14 | |
| P1 × P2 | F1 | 28 | 93.04±8.64 | 9.29 | 5.30±0.70 | 7.57 |
| B1 | 153 | 100.12±10.21 | 10.20 | 5.99±0.91 | 6.58 | |
| B2 | 165 | 84.87±8.90 | 10.49 | 6.02±0.82 | 7.34 | |
| F2 | 274 | 94.67±10.23 | 10.81 | 6.50±1.00 | 6.50 | |
| P2 × P1 | F1 | 28 | 87.54±6.62 | 7.56 | 5.10±0.64 | 7.97 |
| B1 | 136 | 87.04±5.96 | 6.85 | 5.10±0.70 | 7.29 | |
| B2 | 133 | 91.51±6.36 | 6.95 | 5.40±0.70 | 7.71 | |
| F2 | 303 | 90.17±7.61 | 8.44 | 5.73±0.75 | 7.64 | |
新窗口打开|下载CSV
1.2 遗传模型分析
根据盖钧镒等[21]提出的植物数量性状主基因+多基因混合遗传多世代联合分析方法, 对甜荞株高和茎粗进行联合分析, 采用极大似然法和IECM算法估计各世代、各成分分布的参数, 通过AIC值选择最佳模型[31], 并进行一组适合性检验, 包括均匀性U12、U22和U32检验、Kolmo-Smimov检验(nW2)和Kolmo-Gorov检验(Dn), 根据检验结果选择最优遗传模型。最后采用最小二乘法依据最优模型的各成分分布参数估计各基因效应值、方差等遗传参数。2 结果与分析
2.1 不同世代群体的株高和茎粗变异
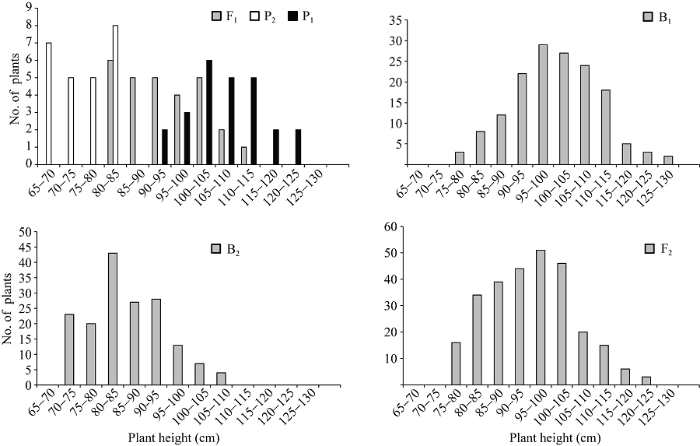
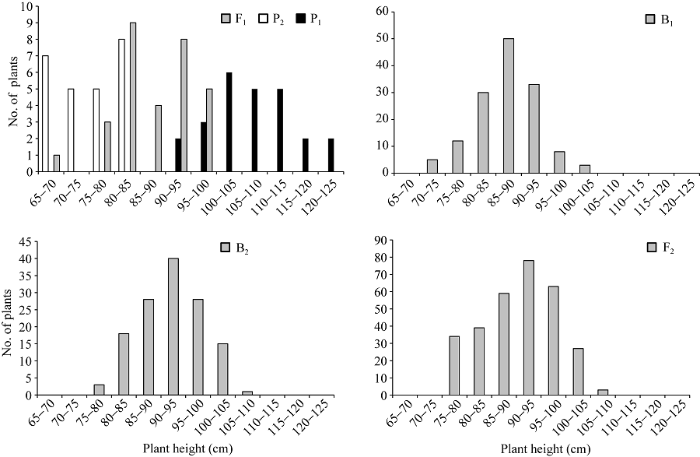
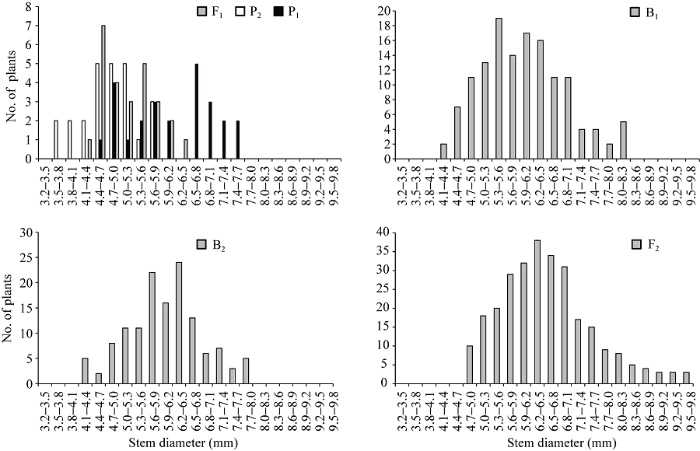
两亲本株高和茎粗平均值分别为90.70 cm和5.45 mm, 两亲本株高相差30.76 cm, 茎粗相差1.30 mm。在正交组合中, 株高和茎粗变异丰富(表1)。例如, F1株高大于双亲平均值, B1和B2代株高分别比低亲小24.80 cm和9.55 cm, F2代略偏向于高亲, B1、B2和F2群体的变异系数明显高于P1、P2和F1, 说明分离世代群体离散程度高, 遗传变异大。而在反交组合中, 株高和茎粗的变异较低(表1)。例如, F1株高小于双亲平均值, B1、B2代株高分别比低亲大11.72 cm和16.19 cm, F2略偏向于低亲, B1和B2代变异系数小于P1、P2和F1群体, 说明B1、B2群体的变异性较低, 但F2群体具有较高的遗传变异。B1、B2和F2群体茎粗的变异系数低于P1和F1群体, 说明分离世代群体的遗传多态性较低。正、反交组合6个世代的株高(附图1和附图2)和茎粗(附图3和附图4)均呈连续性分布, 具有典型的数量遗传特征。株高与茎粗在B1、B2和F2世代群体具有明显多峰现象, 说明株高和茎粗主要受主基因遗传效应影响。株高反交B1世代具有明显正态分布现象, 说明B1世代也受到部分多基因遗传效应影响。
附图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附图1正交组合6世代株高的次数分布
Supplementary fig. 1Frequency distributions of plant height in six generations derived from orthogonal cross
附图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附图2反交组合6世代株高的次数分布
Supplementary fig. 2Frequency distributions of plant height in six generations derived from back cross
附图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附图3正交组合6世代茎粗的次数分布
Supplementary fig. 3Frequency distributions of stem diameter in six generations derived from orthogonal cross
附图4
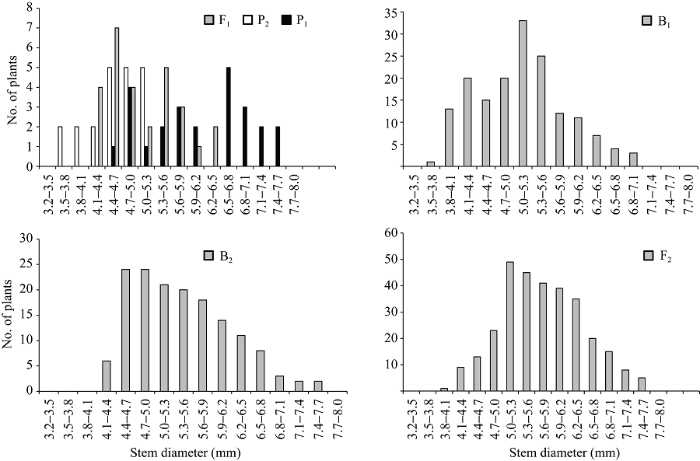 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附图4反交组合6世代茎粗的次数分布
Supplementary fig. 4Frequency distributions of stem diameter in six generations derived from back cross
2.2 株高和茎粗主基因+多基因遗传分析
根据AIC准则, 选取AIC值最小及与最小AIC值比较接近的3组遗传模型作为备选模型(附表1)。对正交组合, B-4、D-2、E-1为株高遗传的备选模型, B-1、E-0、E-1为茎粗遗传的备选模型; 同理, 株高与茎粗反交的3个备选模型均为C-0、D-0和E-1。Supplementary table 1
附表1
附表1正反交组合株高与茎粗表型分布的极大对数似然函数值和AIC值
Supplementary table 1
| 模型 Model | 模型含义 Implication of model | P1 × P2 | P2 × P1 | ||
|---|---|---|---|---|---|
| 株高 Plant height | 茎粗 Stem diameter | 株高 Plant height | 茎粗 Stem diameter | ||
| 极大对数似然函数值 log max likelihood value | |||||
| A-1 | 1MG-AD | -2471.03 | -911.79 | -2221.06 | -831.34 |
| A-2 | 1MG-A | -2471.27 | -913.18 | -2220.93 | -835.71 |
| A-3 | 1MG-EAD | -2517.11 | -912.04 | -2252.92 | -831.55 |
| A-4 | 1MG-AEND | -2542.66 | -907.83 | -2287.76 | -841.14 |
| B-1 | 2MG-ADI | -2462.85 | -854.59 | -2190.52 | -814.82 |
| B-2 | 2MG-AD | -2466.77 | -910.59 | -2221.34 | -832.65 |
| B-3 | 2MG-A | -2524.10 | -934.20 | -2281.29 | -874.56 |
| B-4 | 2MG-EA | -2466.05 | -913.57 | -2213.12 | -837.46 |
| B-5 | 2MG-AED | -2499.91 | -911.93 | -2230.42 | -833.39 |
| B-6 | 2MG-EEAD | -2499.91 | -911.93 | -2230.42 | -833.39 |
| C-0 | PG-ADI | -2473.89 | -859.01 | -2175.84 | -809.55 |
| C-1 | PG-AD | -2467.03 | -905.21 | -2214.12 | -838.68 |
| D-0 | MX1-AD-ADI | -2461.91 | -853.03 | -2174.53 | -803.64 |
| D-1 | MX1-AD-AD | -2466.94 | -906.60 | -2215.13 | -838.79 |
| D-2 | MX1-A-AD | -2461.37 | -893.08 | -2214.78 | -825.99 |
| D-3 | MX1-EAD-AD | -2466.32 | -908.12 | -2215.29 | -838.44 |
| D-4 | MX1-AEND-AD | -2466.40 | -905.01 | -2215.31 | -837.98 |
| E-0 | MX2-ADI-ADI | -2454.25 | -843.30 | -2174.53 | -803.63 |
| E-1 | MX2-ADI-AD | -2450.50 | -844.42 | -2161.64 | -791.16 |
| E-2 | MX2-AD-AD | -2466.33 | -908.12 | -2215.29 | -838.44 |
| E-3 | MX2-A-AD | -2467.93 | -906.89 | -2214.68 | -930.84 |
| E-4 | MX2-EA-AD | -2466.25 | -908.12 | -2215.29 | -838.44 |
| E-5 | MX2-AED-AD | -2466.32 | -908.12 | -2215.29 | -838.44 |
