关键词:水稻; 蜡质基因; 四引物扩增受阻突变体系; 直链淀粉含量; 稻米食味品质改良 Development of PCR Functional Markers for Multiple Alleles of Wx and Their Application in Rice MAO Ting1,2, LI Xu2, LI Zhen-Yu2,*, XU Zheng-Jin1,* 1Rice Research Institute, Shenyang Agricultural University, Shenyang 110866, China
2Liaoning Province Saline and Alkaline Land Utilization and Research Institute, Panjin 124010, China
Fund:This study was supported by the Cultivation Plan for Youth Agricultural Science Technology Innovative Talents of Liaoning Province (2015035, 2015036) and the Natural Science Foundation of China (31371587, 31430062) AbstractAmylose content (AC) is one of the most important factors impacting rice eating quality, which was mainly determined by Wx locus. Multiple alleles, such as Wxa, Wxb, Wxin, and Wxmw contribute to the variation of AC in rice varieties. Selecting rice varieties with low-medium AC through Marker-assisted selection (MAS) is an important way to improve rice eating quality. However, distinguishing the Single Nucleotide Polymorphisms (SNP) of Wx alleles requires the DNA sequencing or restriction enzyme digestion, which is time consuming and labor-intensive. In this study, we developed effective MAS markers by tetra-primer ARMS-PCR technology utilizing functional SNPs corresponding to the different alleles, moreover we put forward corresponding solving schemes on low amplification efficiency and inaccuracy extension through adjusting the location of deliberate mismatch bases introduced. We easily differentiated the multiple alleles of Wx utilizing the markers Waxygt-ARMS2 and Waxyac-ARMS2, showing that these markers have significant application value with the features of rapid operation and low cost. We further analyzed the genotype distribution of Wx allele in Liaoning Province rice varieties, and found all of the forty tested varieties carried the Wxb allele with a narrow genetic diversity. The results of this study could provide theoretical basis and technical support on both new rice varieties breeding and rice eating quality improvement utilizing Wx alleles corresponding to low-medium AC by MAS in Liaoning Province.
Keyword:Rice; Wx; Tetra-primer ARMS-PCR; Amylose content; Rice eating quality improvement Show Figures Show Figures
