关键词:糯玉米; opaque2基因; waxy基因; 近等基因系; 赖氨酸 Construction of Waxy Maize opaque2 Near-isogenic Lines ZHANG Xiao-Xing1, ZHU Hui1, ZHANG Dong-Min1, SONG Li-Ya2, ZHANG De-Gui1, WENG Jian-Feng1, HAO Zhuan-Fang1, LI Ming-Shun1,* 1Institute of Crop Science, Chinese Academy of Agricultural Sciences / Key Laboratory of Crop Genetics and Breeding, Ministry of Agriculture, Beijing 100081, China
2Beijing Technology and Business University / Beijing Key Laboratory of Plant Resources Research and Development, Beijing 100048, China
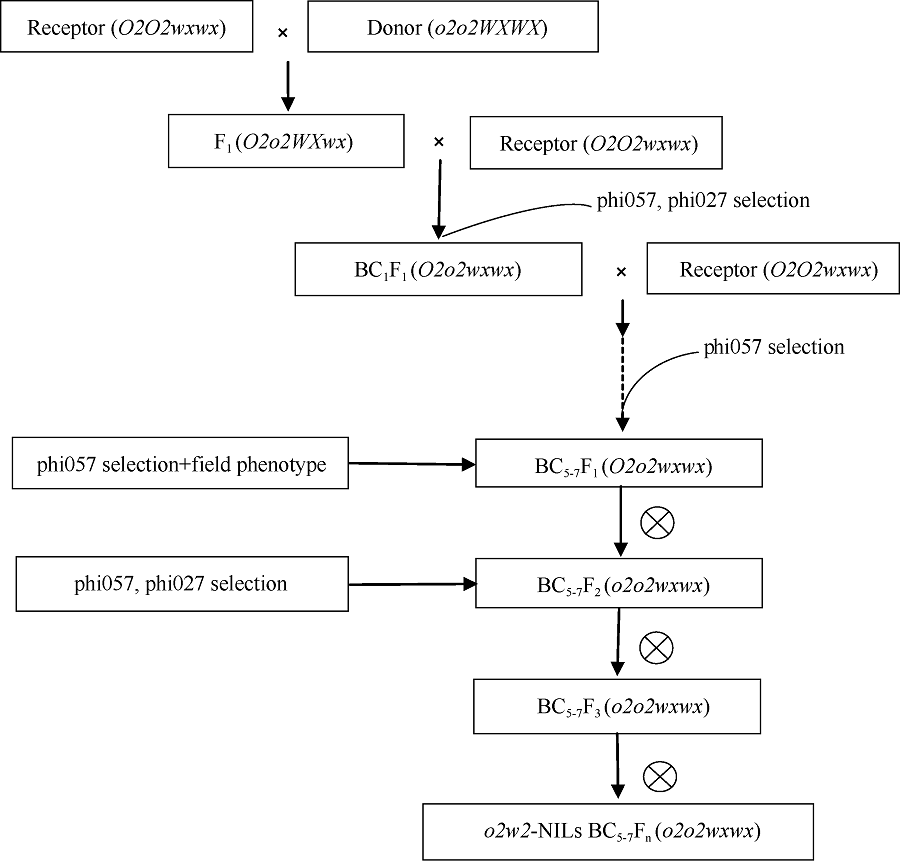
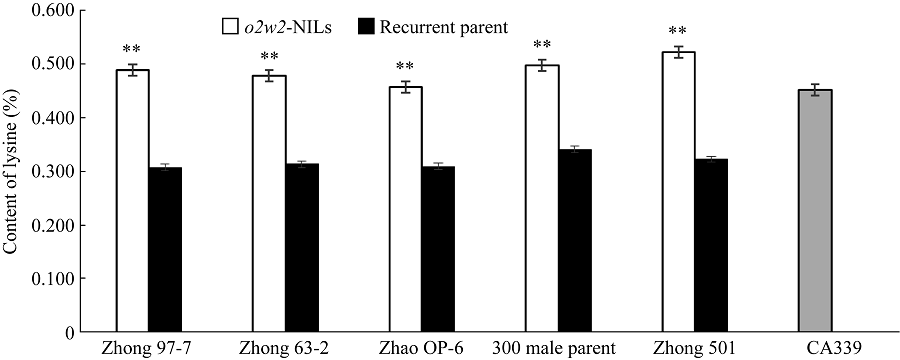
Fund:This study was supported by the National Natural Science Foundation of China (31401390) and the China Agriculture Research System (CAR02-01) AbstractIn waxy maize, the opaque2 gene can increase the content of essential amino acids, such as lysine and tryptophan. Two high quality protein maize (QPM) inbred lines, CA339 and Lu 2548, were used as opaque2 gene donors. Twenty-five waxy maize with advantageous traits were used as receptors. Application of backcrossing and SSR marker-assisted selection in breeding can create waxy maize opaque2 near-isogenic lines. The results of molecular marker indicated that there were polymorphisms in different receptors and between the two donors. Lysine content of five sets of successfully