| E-6 | MX2-EEAD-AD | -2466.32 | -908.12 | -2215.29 | -838.44 |
| AIC值 AIC value | |||||
| A-1 | 1MG-AD | 4950.06 | 1831.59 | 4450.11 | 1670.67 |
| A-2 | 1MG-A | 4948.55 | 1832.36 | 4447.86 | 1677.43 |
| A-3 | 1MG-EAD | 5040.22 | 1830.08 | 4511.84 | 1669.09 |
| A-4 | 1MG-AEND | 5091.33 | 1821.66 | 4581.52 | 1688.29 |
| B-1 | 2MG-ADI | 4945.70 | 1729.19 | 4401.03 | 1649.64 |
| B-2 | 2MG-AD | 4945.53 | 1833.19 | 4454.67 | 1677.30 |
| B-3 | 2MG-A | 5056.20 | 1876.39 | 4570.58 | 1757.12 |
| B-4 | 2MG-EA | 4938.09 | 1833.14 | 4432.25 | 1680.93 |
| B-5 | 2MG-AED | 5007.81 | 1831.85 | 4468.85 | 1674.77 |
| 模型 Model | 模型含义 Implication of model | P1 × P2 | P2 × P1 | ||
| 株高 Plant height | 茎粗 Stem diameter | 株高 Plant height | 茎粗 Stem diameter | ||
| B-6 | 2MG-EEAD | 5005.81 | 1829.85 | 4466.85 | 1672.78 |
| C-0 | PG-ADI | 4967.78 | 1738.03 | 4371.69 | 1639.09 |
| C-1 | PG-AD | 4948.06 | 1824.42 | 4442.24 | 1691.36 |
| D-0 | MX1-AD-ADI | 4947.82 | 1730.07 | 4373.05 | 1631.28 |
| D-1 | MX1-AD-AD | 4951.88 | 1831.20 | 4448.26 | 1695.57 |
| D-2 | MX1-A-AD | 4938.73 | 1802.15 | 4445.57 | 1667.97 |
| D-3 | MX1-EAD-AD | 4948.64 | 1832.24 | 4446.58 | 1692.87 |
| D-4 | MX1-AEND-AD | 4948.81 | 1826.03 | 4446.63 | 1691.95 |
| E-0 | MX2-ADI-ADI | 4944.51 | 1722.60 | 4385.05 | 1643.27 |
| E-1 | MX2-ADI-AD | 4931.01 | 1718.83 | 4353.28 | 1612.32 |
| E-2 | MX2-AD-AD | 4954.65 | 1838.24 | 4452.58 | 1698.87 |
| E-3 | MX2-A-AD | 4953.87 | 1831.77 | 4447.37 | 1879.68 |
| E-4 | MX2-EA-AD | 4948.50 | 1832.24 | 4446.58 | 1692.87 |
| E-5 | MX2-AED-AD | 4950.64 | 1834.24 | 4448.58 | 1694.87 |
| E-6 | MX2-EEAD-AD | 4948.65 | 1832.24 | 4446.58 | 1692.87 |
新窗口打开|下载CSV
对备选模型进行一组适合性测验(均匀性检验、Smirnov检验和Kolmogorov检验的5个统计量U12、U22、U32、nW2和Dn), 选择统计量达到显著水平个数最少的模型为最适模型(表2), 正反交株高与茎粗的最佳遗传模型均为E-1模型, 即2对加性-显性-上位性主基因+加性-显性多基因模型。
Table 2
表2
表2正反交组合后代株高与茎粗备选遗传模型的适合性检验
Table 2
| 模型含义 Implication of model | 株高 Plant height | 茎粗 Stem diameter | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 模型 Model | U12 | U22 | U32 | nW2 | Dn | 模型 Model | U12 | U22 | U32 | nW2 | Dn | |
| P1 × P2 | ||||||||||||
| 2MG-EA | B-4 | 0 | 0 | 0 | 0 | 0 | B-1 | 0 | 0 | 2 | 0 | 0 |
| MX1-A-AD | D-2 | 0 | 0 | 0 | 0 | 0 | E-0 | 0 | 0 | 0 | 0 | 0 |
| MX2-ADI-AD | E-1 | 0 | 0 | 0 | 0 | 0 | E-1 | 0 | 0 | 0 | 0 | 0 |
| P2 × P1 | ||||||||||||
| 2MG-EA | C-0 | 0 | 2 | 0 | 1 | 0 | C-0 | 1 | 2 | 0 | 2 | 0 |
| MX1-A-AD | D-0 | 0 | 0 | 0 | 0 | 0 | D-0 | 0 | 0 | 0 | 0 | 0 |
| MX2-ADI-AD | E-1 | 0 | 0 | 0 | 0 | 0 | E-1 | 0 | 0 | 0 | 0 | 0 |
新窗口打开|下载CSV
2.3 正、反交组合株高和茎粗最适遗传模型遗传参数估算
根据选择的最佳模型E-1得出的极大似然估计值经计算得到株高的一阶(表3)、二阶遗传参数(表4), 正、反交组合中控制株高的两对主基因加性效应相等, 显性效应大于加性效应, 均为负向效应。正交组合中2对主基因显性效应均为负, 且|ha|<|hb|, 说明株高主基因存在杂种优势(负向), 且以第2对主基因的显性效应为主, 反交组合中|ha|>|hb|, 则以第1对主基因的显性效应为主。正交组合2对基因加性和显性效应之间的互作(jab和jba)分别为+8.44和-9.48, 表明基因互作对该正交组合株高影响较大。Table 3
表3
表3正、反交组合株高与茎粗一阶遗传参数估计值
Table 3
| 遗传参数 Genetic parameter | P1 × P2 | P2 × P1 | |||
|---|---|---|---|---|---|
| 株高 Plant height | 茎粗 Stem diameter | 株高 Plant height | 茎粗 Stem diameter | ||
| m | 102.81 | 7.01 | 97.10 | 6.53 | |
| da | -1.39 | 0.03 | -1.63 | -0.15 | |
| db | -1.39 | 0.03 | -1.63 | -0.15 | |
| ha | -6.59 | -0.50 | -7.03 | -0.30 | |
| hb | -7.91 | -0.08 | -4.19 | -0.16 | |
| i | -12.06 | -1.55 | -6.30 | -1.13 | |
| jab | 8.44 | 0.04 | 6.82 | -0.27 | |
| jba | -9.48 | -0.41 | 10.88 | 0.86 | |
| l | 10.86 | -0.63 | 8.06 | 0.07 | |
| [d] | 18.13 | 0.31 | -11.46 | -0.32 | |
| [h] | -6.03 | -0.23 | -6.24 | -1.31 | |
| ha/da | 4.73 | -15.53 | 4.31 | 2.04 | |
| hb/db | 5.68 | -2.35 | 2.57 | 1.08 | |
新窗口打开|下载CSV
Table 4
表4
表4正、反交组合株高与茎粗二阶遗传参数估计值
Table 4
| 组合 Combination | 遗传参数 Genetic parameter | B1 | B2 | F2 | |||
|---|---|---|---|---|---|---|---|
| 株高 Plant height | 茎粗 Stem diameter | 株高 Plant height | 茎粗 Stem diameter | 株高 Plant height | 茎粗 Stem diameter | ||
| P1 × P2 | σp2 | 104.83 | 0.82 | 79.15 | 0.76 | 113.91 | 1.16 |
| σmg2 | 47.94 | 0.31 | 50.25 | 0.33 | 85.76 | 0.74 | |
| σpg2 | 28.74 | 0.09 | 0.75 | 0 | 0 | 0 | |
| σe2 | 28.15 | 0.42 | 28.15 | 0.42 | 28.15 | 0.42 | |
| hmg2 (%) | 45.73 | 37.26 | 63.49 | 48.80 | 81.12 | 72.10 | |