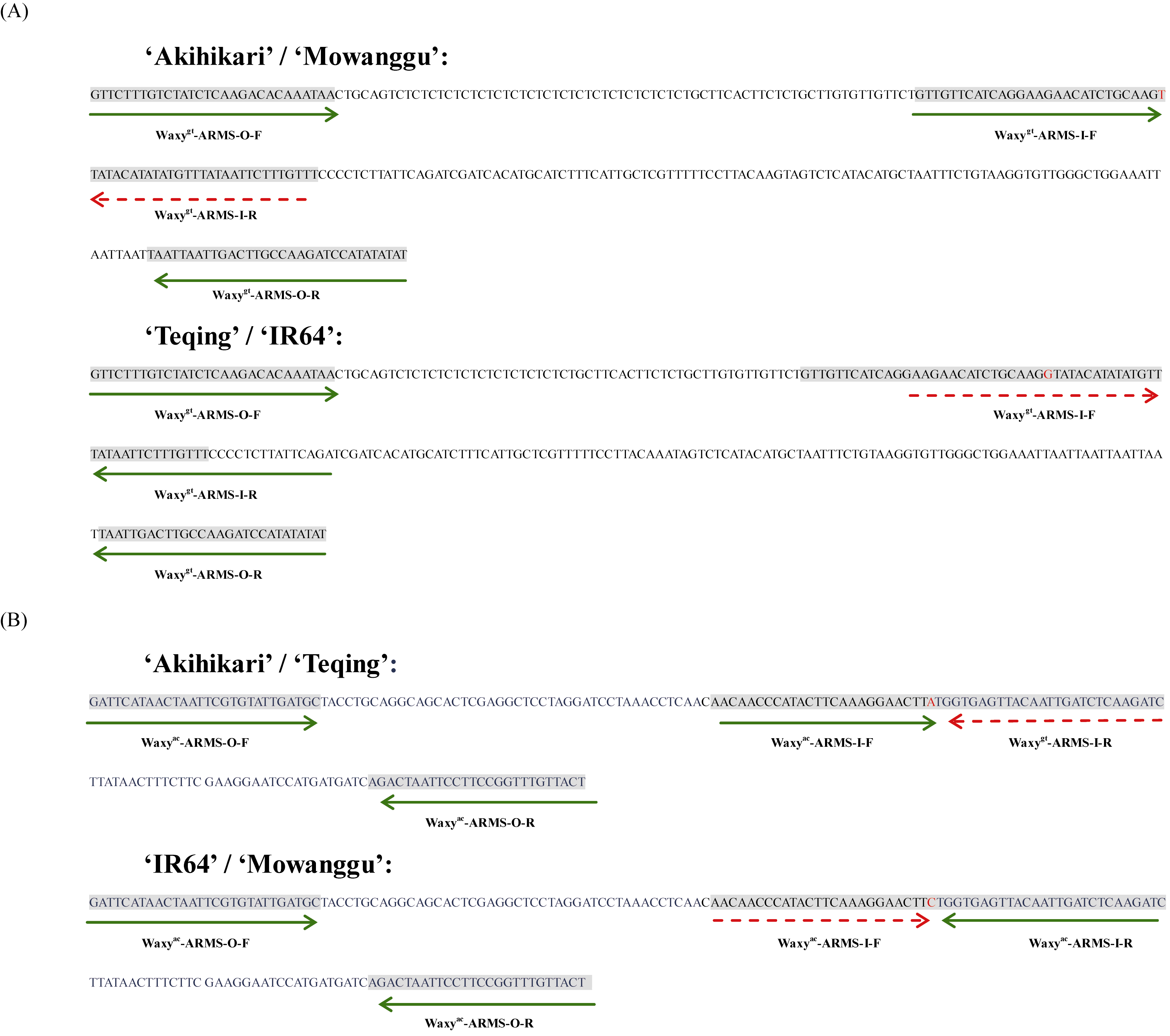
图1 引物设计策略 A为SNP1引物设计策略, B为SNP2引物设计策略, 功能性SNP多态性位点用红色表示, 添加底纹背景的碱基表示引物结合位点, 绿色实线箭头表示引物扩增方向, 红色虚线箭头表示不能有效扩增。Fig. 1 Strategy of primer design A:design strategies for SNP1; B:design strategies for SNP2, functional SNPs are shown in red, gray background shows primers’ binding site, the green full arrows indicate the primers’ amplification direction, the red short dash arrows indicate the primer could not amplify effectively.
1.4 DNA提取、PCR扩增及电泳检测全基因组DNA选用全式金生物技术有限公司的植物DNA提取试剂盒提取(具体操作按照产品说明)。 20 μ L PCR体系包括:DNA 0.5 μ L (50 ng μ L-1), Primer 2.0 μ L (4 pmol μ L-1, 内外引物浓度比为1.5∶ 1.0), 2 × PCR Master Mix 10.0 μ L (带染料, 北京康为世纪生物科技有限公司), ddH2O 7.5 μ L。 反应程序为:95℃预变性5 min; 然后95℃变性30 s、56℃复性30 s (表2)、72℃延伸30 s, 30次循环; 72℃延伸10 min; 4℃冷却10 min后, 反应产物在3%或5%琼脂糖凝胶上100 V电泳1 h; 经溴化乙锭(ethidium bromide, EB)染色后于凝胶成像系统下观察, 记录。 表2 Table 2 表2(Table 2)
表2 设计引物基本信息 Table 2 Molecular markers desigaed in this study
表2 设计引物基本信息 Table 2 Molecular markers desigaed in this study
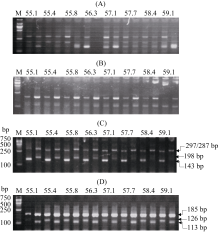
图2 候选标记在不同退火温度下的表现 M为marker 2000。A为Waxygt-ARMS1在不同退火温度下的表现, 在每一退火温度组, 前泳道为秋光, 后泳道为特青, B为Waxyac-ARMS1在不同退火温度下的表现, 在每一退火温度组, 前泳道为魔王谷, 后泳道为秋光, C为Waxygt-ARMS2在不同退火温度下的表现, 在每一退火温度组, 前泳道为秋光, 后泳道为特青, D为Waxyac-ARMS2在不同退火温度下的表现, 在每一退火温度组, 前泳道为魔王谷, 后泳道为秋光。Fig. 2 Molecular detections on the control varieties between different annealing temperatures M indicates marker 2000. A:PCR product on different annealing temperature (° C) using the Waxygt-ARMS1 marker, on every groups of annealing temperatures, the former lane is ‘ Akihikari’ , later lane is ‘ Teqing’ . B:PCR product on different annealing temperature (° C) using the Waxyac-ARMS1 marker, on every groups of annealing temperatures, the former lane is ‘ Mowanggu’ , later lane is ‘ Akihikari’ . C:PCR product on different annealing temperature (° C) using the Waxygt-ARMS2 marker, on every groups of annealing temperatures, the former lane is ‘ Akihikari’ , later lane is ‘ Teqing’ . D:PCR product on different annealing temperature (° C) using the Waxyac-ARMS2 marker, on every groups of annealing temperatures, the former lane is ‘ Mowanggu’ , later lane is ‘ Akihikari’ .