created opaque2 near-isogenic lines was tested, showing that the content of lysine in o2w-NILs was 59.0%, 52.7%, 48.5%, 46.3%, and 61.9% higher than that in their recurrent parents, which was increased from 0.308%, 0.313%, 0.309%, 0.341%, 0.323% to 0.489%, 0.478%, 0.458%, 0.498%, and 0.522%, respectively. Using this method, opaque2 could be introgressed into multiple receptors with different genetic backgrounds to select near-isogenic lines with high lysine content and transparent phenotype, so as to improve nutritional and economic values of waxy maize.
Keyword:Waxy maize; Opaque2 gene; Waxy gene; Near-isogenic lines; Lysine Show Figures Show Figures
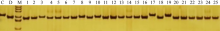
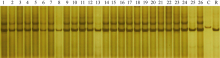
图2 引物phi057检测供体和受体亲本间的遗传多态性 C: CA339; D: 鲁2548; M: DNA marker; 1~25: 25个糯玉米自交系。Fig. 2 Polymorphism analysis of SSR primer phi057 between donors and receptors C: CA339; D: Lu 2548; M: DNA marker; 1-25: 25 waxy maize inbred lines.
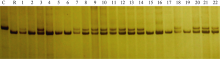
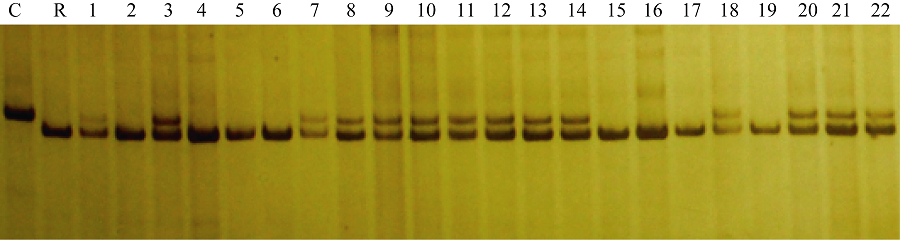
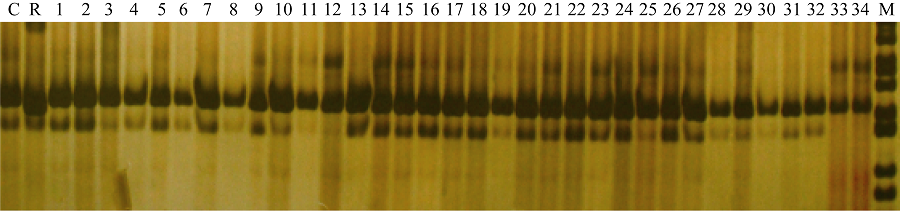
图5 引物phi057检测BC5F3世代基因型 C: CA339; R: 轮回亲本; 1~34: BC5F3世代不同单株; M: DNA marker。Fig. 5 Genotype identification of waxy maize in BC5F3family with primer phi057 C: CA339; R: Recurrent parent; 1-34: Different individuals from BC5F3family; M: DNA marker.