| hpg2 (%) | 27.41 | 11.18 | 0.95 | 0 | 0 | 0 | |
| P2 × P1 | σp2 | 35.55 | 0.63 | 43.58 | 0.56 | 64.19 | 0.68 |
| σmg2 | 14.76 | 0.42 | 26.76 | 0.26 | 47.37 | 0.47 | |
| σpg2 | 3.98 | 0 | 0 | 0.08 | 0 | 0 | |
| σe2 | 16.82 | 0.21 | 16.82 | 0.21 | 16.82 | 0.21 | |
| hmg2 (%) | 41.51 | 76.22 | 66.18 | 47.12 | 81.81 | 82.51 | |
| hpg2 (%) | 11.19 | 0 | 0 | 14.53 | 0 | 0 | |
新窗口打开|下载CSV
正、反交组合中, F2代株高的主基因表现出较高的遗传力, 分别达81.12%和81.81%, 具有很高的选择率。正交组合中B1、B2和F2世代的主基因+多基因效应决定株高表型变异的73.14%、64.44%和81.12%, 反交组合中这3个世代的遗传效应分别为52.70%、66.18%和81.81%, 可见, 虽然环境因素对表型变异有一定贡献, 但环境效应明显小于遗传效应, 说明环境对甜荞株高的影响较小。
正、反交组合中控制茎粗的2对主基因加性相等, 显性效应大于加性效应, 2对主基因显性效应均为负, 且|ha|>|hb|, 说明茎粗主基因存在杂种优势(负向), 且以第1对主基因的显性效应为主。正交组合中两对基因加性和显性互作效应(jab和jba)分别为+0.04和-0.41, 而反交jab和jba分别为-0.27和+0.86, 表明两对基因互作对茎粗影响较大。
正、反交组合F2世代茎粗的主基因表现出较高的遗传力, 分别达72.10%和82.51%, 具有较高的选择率。正交组合中, 主基因+多基因分别解释了B1、B2和F2代茎粗变异的48.44%、48.80%和72.10%, 多基因遗传率(11.18%)仅在B1代表现; 各群体环境变异占表型变异的36.21%~55.26%, 平均47.56%。反交组合中, 主基因+多基因可分别解释B1、B2和F2代茎粗变异的76.22%、61.65%和82.51%, 多基因遗传率(14.53%)只在B2代存在; 各群体环境变异占表型变异的30.88%~37.50%, 平均33.90%。可见, 环境对甜荞茎粗影响较大。
3 讨论
株高和茎粗是评定作物抗倒伏能力的重要指标, 明确其遗传规律是指导育种实践的基础, 陈桂华等[32]研究表明, 株高、节间粗度、单茎鲜重和弯曲力矩等是影响水稻抗倒伏能力的重要因素; 朱新开等[33]发现, 矮秆、基部节间短与重心低有利于小麦抗倒伏; 丰光等[34]发现玉米倒伏与株高极显著正相关。在荞麦上, 刘星贝等[35]研究发现茎秆形态特性与抗倒伏能力密切相关, 植株越矮、茎秆越粗壮、节间越密集、节间充实度高、机械组织层数多、机械组织和茎壁厚、维管束数目多且面积大会增强茎秆抗倒伏性能。主基因+多基因混合模型联合多世代分析法自问世以来, 已在多种作物的株高和茎粗研究中得到应用, 如高淀粉玉米株高符合1对加性主基因+加性-显性多基因模型[36], 甘蓝型油菜株高受到1对加性-显性主基因+加性-显性-上位性多基因混合遗传模型控制(D-0模型)[29]。甜荞株高和茎粗相关遗传分析尚未见报道。本研究表明, 株高和茎粗的最佳遗传模型均为E-1模型, 即2对加性-显性-上位性主基因+加性-显性多基因模型, 其中显性效应大于加性效应, 表现为超显性, 这与玉米[36]和甘蓝型油菜[29]上的报道不同, 可能与不同物种的遗传机制差异有关。
从遗传率来看, 本研究正、反交组合株高和茎粗的遗传率范围分别为52.7%~81.81%和48.44%~ 82.51%, 其中主基因遗传率大于多基因遗传率。株高正交组合多基因遗传率在B1和B2世代存在, 反交组合只在B1世代存在; 茎粗正交组合多基因遗传率仅在B1世代表现, 反交组合只在B2世代存在, 正、反交组合株高和茎粗多基因遗传率在F2代都为0, 主基因遗传率在F2世代表现最高, 具有很高的选择率, 表明早期世代选择是有效的。油菜株高和茎粗主要受显性效应控制[37], 环境因素对株高的影响较小, 对茎粗有一定的影响。本研究得到相似结论, 甜荞株高各群体的环境变异占表型变异的25.46%~ 37.08%, 而遗传贡献率为62.92%~74.54%, 说明株高主要受显性效应控制; 茎粗各群体的环境变异占表型变异的33.54%~46.38%, 略低于遗传贡献率(54.62%~66.46%), 说明环境对甜荞茎粗有一定影响。无论正交还是反交, 株高和茎粗的最佳遗传模型均一致, 皆表现为超显性, 遗传率范围大致相同, 主基因遗传率大于多基因遗传率, 表明甜荞株高和茎粗不受细胞质遗传的影响。
植物数量性状的遗传模型分析方法与QTL检测主基因的数量相对一致[38], 由于分析所推论的基因只是概念上的基因, 难以做个别比较, 为了更好地解释本文中控制株高及茎粗的基因数目多少、效应值大小和是否为同一基因, 有必要利用分子标记进行QTL定位研究。因此可利用分子标记对甜荞F2群体开展株高和茎粗QTL分析, 为甜荞株型改良分子标记育种和图位克隆奠定前期研究基础。
4 结论
在高秆健壮品种酉荞2号和矮秆纤细品种乌克兰大粒荞正、反交组合衍生的P1、P2、F1、B1、B2和F2世代群体中, 株高和茎粗均呈连续分布, 且符合2对加性-显性-上位性主基因+加性-显性多基因模型。株高的主基因+多基因遗传率在80%以上, 可在早代进行株高选择, 以提高育种效率; 茎粗的主基因+多基因遗传率在80%以下, 环境因素对茎粗有一定效应, 可利用栽培措施提高甜荞抗倒伏能力。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
DOI:10.1007/s00217-006-0405-yURL [本文引用: 1]

To improve the use of common buckwheat ( Fagopyrum esculentum ), characterized by interesting nutritional properties, it could be used in pasta formulations. In particular, as buckwheat is devoid of the gluten-forming proteins, it might be an ingredient for celiac patient food. The aim of this study was to develop both fresh egg pastas integrated with buckwheat and fresh egg pasta analogues classifiable as gluten-free, based on buckwheat and rice flours. Matter loss in the cooking water and weight increase during cooking of buckwheat pasta were higher than those of a reference sample made of common wheat flour. As buckwheat integration increased, sample break strain was significantly lower, as a result of the progressive reduction in gluten content. In the production of gluten-free pasta analogues, wheat flour was substituted with rice flour, precooked rice flour or pregelatinized rice starch. Since samples containing precooked rice flour gave the best results, in terms of workability, break strain and weight increase during cooking, they were also produced on an industrial scale. Industrial gluten-free fresh egg pasta analogues were tougher and less deformable in comparison with the laboratory-produced samples. These results were determined by the presence of the double thermal pasteurisation treatment, which allows to obtain a better structure of the product, showing also a lower matter loss during cooking.