图3 利用Waxygt-ARMS2和Waxyac-ARMS2对对照品种进行基因型分析 M为marker 2000。A为利用Waxygt-ARMS标记分析结果, B为利用Waxyac-ARMS标记分析结果; 5条泳道由左至右分别为marker 2000、魔王谷、IR 64、秋光和特青。Fig. 3 Genotype analysis on control varieties utilizing Waxygt-ARMS2and Waxyac-ARMS2 M indicates marker 2000. A:PCR product of Waxygt-ARMS2 marker. B:PCR product of Waxyac-ARMS2 marker. Lanes from left to right are marker 2000, Mowanggu, IR64, Akihikari, and Teqing.
Li YB, Fan CC, Xing YZ, YunP, Luo LJ, YanB, PengB, Xie WB, Wang GW, Li XH, Xiao JH, Xu CG, He YQ. Chalk5 encodes a vacuolar H+-translocating pyrophosphatase influencing grain chalkiness in rice. Nat Genet, 2014, 46:398-404[本文引用:1]
[2]
Zeng DL, Tian ZX, Rao YC, Dong GJ, Yang YL, Huang LC, Leng YJ, XuJ, SunC, Zhang GH, HuJ, ZhuL, Gao ZY, Hu XM, Guo LB, Xiong GS, Wang YH, Li JY, QianQ. Rational design of high-yield and superior-quality rice. Nat Plants, 2017, 3:17031[本文引用:2]
[3]
吴长明, 孙传清, 付秀林, 王象坤, 李自超, 张强. 稻米品质性状与产量性状及籼粳分化度的相互关系研究. 作物学报, 2003, 29:822-828Wu CM, Sun CQ, Fu XL, Wang XK, Li ZC, ZhangQ. Study on the relationship between quality, yield characters or indica- japonica differentiation in rice (Oryza sativa L. ). Acta Agron Sin, 2003, 29:822-828 (in Chinese with English abstract)[本文引用:1]
[4]
王才林, 陈涛, 张亚东, 朱镇, 赵凌, 林静. 通过分子标记辅助选择培育优良食味水稻新品种. 中国水稻科学, 2009, 23:25-30Wang CL, ChenT, Zhang YD, ZhuZ, ZhaoL, LinJ. Breeding of a new rice variety with good eating quality by marker assisted selection. Chin J Rice Sci, 2009, 23:25-30 (in Chinese with English abstract)[本文引用:2]
[5]
隋炯明, 李欣, 严松, 严长杰, 张蓉, 汤述翥, 陆驹飞, 陈宗祥, 顾铭洪. 稻米淀粉 RVA 谱特征与品质性状相关性研究. 中国农业科学, 2005, 38:657-663Sui JM, LiX, YanS, Yan CJ, ZhangR, Tang SZ, Lu JF, Chen ZX, Gu MH. Studies on the rice RVA profile characteristics and its correlation with the quality. Sci Agric Sin, 2005, 38:657-663 (in Chinese with English abstract)[本文引用:1]
[6]
SanoY. Differential regulation of waxy gene expression in rice endosperm. Theor Appl Genet, 1984, 68:467-473[本文引用:1]
[7]
Ayres NM, McClung A M, Larkin P D, Bligh H F J, Jones C A, Park W D. Microsatellites and a single-nucleotide polymorphism differentiate apparent amylose classes in an extended pedigree of US rice germ plasm. Theor Appl Genet, 1997, 94:773-781[本文引用:1]
[8]
MikamiI, AikawaM, Hirano HY, SanoY. Altered tissue-specific expression at the Wx gene of the opaque mutants in rice. Euphytica, 1999, 105:91-97[本文引用:1]
[9]