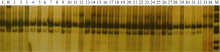
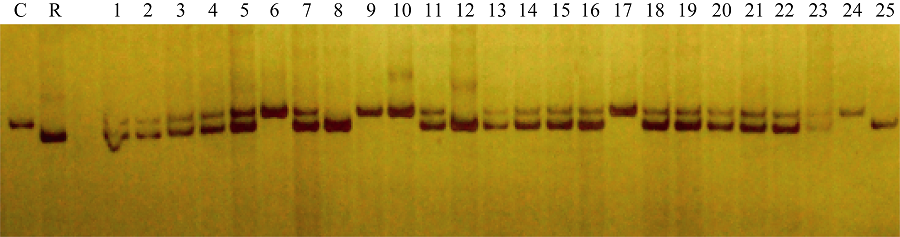
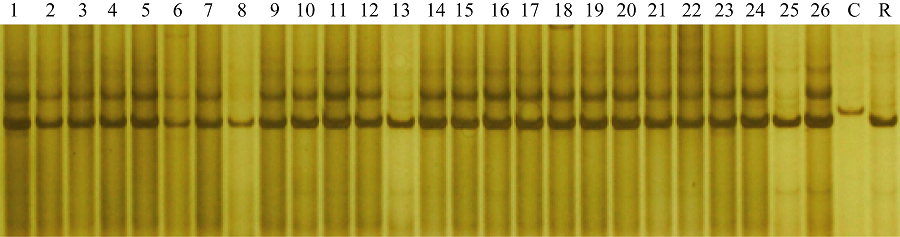
图6 引物phi027检测BC5F3世代基因型 1~26: BC5F3世代不同单株; C: CA339; R: 轮回亲本。Fig. 6 Genotype identification of waxy maize in BC5F3family with primer phi027 1-26: Different individuals from BC5F3family; C: CA339; R: Recurrent parent.
Briggs RW, AmanoE, Smith HH. Genetic recombination with ethylmethane sulphonate induced Waxy mutants in maize. Nature, 1965, 207: 890-891[本文引用:1]
[2]
Wessler SR, TarpleyA, PuruggananM, SpellM, OkagakiR. Filler DNA is associated with spontaneous deletions in maize. Proc Natl Acad Sci USA, 1990, 87: 8731-8735[本文引用:1]
[3]
彭泽斌, 田志国. 我国糯玉米产业现状与发展战略. 玉米科学, 2004, 12(3): 116-118Peng ZB, Tian ZG. Industrialization present status and strategies for development of waxy corn in China. J Maize Sci, 2004, 12(3): 116-118 (in Chinese with English abstract)[本文引用:1]
[4]
杨引福, 郭强, 陈婧, 郑小亚, 蔺崇明. 中国温带糯玉米自交系遗传品质性状分析. 西北植物学报, 2009, 29: 2213-2220Yang YF, GuoQ, ChenJ, Zheng XY, Lin CM. Analysis of genetic and quality traits of waxy corn inbred lines in China temperate zone. Acta Bot Boreali-Occident Sin, 2009, 29: 2213-2220 (in Chinese with English abstract)[本文引用:1]
[5]
田清震, 李新海, 李明顺, 姜伟, 张世煌. 优质蛋白玉米的分子标记辅助选择. 玉米科学, 2004, 12(2): 108-110Tian QZ, Li XH, Li MS, JiangW, Zhang SH. Molecular markers assisted selection to quality protein maize. J Maize Sci, 2004, 12(2): 108-110 (in Chinese with English abstract)[本文引用:1]
[6]
Mertz ET, Bates LS, Nelson OE. Mutant gene that changes protein composition and increases lysine content of maize endosperm. Science, 1964, 145: 270-280[本文引用:1]
[7]
HartingsH, MaddaloniM, LazzaroniN, Fonzo ND, MottoM, SalaminiF, ThompsonR. The O2 gene which regulates zein deposition in maize endosperm encodes a protein with structural homologies to transcriptional activators. EMBO J, 1989, 8: 2795-2801[本文引用:1]
[8]
MaddaloniM, Fonzo ND, HartingsH, LazzaroniN, SalaminiF, ThompsonR, MottoM. The sequence of the zein regulatory gene opaque-2 (O2) of Zea mays. Nucl Acids Res, 1989, 17: 7532[本文引用:1]
[9]
Schmidt RJ, Burr FA, Aukerman MJ, BurrB. Maize regulatory gene opaque-2 encodes a protein with a “leucine-zipper” motif that binds to zein DNA. Proc Natl Acad Sci USA, 1990, 87: 46-50[本文引用:1]
[10]
Schmidt RJ, KetudatM, Aukerman MJ, HoschekG. Opaque-2 is a transcriptional activator that recognizes a specific target site in 22-kD zein genes. Plant Cell, 1992, 4: 689-700[本文引用:1]
[11]