DOI:10.3892/mmr_00000228URLPMID:21472210 [本文引用: 1]
Buckwheat flour is well known for its highly antioxidative ingredient, rutin. We have undertaken to examine alterations in the characteristics of rutin treated with various proteins. In this study, the radical scavenging activities of a rutin-ovalbumin complex were examined. Dissolved rutin hydrate and ovalbumin were combined and boiled in water for 10 min. In the resulting rutin-ovalbumin complex, a new high molecular weight peak was detected using gel permeation chromatography analysis, and an existing high molecular weight area of ovalbumin was observed to be increased by the addition of rutin. This suggested that ovalbumin molecules produce a complex through their interaction with rutin. Alkaline luminol chemiluminescence and electron spin resonance analysis revealed the formation of a rutin-ovalbumin complex that markedly enhanced the peroxyl, but not the hydroxyl, radical scavenging activity of rutin. Rutin also demonstrated antioxidative activity against hydroxyl radicals in a DNA protection assay. We therefore conclude that, compared with ovalbumin or rutin alone, the rutin-ovalbumin complex has improved antioxidative activities in the form of enhanced peroxyl radical scavenging activity and DNA protection from apurinic/apyrimidinic site formation caused by hydroxyl radicals.
DOI:10.3181/00379727-55-14532URL [本文引用: 1]
ConclusionsAt least in certain cases, rutin appears to have the property of decreasing capillary fragility in subjects in whom this fragility is initially increased. In this effect it resembles hesperidin.
[本文引用: 1]
DOI:10.1620/tjem.225.123URL [本文引用: 1]
DOI:10.3969/j.issn.1000-2340.2002.03.025URL [本文引用: 1]
测定结果表明:西藏日喀则地区的荞麦中富含蛋白质、脂肪、淀粉、维生素、矿物质、氨基酸等多种营养成分,与其它谷物如小麦、大米等相比,荞麦的蛋白质、脂肪含量较高,VB1,VB2,Vpp(烟酸),Vp(芦丁)的含量较为丰富.荞麦中含有多种易被人体吸收的矿物质元素,其中Mg含量较高,Fe含量较丰富.荞麦中含有8种人体必需氨基酸,且配比较合理.
DOI:10.3969/j.issn.1000-2340.2002.03.025URL [本文引用: 1]
测定结果表明:西藏日喀则地区的荞麦中富含蛋白质、脂肪、淀粉、维生素、矿物质、氨基酸等多种营养成分,与其它谷物如小麦、大米等相比,荞麦的蛋白质、脂肪含量较高,VB1,VB2,Vpp(烟酸),Vp(芦丁)的含量较为丰富.荞麦中含有多种易被人体吸收的矿物质元素,其中Mg含量较高,Fe含量较丰富.荞麦中含有8种人体必需氨基酸,且配比较合理.
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s13197-011-0316-1URLPMID:24425891 [本文引用: 1]
Germinated buckwheat is buckwheat seeds soaked in water just until it begins to bud. Buckwheat sprouts are seedling plants of buckwheat grown up to 10–1502cm. The purpose of this study was to determine the optimal growth period for accumulating the most abundant functional phenol(s) in germinated buckwheat that had been soaked in darkness and buckwheat sprouts cultivated by hydroponic culture. The rutin contained in germinated buckwheat was analyzed by CE (capillary electrophoresis). Phenols, including isoorientin, orientin, isovitexin, vitexin, and rutin were separated from buckwheat sprouts by HPLC and identified by LC-MS. The highest rutin content in germinated buckwheat was found to be 15.802mg/10002g DW at 2002h after germination. Buckwheat sprouts contained five kinds of major phenols. The highest amounts of isoorientin, orientin, isovitexin, and vitexin were measured at day023, with the exception of rutin, and then a gradual decrease was observed as the sprouts grew. The quantities of isoorientin, orientin, isovitexin, and vitexin at day023 were 5.8, 11.7, 26.2, and 28.902mg/10002g FW, respectively. The rutin content rapidly increased to 109.002mg/10002g FW until day026. The highest total phenols in buckwheat sprouts were 162.902mg/10002g FW at day026. Germinated buckwheat soaked for 2002h and buckwheat sprouts cultivated for 602days were rich in dietary phenol(s), which makes these plants a valuable functional food for human consumption.
URL [本文引用: 1]
ABSTRACT From the view point of Tartary buckwheat breeding, varietal difference of shoot growth characters related to lodging was investigated. Lodging degree (LD) was determined by the angle between the main stem and the ground. When calculated for all strains, the correlation coefficient between the shoot growth character and LD was the highest (negative) and significant for main stem length and main stem diameter showed the second highest (negative) and significant correlation coefficient. Three types of main stem elongation pattern were classified: mid-stem elongation type (main stem internode length increased markedly at the mid-position in ternodes), uniform elongation type (main stem internode length was almost constant) and the intermediate type. The mid-stem elongation type , usuaully tall and late maturing, was distributed from lowland areas to 3000 m, mainly in relatively low latitude areas, while the uniform elongation type, usally short and early maturing were distributed, in high altitude and middle latitude areas . In the mid-stem elongation type, (branch dry weight/main stem dry weight) and branch dry weight were highly and significantly correlated with LD, showing that the relative size of the branch to the main stem caused variation in LD. The number of main stem nodes were negatively correlated with LD in the uniform elongation type.
DOI:10.3969/j.issn.2095-0896.2007.05.018URL [本文引用: 1]
针对我国北方荞麦生产中倒伏严重影响产量的现状,设计、研究了多个抗倒伏栽培、调控试验方案。结果表明,选用抗倒品种、采用多效唑拌种、实行沟播培土对荞麦抗倒有较显著效果,可在生产实践中予以推广应用。
DOI:10.3969/j.issn.2095-0896.2007.05.018URL [本文引用: 1]
针对我国北方荞麦生产中倒伏严重影响产量的现状,设计、研究了多个抗倒伏栽培、调控试验方案。结果表明,选用抗倒品种、采用多效唑拌种、实行沟播培土对荞麦抗倒有较显著效果,可在生产实践中予以推广应用。
DOI:10.1139/b89-232URL [本文引用: 1]
Abstract Five registered Canadian six-row barley (Hordeum vulgare L.) cultivars, including two recently registered semidwarfs ('Duke' and 'Samson'), that differ in lodging resistance and height were studied over 2 years for differences in morphological and anatomical characteristics of culms that could be related to lodging resistance. Plants were grown in nonirrigated field plots at a population density of 220 plants/m2 under conditions of high soil fertility. Significant cultivar differences were observed for culm length, number of internodes, length of four basal internodes, culm diameter, culm wall thickness, number of vascular bundles, and thickness of the sclerenchyma ring. No cultivar differences were found for thickness of the sclerenchyma cell walls. Of the characters studied, culm length, basal internode length, culm wall thickness, and sclerenchyma ring thickness were most closely associated with differences in lodging resistance among the cultivars. The results of this study indicate that it may be possible to select lodging resistant genotypes from early generations in breeding programs on the basis of these traits.