Wang ZY, Zheng FQ, Shen GZ, Gao JP, Snustad DP, Li MG, Zhang JL, Hong MM. The amylose content in rice endosperm is related to the post-transcriptional regulation of the waxy gene. Plant J, 1995, 7:613-622[本文引用:2]
[10]
YamanakaS, NakamuraI, Watanabe KN, Sato YI. Identification of SNPs in the waxy gene among glutinous rice cultivars and their evolutionary significance during the domestication process of rice. Theor Appl Genet, 2004, 108:1200-1204[本文引用:1]
[11]
朱霁晖, 张昌泉, 顾铭洪, 刘巧泉. 水稻Wx基因的等位变异及育种利用研究进展. 中国水稻科学, 2015, 29:431-438Zhu JH, Zhang CQ, Gu MH, Liu QQ. Progress in the allelic variation of Wx gene and its application in rice breeding. Chin J Rice Sci, 2015, 29:431-438 (in Chinese with English abstract)[本文引用:5]
[12]
MikamiI, UwatokoN, IkedaY, YamaguchiJ, Hirano HY, SuzukiY, SanoY. Allelic diversification at the wx locus in land races of Asian rice. Theor Appl Genet, 2008, 116:979-989[本文引用:1]
[13]
倪晖. 水稻Wx基因不同等位变异的效应及其育种应用的初步研究. 扬州大学硕士学位论文, 江苏扬州, 2013NiH. A Preliminary Analysis of the Effect of different Wx Alleles on Rice Grain Quality. MS Thesis of Yangzhou University, Yangzhou, China, 2013 (in Chinese with English abstract)[本文引用:9]
[14]
刘巧泉, 蔡秀玲, 李钱锋, 汤述翥, 龚志云, 于恒秀, 严长杰, 王宗阳, 顾铭洪. 分子标记辅助选择改良特青及其杂交稻米的蒸煮与食味品质. 作物学报, 2006, 32:64-69Liu QQ, Cai XL, Li QF, Tang SZ, Gong ZY, Yu HX, Yan CJ, Wang ZY, Gu MH. Molecular marker-assisted selection for improving cooking and eating quality in Teqing and its hybrid rice. Acta Agron Sin, 2006, 32:64-69 (in Chinese with English abstract)[本文引用:3]
[15]
蔡秀玲, 刘巧泉, 汤述翥, 顾铭洪, 王宗阳. 用于筛选直链淀粉含量为中等的籼稻品种的分子标记. 植物生理与分子生物学报, 2002, 28:137-144Cai XL, Liu QQ, Tang SZ, Gu MH, Wang ZY. Development of a molecular marker for screening the rice cultivars with intermediate amylose content in Oryza sativa subsp. indica. J Plant Physiol Mol Biol, 2002, 28:137-144 (in Chinese with English abstract)[本文引用:6]
[16]
姚姝, 陈涛, 张亚东, 朱镇, 赵凌, 赵庆勇, 周丽慧, 王才林. 分子标记辅助选择聚合水稻暗胚乳突变基因Wx-mq和抗条纹叶枯病基因Stv-bi. 中国水稻科学, 2010, 24:341-347YaoS, ChenT, Zhang YD, ZhuZ, ZhaoL, Zhao QY, Zhou LH, Wang CL. Pyramiding of translucent endosperm mutant gene Wx-mq and rice stripe disease resistance gene Stv-bi by marker-assisted selection in rice (Oryza sativa). Chin J Rice Sci, 2010, 24:341-347 (in Chinese with English abstract)[本文引用:4]
[17]
田志喜, 严长杰, 钱前, 严松, 谢会兰, 王芳, 徐洁芬, 刘贵富, 王永红, 刘巧泉, 汤述翥, 李家洋, 顾铭洪. 水稻淀粉合成相关基因分子标记的建立. 科学通报, 2010, 55:2591-2601Tian ZX, Yan CJ, QianQ, YanS, Xie HL, WangF, Xu JF, Liu FG, Wang YH, Liu QQ, Tang SZ, Li JY, Gu MH. Development of gene-tagged molecular markers for starch synthesis-related genes in rice. Chin Sci Bull, 2010, 55:2591-2601 (in Chinese)[本文引用:1]