SchmitzD, LohmerS, SalaminiF, Thompson RD. The activation domain of the maize transcription factor Opaque-2 resides in a single acidic region. Nucl Acids Res, 1997, 25: 756-763[本文引用:1]
ViottiA, SalaE, MarottaR, AlberiP, BalducciC, SoaveC. Genes and mRNAs coding for zein polypeptides in Zea mays. Eur J Biochem, 1979, 102: 211-222[本文引用:1]
[14]
Crow JF, KermicleJ. Oliver Nelson and quality protein maize. Genetics, 2002, 160: 819-821[本文引用:1]
[15]
Vasal SK. High quality protein corn. In: Hallawer A R ed. Specialty Corns. Boca Raton: CRC Press, 2001. pp 85-121[本文引用:1]
[16]
宋敏, 张世煌. 优质蛋白玉米(QPM)育种研究进展. 新疆农业科学, 2007, 44(增刊3): 30-35SongM, Zhang SH. Comprehensive report on study advance of breeding of high-quality protein maize (QPM). Xinjiang Agric Sci, 2007, 44(suppl-3): 30-35 (in Chinese)[本文引用:1]
[17]
ZhangW, YangW, WangM, WangW, ZengG, ChenZ, CaiY. Increasing lysine content of waxy maize through introgression of Opaque-2 and Opaque-16 genes using molecular assisted and biochemical development. PLoS One, 2013, 8: e56227[本文引用:1]
[18]
昂沃, 郝小琴, 吴子恺. Opaque2基因导入各类隐性纯合体玉米早代籽粒赖氨酸含量的效应研究. 玉米科学, 2012, 20(2): 56-58AngW, Hao XQ, Wu ZK. Analysis of Opaque-2 gene effects on kernel lysine content into some early generations’ homozygote recessive maize. J Maize Sci, 2012, 20(2): 56-58 (in Chinese with English abstract)[本文引用:1]
[19]
卢振宇, 李明顺, 谢传晓, 李新海, 曲延英, 张世煌. 玉米叶片DNA快速提取方法改进研究. 玉米科学, 2008, 16(2): 50-53Lu ZY, Li MS, Xie CX, Li XH, Qu YY, Zhang SH. An improved study on rapid DNA extraction method from young leaves of maize. J Maize Sci, 2008, 16(2): 50-53 (in Chinese with English abstract)[本文引用:1]
[20]
Vasal SK. The quality protein maize story. Food Nutr Bull, 2000, 21: 445-450[本文引用:1]
[21]
Chin EC, Senior ML, ShuH, Smith JS. Maize simple repetitive DNA sequences: abundance and allele variation. Genome, 1996, 39: 866-873[本文引用:1]
[22]
ChenY, ZhouZ, ZhaoG, LiX, SongL, YanN, WengJ, HaoZ, ZhangD, LiM, ZhangS. Transposable element rbg induces the differential expression of opaque-2 mutant gene in two maize o2 NILs derived from the same inbred line. PLoS One, 2014, 9: e85159[本文引用:1]
[23]
赵刚, 吴子恺, 陈亮, 张德贵, 张世煌, 卢振宇, 白丽, 李明顺. 几个供体对优质蛋白玉米(QPM)近等基因系构建效果的比较. 玉米科学, 2010, 18(6): 11-14ZhaoG, Wu ZK, ChenL, Zhang DG, Zhang SH, Lu ZY, BaiL, Li MS. Comparison of several donor inbred lines on development of quality protein maize (QPM) near-isogenic lines. J Maize Sci, 2010, 18(6): 11-14 (in Chinese with English abstract)[本文引用:1]
 , 朱慧
, 朱慧




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}