DOI:10.1146/annurev.pp.23.060172.002253URL [本文引用: 1]
Bogolyubov's chain of equations for classical one-time correlation functions in a gas with binary collisions is used to obtain an equation for the long-wavelength (ka << 1) part of the binary correlation function g(2)(p(1)r(1), p(2)r(2), t) (a is the radius of the interaction between the particles and k is the wave vector in the Fourier decomposition of g(2) as a function of r(1)-r(2)). This equation is inhomogeneous and the right-hand side (source) is proportional to delta(r(1)-r(2)) and is nonvanishing in a nonequilibrium state (in the absence of detailed balance in the gas). In contrast to the well-known Bogolyubov function g(2), which describes the correlation at distances vertical bar r(1)-r(2)vertical bar < a, the correlation function that is obtained describes the correlation at distances of the order of the mean free path and greater. It does not follow adiabatically the change in time (and in space) of the first distribution function.
DOI:10.3321/j.issn:1000-4025.2007.11.034URL [本文引用: 1]
水稻茎秆形态结构特征和化学成分与其抗倒伏性能相关。本文对水稻茎秆高度、茎秆基部第1、2节间长度、茎秆粗细、茎壁厚度、厚壁组织(机械组织)数量和强度、维管束数量、细胞壁纤维素和木质素含量、细胞中碳水化合物积累的数量、硅与钾的含量以及茎秆抗倒伏相关的QTL等与水稻的抗倒伏性的相互关系进行了综述,为水稻抗倒伏优良品种性状的选育提供参考。
DOI:10.3321/j.issn:1000-4025.2007.11.034URL [本文引用: 1]
水稻茎秆形态结构特征和化学成分与其抗倒伏性能相关。本文对水稻茎秆高度、茎秆基部第1、2节间长度、茎秆粗细、茎壁厚度、厚壁组织(机械组织)数量和强度、维管束数量、细胞壁纤维素和木质素含量、细胞中碳水化合物积累的数量、硅与钾的含量以及茎秆抗倒伏相关的QTL等与水稻的抗倒伏性的相互关系进行了综述,为水稻抗倒伏优良品种性状的选育提供参考。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1626/pps.18.218URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.3321/j.issn:0253-9772.2008.02.016URL [本文引用: 1]
测定了粳稻品种丙8979与C堡杂交组合的P1、P2及其重组自交系350个株系种子劣变后的相对发芽率、相对发芽势和相对发芽指数,运用主基因+多基因混合遗传模型,对水稻种子的3个抗老化性状进行了遗传分析。结果表明,相对发芽率、相对发芽势、相对发芽指数均受2对加性-上位性主基因+加性?上位性多基因控制,且2对主基因的加性效应大于上位性效应。3性状均是主基因遗传率大于多基因遗传率。
DOI:10.3321/j.issn:0253-9772.2008.02.016URL [本文引用: 1]
测定了粳稻品种丙8979与C堡杂交组合的P1、P2及其重组自交系350个株系种子劣变后的相对发芽率、相对发芽势和相对发芽指数,运用主基因+多基因混合遗传模型,对水稻种子的3个抗老化性状进行了遗传分析。结果表明,相对发芽率、相对发芽势、相对发芽指数均受2对加性-上位性主基因+加性?上位性多基因控制,且2对主基因的加性效应大于上位性效应。3性状均是主基因遗传率大于多基因遗传率。
URL [本文引用: 1]
利用主基因与多基因混合遗传模型联合分析方法,通过纤维强度不同 的5个亲本配制的8个组合,研究了棉花主要纤维品质性状的遗传.联合分析发现,在不同性状不同组配方式的14个组合中,有12个存在主基因,表明了纤维性 状主基因存在的普遍性,以F2∶3家系的预测效果最好;双亲纤维品质性状均存在较大差异的组合--7235×TM1F2代强度主基因的遗传率为 0.196,麦克隆值为0.320,长度为0.139,回交世代的主基因遗传率小.除纤维长度总的显性效应为较高的正值外,其余各纤维性状的主基因显性与 多基因显性的总和为负值或接近0,杂合状态下大多数纤维品质性状表型值会偏向中亲值或低亲值,单纯依靠表型选择效率低.因此,很有必要对棉花品质性状进行 分子标记辅助育种选择.
URL [本文引用: 1]
利用主基因与多基因混合遗传模型联合分析方法,通过纤维强度不同 的5个亲本配制的8个组合,研究了棉花主要纤维品质性状的遗传.联合分析发现,在不同性状不同组配方式的14个组合中,有12个存在主基因,表明了纤维性 状主基因存在的普遍性,以F2∶3家系的预测效果最好;双亲纤维品质性状均存在较大差异的组合--7235×TM1F2代强度主基因的遗传率为 0.196,麦克隆值为0.320,长度为0.139,回交世代的主基因遗传率小.除纤维长度总的显性效应为较高的正值外,其余各纤维性状的主基因显性与 多基因显性的总和为负值或接近0,杂合状态下大多数纤维品质性状表型值会偏向中亲值或低亲值,单纯依靠表型选择效率低.因此,很有必要对棉花品质性状进行 分子标记辅助育种选择.
DOI:10.3969/j.issn.1005-0906.2007.02.002URL [本文引用: 1]
运用植物数量性状主基因+多基因混合遗传模型的方法对普通玉米自交系R08与A318杂交组合的P1、P2、F1和F2∶34个世代群体的种子休眠性进行了分析。结果表明:R08×A318组合种子休眠性的遗传符合一对加性-显性主基因+加性-显性-上位性多基因模型(D-0模型)。在F2∶3家系世代,主基因方差为0.9455,多基因方差为0.1196。主基因遗传率在F2∶3家系群体中为72.49%,多基因遗传率为9.17%。
DOI:10.3969/j.issn.1005-0906.2007.02.002URL [本文引用: 1]
运用植物数量性状主基因+多基因混合遗传模型的方法对普通玉米自交系R08与A318杂交组合的P1、P2、F1和F2∶34个世代群体的种子休眠性进行了分析。结果表明:R08×A318组合种子休眠性的遗传符合一对加性-显性主基因+加性-显性-上位性多基因模型(D-0模型)。在F2∶3家系世代,主基因方差为0.9455,多基因方差为0.1196。主基因遗传率在F2∶3家系群体中为72.49%,多基因遗传率为9.17%。
DOI:10.3321/j.issn:0496-3490.2007.09.025URL [本文引用: 1]
应用植物数量性状"主基因+多基因混合遗传模型"方法,分析了光温敏核雄性不育系BS210,与两个恢复系(BY149和O201)配制的2个杂交组合的亲本P1、P2、F1、F2育性的遗传效应.结果表明,两个组合F2的育性(结实率)次数分布均呈混合的正态分布,最适遗传模型均为E-1,即育性由两对加性-显性-上位性主基因和加性-显性多基因共同控制.两对主基因的加性效应近似相等,在两个组合中分别为-10.626、-10.068和-14.659、-14.655,主基因遗传力分别为25%和40%.两个组合的多基因加性效应分别为-6.225和5.025,多基因遗传力分别为16.67%和13.33%.两个组合的主效基因表现类似,但多基因效应存在较大的差异.环境对育性的影响较大,二系杂交小麦组合的育性受遗传因素和环境因素的共同控制.