[18]
刘兴顺, 程牛亮, 梁小波, 赵彩虹. 四引物扩增受阻突变体系聚合酶链反应在SNP基因分型中的研究. 山西医科大学学报, 2008, 39:483-485Liu XS, Cheng NL, Liang XB, Zhao CH. Study on tetra-primer amplification refractory mutation system-polymerase chain reaction in SNP genotyping. J Shanxi Med Univ, 2008, 39:483-485 (in Chinese with English abstract)[本文引用:5]
[19]
YeS, DhillonS, KeX, Collins AR, Day IN. An efficient procedure for genotyping single nucleotide polymorphisms. Nucl Acids Res, 29:E88[本文引用:11]
[20]
陈涛, 骆名瑞, 张亚东, 朱镇, 赵凌, 赵庆勇, 周丽慧, 姚姝, 于新, 王才林. 利用四引物扩增受阻突变体系PCR技术检测水稻低直链淀粉含量基因Wx-mq. 中国水稻科学, 2013, 27:529-534ChenT, Luo MR, Zhang YD, ZhuZ, ZhaoL, Zhao QY, Zhou LH, YaoS, YuX, Wang CL. Detection of Wx-mq gene for low-amylose content by tetra-primer amplification refractory mutation system PCR in rice. Chin J Rice Sci, 2013, 27:529-534 (in Chinese with English abstract)[本文引用:5]
[21]
Kim SR, RamosJ, AshikariM, Virk PS, Torres EA, NissilaE, Hechanova SL, MauleonR, Jena KK. Development and validation of allele-specific SNP/indel markers for eight yield-enhancing genes using whole-genome sequencing strategy to increase yield potential of rice, Oryza sativa L. Rice, 2016, 9:12[本文引用:1]
HuB, WangW, Ou SJ, Tang JY, LiH, Che RH, Zhang ZH, Chai XY, Wang HR, Wang YQ, Liang CZ, Liu LC, Piao ZZ, Deng QY, DengK, XuC, LiangY, Zhang LH, Li LG, Chu CC. Variation in NRT1. 1B contributes to nitrate-use divergence between rice subspecies. Nat Genet, 2015, 47:834-838[本文引用:1]
[24]
潘存红, 王子斌, 马玉银, 殷跃军, 张亚芳, 左示敏, 陈宗祥, 潘学彪. InDel和SNP标记在水稻图位克隆中的应用. 中国水稻科学, 2007, 21:447-453Pan CH, Wang ZB, Ma YY, Yin YJ, Zhang YF, Zuo SM, Chen ZX, Pan XB. InDel and SNP markers and their application in map-based cloning of rice genes. Chin J Rice Sci, 2007, 21:447-453 (in Chinese with English abstract)[本文引用:1]
[25]
SunJ, LiuD, Wang JY, Ma DR, TangL, GaoH, Xu ZJ, Chen WF. The contribution of intersubspecific hybridization to the breeding of super-high-yielding japonica rice in northeast China. Theor Appl Genet, 2012, 125:1149-1157[本文引用:1]
[26]
陈温福, 徐正进, 张龙步, 张文忠, 马殿荣. 北方粳型稻超高产育种理论与实践. 中国农业科学, 2007, 40:869-874Chen WF, Xu ZJ, Zhang LB, Zhang WZ, Ma DR. Theories and practices of breeding japonica rice for super high yield. Sci Agric Sin, 2007, 40:869-874 (in Chinese with English abstract)[本文引用:2]
 , 李旭
, 李旭





{kind=link}
{kind=link}
{kind=link}