DOI:10.3321/j.issn:0496-3490.2007.09.025URL [本文引用: 1]
应用植物数量性状"主基因+多基因混合遗传模型"方法,分析了光温敏核雄性不育系BS210,与两个恢复系(BY149和O201)配制的2个杂交组合的亲本P1、P2、F1、F2育性的遗传效应.结果表明,两个组合F2的育性(结实率)次数分布均呈混合的正态分布,最适遗传模型均为E-1,即育性由两对加性-显性-上位性主基因和加性-显性多基因共同控制.两对主基因的加性效应近似相等,在两个组合中分别为-10.626、-10.068和-14.659、-14.655,主基因遗传力分别为25%和40%.两个组合的多基因加性效应分别为-6.225和5.025,多基因遗传力分别为16.67%和13.33%.两个组合的主效基因表现类似,但多基因效应存在较大的差异.环境对育性的影响较大,二系杂交小麦组合的育性受遗传因素和环境因素的共同控制.
DOI:10.3969/j.issn.1671-8631.2012.05.014URL [本文引用: 1]
从301份黄淮海和长江中下游地区代表性大豆地方品种和育成品种(系)中按根系类型选取59份,在苗期干旱胁迫和非胁迫条件下对地上部和地下部性状进行2年重复鉴定,发现材料间性状隶属函数值具有丰富遗传变异,以株高、叶龄、根干重和茎叶干重隶属函数的算术平均数为抗旱综合指标从中筛选出汉中八月黄、晋豆14,科丰1号,圆黑豆等强耐旱型(1级)和临河大粉青、宁海晚黄豆等干旱敏感型(5级)材料。比根干重、比总根长、比根体积与耐旱隶属函数平均值均呈极显著正相关,可作为耐旱性的根系性状指标。利用“科丰1号×南农1138 2”(1级×4级)衍生的RIL群体为材料,对耐旱相关根系性状采用主基因+多基因混合遗传模型分离分
DOI:10.3969/j.issn.1671-8631.2012.05.014URL [本文引用: 1]
从301份黄淮海和长江中下游地区代表性大豆地方品种和育成品种(系)中按根系类型选取59份,在苗期干旱胁迫和非胁迫条件下对地上部和地下部性状进行2年重复鉴定,发现材料间性状隶属函数值具有丰富遗传变异,以株高、叶龄、根干重和茎叶干重隶属函数的算术平均数为抗旱综合指标从中筛选出汉中八月黄、晋豆14,科丰1号,圆黑豆等强耐旱型(1级)和临河大粉青、宁海晚黄豆等干旱敏感型(5级)材料。比根干重、比总根长、比根体积与耐旱隶属函数平均值均呈极显著正相关,可作为耐旱性的根系性状指标。利用“科丰1号×南农1138 2”(1级×4级)衍生的RIL群体为材料,对耐旱相关根系性状采用主基因+多基因混合遗传模型分离分
[本文引用: 1]
DOI:10.3321/j.issn:0496-3490.2008.03.005URL [本文引用: 1]
应用植物数量性状主基因+多基因混合遗传模型多世代联合分析方法,对甘蓝型油菜3个组合浙平1号×高芥1号(Ⅰ)、宁1243×高芥1号(Ⅱ)和宁3067×高芥1号(Ⅲ)的P1、P2、F1、B1、B2和F2等6个世代初花期单株抗折力进行了分析,结果表明:(1)组合Ⅰ抗倒伏性状的遗传受两对加性-显性-上位性主基因+加性-显性-上位性多基因控制,组合Ⅱ和组合Ⅲ抗倒伏性状的遗传受两对加性-显性-上位性主基因+加性-显性多基因控制。(2)在组合Ⅰ和组合Ⅱ中,控制抗倒伏性状的两对主基因都以加性效应为主,在组合Ⅲ中,控制抗倒伏性状的两对主基因以显性效应为主。(3)3个组合中控制抗倒伏性状的两对主基因都表现为抗倒对易倒为部分显性(-1h/d0)或完全显性(-∞h/d-1)。(4)两对主基因间存在明显的加性×加性(i)、加性×显性(jab)、显性×加性(jba)和显性×显性(l)的交互作用,多基因间也存在加性([d])和显性([h])的交互作用。(5)3个组合中,各群体的主基因遗传率为21.92%~64.00%,而多基因遗传率只在部分群体中检测到,为0.00%~38.61%,表明3个组合的抗倒伏性状主要是以主基因遗传。(6)环境变异占表型变异的30.95%~65.47%,表明环境因素对油菜抗倒伏性状的影响比较大。
DOI:10.3321/j.issn:0496-3490.2008.03.005URL [本文引用: 1]
应用植物数量性状主基因+多基因混合遗传模型多世代联合分析方法,对甘蓝型油菜3个组合浙平1号×高芥1号(Ⅰ)、宁1243×高芥1号(Ⅱ)和宁3067×高芥1号(Ⅲ)的P1、P2、F1、B1、B2和F2等6个世代初花期单株抗折力进行了分析,结果表明:(1)组合Ⅰ抗倒伏性状的遗传受两对加性-显性-上位性主基因+加性-显性-上位性多基因控制,组合Ⅱ和组合Ⅲ抗倒伏性状的遗传受两对加性-显性-上位性主基因+加性-显性多基因控制。(2)在组合Ⅰ和组合Ⅱ中,控制抗倒伏性状的两对主基因都以加性效应为主,在组合Ⅲ中,控制抗倒伏性状的两对主基因以显性效应为主。(3)3个组合中控制抗倒伏性状的两对主基因都表现为抗倒对易倒为部分显性(-1h/d0)或完全显性(-∞h/d-1)。(4)两对主基因间存在明显的加性×加性(i)、加性×显性(jab)、显性×加性(jba)和显性×显性(l)的交互作用,多基因间也存在加性([d])和显性([h])的交互作用。(5)3个组合中,各群体的主基因遗传率为21.92%~64.00%,而多基因遗传率只在部分群体中检测到,为0.00%~38.61%,表明3个组合的抗倒伏性状主要是以主基因遗传。(6)环境变异占表型变异的30.95%~65.47%,表明环境因素对油菜抗倒伏性状的影响比较大。
DOI:10.3724/SP.J.1006.2013.00207URL [本文引用: 3]
株型改良是油菜高产、优质育种的主攻方向之一。矮杆及半矮杆株型有利于提高植株抗倒伏能力和经济系数、减少收获难度。10D130是一个半矮杆新品系, 用10D130和常规优良品种中双11杂交, 构建6世代遗传群体(P1、F1、P2、B1、B2和F2), 以主基因+多基因混合遗传模型对该组合株高及其关联性状进行遗传分析。结果表明, 10D130×中双11组合株高、分枝部位、主花序长度的遗传均受到1对加性-显性-上位性主基因+加性-显性-上位性多基因控制(D-0模型)。其中, 株高性状加性效应值为–8.58, 显性效应值为7.44, 主基因遗传率在B1、B2和F2中分别为23.52%、0.91%和17.81%;一次有效分枝起始部位的1对主基因加性效应值为–22.11, 显性效应值为3.13, 主基因遗传率在B1、B2和F2中分别为49.95%、40.85%和61.15%;主花序长的主基因加性效应值为–2.21, 显性效应值为1.6, 主基因遗传率在B1、B2和F2中分别为0.68%、47.94%和40.07%。一次有效分枝间距的最适宜遗传模型为E-1模型(2对加-显-上位性主基因+加-显-上位性多基因混合遗传模型), 其中第1对主基因加性效应值为–0.55、显性效应值为–1.66, 第2对主基因加性效应值为0.74、显性效应值为–1.29, 均表现超显性遗传, 主基因遗传率在B1、B2和F2三个分离世代群体中分别为10.99%、38.65%和44.10%。一次有效分枝部位高度、主花序长、有效分枝节间距和有效分枝数与株高均呈显著正相关。
DOI:10.3724/SP.J.1006.2013.00207URL [本文引用: 3]
株型改良是油菜高产、优质育种的主攻方向之一。矮杆及半矮杆株型有利于提高植株抗倒伏能力和经济系数、减少收获难度。10D130是一个半矮杆新品系, 用10D130和常规优良品种中双11杂交, 构建6世代遗传群体(P1、F1、P2、B1、B2和F2), 以主基因+多基因混合遗传模型对该组合株高及其关联性状进行遗传分析。结果表明, 10D130×中双11组合株高、分枝部位、主花序长度的遗传均受到1对加性-显性-上位性主基因+加性-显性-上位性多基因控制(D-0模型)。其中, 株高性状加性效应值为–8.58, 显性效应值为7.44, 主基因遗传率在B1、B2和F2中分别为23.52%、0.91%和17.81%;一次有效分枝起始部位的1对主基因加性效应值为–22.11, 显性效应值为3.13, 主基因遗传率在B1、B2和F2中分别为49.95%、40.85%和61.15%;主花序长的主基因加性效应值为–2.21, 显性效应值为1.6, 主基因遗传率在B1、B2和F2中分别为0.68%、47.94%和40.07%。一次有效分枝间距的最适宜遗传模型为E-1模型(2对加-显-上位性主基因+加-显-上位性多基因混合遗传模型), 其中第1对主基因加性效应值为–0.55、显性效应值为–1.66, 第2对主基因加性效应值为0.74、显性效应值为–1.29, 均表现超显性遗传, 主基因遗传率在B1、B2和F2三个分离世代群体中分别为10.99%、38.65%和44.10%。一次有效分枝部位高度、主花序长、有效分枝节间距和有效分枝数与株高均呈显著正相关。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.7606/j.issn.1009-1041.2006.01.024Magsci [本文引用: 1]
选用6个株高不同的小麦品种,在自然倒伏条件下,研究籽粒形成期倒伏植株的茎秆性状、倒伏对产量和品质的影响。结果表明,矮秆、基部节间较短、重心高度较低、穗下节间较长且株高构成指数大的植株有利于抗倒伏。小麦在籽粒形成期倒伏对产量、品质均有较大的影响,表现为产量下降,籽粒中蛋白质含量、湿面筋含量、沉降值有所上升,但蛋白质产量下降,支链淀粉、直链淀粉和总淀粉含量亦下降。
DOI:10.7606/j.issn.1009-1041.2006.01.024Magsci [本文引用: 1]
选用6个株高不同的小麦品种,在自然倒伏条件下,研究籽粒形成期倒伏植株的茎秆性状、倒伏对产量和品质的影响。结果表明,矮秆、基部节间较短、重心高度较低、穗下节间较长且株高构成指数大的植株有利于抗倒伏。小麦在籽粒形成期倒伏对产量、品质均有较大的影响,表现为产量下降,籽粒中蛋白质含量、湿面筋含量、沉降值有所上升,但蛋白质产量下降,支链淀粉、直链淀粉和总淀粉含量亦下降。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 2]
利用植物数量性状主基因+多基因混合遗传模型,以高淀粉玉米杂交组合"郑单958"的P1、F1、P2、B1∶2、B2∶2和F2∶36个家系世代为材料,多世代联合分析了高淀粉玉米主要性状的遗传效应。结果表明:穗长、穗位高由多基因控制;百粒重、单穗重、行粒数、株高由1对加性主基因+加性-显性多基因控制;秃尖长由1对加性-显性主基因+加性-显性多基因控制。
URL [本文引用: 2]
利用植物数量性状主基因+多基因混合遗传模型,以高淀粉玉米杂交组合"郑单958"的P1、F1、P2、B1∶2、B2∶2和F2∶36个家系世代为材料,多世代联合分析了高淀粉玉米主要性状的遗传效应。结果表明:穗长、穗位高由多基因控制;百粒重、单穗重、行粒数、株高由1对加性主基因+加性-显性多基因控制;秃尖长由1对加性-显性主基因+加性-显性多基因控制。
URL [本文引用: 1]
油菜是我国重要的油料作物之一,如何最大程度的提高油菜的产量和实现油菜的机械化生产是油菜育种和栽培的主要目标。通过对油菜株型相关性状的遗传规律和QTL定位进行研究分析,可以为这个目标的实现提供了理论依据。本文采用松散型甘蓝型油菜S33B和紧凑型甘蓝型油菜86-5-10-17组合杂交所得的F2群体为对象,探讨了与油菜机械化收获紧密相关的包括株高、分枝高度、分枝数和分枝角度4个性状的发育动态及遗传规律,并在构建遗传连锁图谱的基础上进行了4个性状的QTL定位。主要研究结果如下: 1.根据分枝角度的大小将甘蓝型油菜的株型分为紧凑型、中间型和分散型。这三种类型油菜的株高、分枝高度和分枝数的发育动态都不相同,但是每个性状的发育动态变化趋势几乎是一致的。从初花期到盛花期是油菜株高、分枝高度和分枝数快速生长期,在这四个生育期内株高、分枝数都是中间型株型松散型株型紧凑型株型;分枝高度则是松散株型中间型株型紧凑型株型。 2.在紧凑型株型中,第一分枝角度和第三分枝角都呈先缓下降后上升最后快速下降的趋势,第二分枝角度呈先上升后下降的趋势,第四分枝角度在整个生育期内都是呈直线缓慢下降的趋势;在中间型油菜株型中,第二分枝角度和第四分枝角度变化趋势是一致的,都是先下降后上升的的趋势,第一分枝角度呈先下降后上升最后再下降的趋势,第三分枝角度呈不断上升趋势;在松散型油菜株型中,第一个分枝角度,在整个生育期内都呈不断增加趋势,第二个分枝角度呈先下降后上升的趋势,从第三个分枝角呈先缓慢下降后上升的趋势第四分枝角度呈先下降后上升的趋势。 3.甘蓝型油菜株高和分枝数的F1正反交均值都超过了亲本,说明株高和分枝数F1具有很好的杂种优势;分枝角度的F1正反交均值都介于双亲之间,都偏向于亲本S33B,说明控制分枝角度的基因是以加性效应为主,有部分显性效应,分散型对紧凑型为部分显性;分枝高度的F1正交均值比亲本的要大,F1反交均值介于双亲之间,偏向于亲本86-5-10-17,说明分枝高度可能存在一定的胞质效应。 4.甘蓝型油菜的株高符合两对主基因控制的等显性模型(B-6模型),这两个主基因的显性效应相等并与相应的加性效应也相等,无上位性效应存在;分枝高度是一对主基因控制的加性和部分显性或超显性模型(A-1模型):分枝数符合两对主基因控制的等加性模型(B-4模型),两对主基因的加性效应相等且无显性效应和上位性效应。分枝角度平均数符合加性-显性-上位性模型(B-1模型),有两个主基因控制的株高,分枝高度,分枝数,和分枝角度的主基因遗传率分别为:44.71%,64.43%,4.55%,60.52%。多基因遗传率分别为:1.36%,8.34%,0.56%,0.95%。都以主基因遗传为主。 5.用223个SSR多态性标记构建了包含了13个连锁群163个标记位点的F2家系群体连锁图谱,有60个标记位点没有进入任何连锁群,占总数的26.9%。该遗传连锁图谱总长度为906.986cM。最大的连锁群有63个标记,最小的只有2个标记,标记间平均距离为12.286cM。其中超过100cM的有3个连锁群,分别是LG1、LG2和LG8。其中LG1距离最长,为177.222cM,LG13距离最短,仅为9.481cM。平均间距最大的为LG11,达30.968cM;平均间距最小的为LG1,只有2.858cM。 6.共检测到11个与这四个株型性状相关的QTLs,分别位于LG1、LG2、 LG3、LG5、LG8连锁群上,且都具有较大的遗传效率。其中株高4个,分别位于LG1、LG2连锁群上,可解释表型变异的8.54%-17.04%;分枝高度5个,分别位于LG3、LG5、LG8连锁群上,可解释表型变异的3.92%-21.82%;分枝数1个,位于LG3连锁群上,可解释表型变异的11.08%;分枝角度1个,位于LG1连锁群上,可解释表型变异的14.16%。只有分枝高度的第二个QTL、分枝数和分枝角度QTL的增效基因都来源于松散型亲本S33B,其余QTL的增效基因都来源于紧凑型亲本86-5-10-17。油菜株型的初步定位结果为该基因的精细定位、分子标记辅助育种和基因克隆功能分析等奠定基础。
URL [本文引用: 1]
油菜是我国重要的油料作物之一,如何最大程度的提高油菜的产量和实现油菜的机械化生产是油菜育种和栽培的主要目标。通过对油菜株型相关性状的遗传规律和QTL定位进行研究分析,可以为这个目标的实现提供了理论依据。本文采用松散型甘蓝型油菜S33B和紧凑型甘蓝型油菜86-5-10-17组合杂交所得的F2群体为对象,探讨了与油菜机械化收获紧密相关的包括株高、分枝高度、分枝数和分枝角度4个性状的发育动态及遗传规律,并在构建遗传连锁图谱的基础上进行了4个性状的QTL定位。主要研究结果如下: 1.根据分枝角度的大小将甘蓝型油菜的株型分为紧凑型、中间型和分散型。这三种类型油菜的株高、分枝高度和分枝数的发育动态都不相同,但是每个性状的发育动态变化趋势几乎是一致的。从初花期到盛花期是油菜株高、分枝高度和分枝数快速生长期,在这四个生育期内株高、分枝数都是中间型株型松散型株型紧凑型株型;分枝高度则是松散株型中间型株型紧凑型株型。 2.在紧凑型株型中,第一分枝角度和第三分枝角都呈先缓下降后上升最后快速下降的趋势,第二分枝角度呈先上升后下降的趋势,第四分枝角度在整个生育期内都是呈直线缓慢下降的趋势;在中间型油菜株型中,第二分枝角度和第四分枝角度变化趋势是一致的,都是先下降后上升的的趋势,第一分枝角度呈先下降后上升最后再下降的趋势,第三分枝角度呈不断上升趋势;在松散型油菜株型中,第一个分枝角度,在整个生育期内都呈不断增加趋势,第二个分枝角度呈先下降后上升的趋势,从第三个分枝角呈先缓慢下降后上升的趋势第四分枝角度呈先下降后上升的趋势。 3.甘蓝型油菜株高和分枝数的F1正反交均值都超过了亲本,说明株高和分枝数F1具有很好的杂种优势;分枝角度的F1正反交均值都介于双亲之间,都偏向于亲本S33B,说明控制分枝角度的基因是以加性效应为主,有部分显性效应,分散型对紧凑型为部分显性;分枝高度的F1正交均值比亲本的要大,F1反交均值介于双亲之间,偏向于亲本86-5-10-17,说明分枝高度可能存在一定的胞质效应。 4.甘蓝型油菜的株高符合两对主基因控制的等显性模型(B-6模型),这两个主基因的显性效应相等并与相应的加性效应也相等,无上位性效应存在;分枝高度是一对主基因控制的加性和部分显性或超显性模型(A-1模型):分枝数符合两对主基因控制的等加性模型(B-4模型),两对主基因的加性效应相等且无显性效应和上位性效应。分枝角度平均数符合加性-显性-上位性模型(B-1模型),有两个主基因控制的株高,分枝高度,分枝数,和分枝角度的主基因遗传率分别为:44.71%,64.43%,4.55%,60.52%。多基因遗传率分别为:1.36%,8.34%,0.56%,0.95%。都以主基因遗传为主。 5.用223个SSR多态性标记构建了包含了13个连锁群163个标记位点的F2家系群体连锁图谱,有60个标记位点没有进入任何连锁群,占总数的26.9%。该遗传连锁图谱总长度为906.986cM。最大的连锁群有63个标记,最小的只有2个标记,标记间平均距离为12.286cM。其中超过100cM的有3个连锁群,分别是LG1、LG2和LG8。其中LG1距离最长,为177.222cM,LG13距离最短,仅为9.481cM。平均间距最大的为LG11,达30.968cM;平均间距最小的为LG1,只有2.858cM。 6.共检测到11个与这四个株型性状相关的QTLs,分别位于LG1、LG2、 LG3、LG5、LG8连锁群上,且都具有较大的遗传效率。其中株高4个,分别位于LG1、LG2连锁群上,可解释表型变异的8.54%-17.04%;分枝高度5个,分别位于LG3、LG5、LG8连锁群上,可解释表型变异的3.92%-21.82%;分枝数1个,位于LG3连锁群上,可解释表型变异的11.08%;分枝角度1个,位于LG1连锁群上,可解释表型变异的14.16%。只有分枝高度的第二个QTL、分枝数和分枝角度QTL的增效基因都来源于松散型亲本S33B,其余QTL的增效基因都来源于紧凑型亲本86-5-10-17。油菜株型的初步定位结果为该基因的精细定位、分子标记辅助育种和基因克隆功能分析等奠定基础。
URL [本文引用: 1]
【目的】优质高产豆腐与豆乳专用品种的选育是现代大豆品质育种的重要方向,本研究欲通过对大豆同一重组自交系群体2004和2005两年的豆腐与豆乳得率进行相关的遗传分析与QTL定位,为豆腐与豆乳专用品种选育提供遗传学依据。【方法】以干豆腐与干豆乳得率均差异极显著的大豆品种科丰1号与南农1138-2及其构建的184个重组自交系的群体为试验材料,应用主基因+多基因混合遗传模型进行遗传分析;以该群体所构建,由488个分子标记组成,覆盖4226.40cM,平均图距8.66cM的遗传连锁图谱为基础,应用软件CartographerV2.5的复合区间作图(CIM)程序检测QTL。【结果】两个年份两个性状均存在双向超亲变异,年份间、群体各家系间、以及年份与家系互作间的差异均极显著;干豆腐得率的遗传,两个年份及两年平均值均属两对具有累加作用的连锁主基因加多基因混合遗传模型,重组率均为0.00,主基因遗传率为13.23%~26.84%,多基因遗传率为73.15%~86.77%;各年份及两年平均干豆乳得率的遗传均为两对连锁主基因加多基因混合遗传模型,重组率均为0.00,主基因遗传率为17.27%~22.29%,多基因遗传率为77.71%~82.73%。CIM检测的QTL结果显示,在C2连锁群STAS815T~A676I标记区间检测到与干豆腐得率相关的2个紧密连锁的QTL,能在不同年份稳定表达,对表型变异的贡献率累计为16.23%~23.18%;在M连锁群satt728~K24I标记区间定位到1个控制干豆乳得率的QTL,在不同年份稳定表达,距离其左侧标记0.01cM,对表型变异的贡献率为4.73%~7.14%。【结论】豆腐与豆乳得率均属主基因加多基因遗传,主基因遗传贡献不大,多基因占主要部分(≥73.15%),遗传分析和QTL定位的结果可以相互验证,遗传改良需要更多地依靠多基因积聚。
URL [本文引用: 1]
【目的】优质高产豆腐与豆乳专用品种的选育是现代大豆品质育种的重要方向,本研究欲通过对大豆同一重组自交系群体2004和2005两年的豆腐与豆乳得率进行相关的遗传分析与QTL定位,为豆腐与豆乳专用品种选育提供遗传学依据。【方法】以干豆腐与干豆乳得率均差异极显著的大豆品种科丰1号与南农1138-2及其构建的184个重组自交系的群体为试验材料,应用主基因+多基因混合遗传模型进行遗传分析;以该群体所构建,由488个分子标记组成,覆盖4226.40cM,平均图距8.66cM的遗传连锁图谱为基础,应用软件CartographerV2.5的复合区间作图(CIM)程序检测QTL。【结果】两个年份两个性状均存在双向超亲变异,年份间、群体各家系间、以及年份与家系互作间的差异均极显著;干豆腐得率的遗传,两个年份及两年平均值均属两对具有累加作用的连锁主基因加多基因混合遗传模型,重组率均为0.00,主基因遗传率为13.23%~26.84%,多基因遗传率为73.15%~86.77%;各年份及两年平均干豆乳得率的遗传均为两对连锁主基因加多基因混合遗传模型,重组率均为0.00,主基因遗传率为17.27%~22.29%,多基因遗传率为77.71%~82.73%。CIM检测的QTL结果显示,在C2连锁群STAS815T~A676I标记区间检测到与干豆腐得率相关的2个紧密连锁的QTL,能在不同年份稳定表达,对表型变异的贡献率累计为16.23%~23.18%;在M连锁群satt728~K24I标记区间定位到1个控制干豆乳得率的QTL,在不同年份稳定表达,距离其左侧标记0.01cM,对表型变异的贡献率为4.73%~7.14%。【结论】豆腐与豆乳得率均属主基因加多基因遗传,主基因遗传贡献不大,多基因占主要部分(≥73.15%),遗传分析和QTL定位的结果可以相互验证,遗传改良需要更多地依靠多基因积聚。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}