关键词:棉花; 嫁接; 根-冠互作; 下胚轴; 解剖结构 Effect of Root-shoot Interaction on Cotton Hypocotyl Anatomy by Using Grafting Technique under Premature Senescence Induced by Potassium Deficiency ZHANG Qiao-Yu1, ZHANG Rui1, LIU Zhao-Hui2,*, TIAN Xiao-Li1,* 1Center of Crop Chemical Regulation, College of Agricultural and Biotechnology, China Agricultural University, Beijing 100193, China
2Department of Plant Sciences, College of Biological Sciences, China Agricultural University, Beijing 100193, China
Fund:This study was supported by the National Natural Science Foundation of China (31271629). AbstractUnder potassium (K) deficiency, effect of root-shoot interaction on cotton leaf premature senescence includes shoot-dominated and root-dominated patterns. Indirect evidences from grafting study under K deficiency have suggested that scion hypocotyl of shoot-dominated pattern (CCRI41 and SCRC22 combination) may have the ability to synthesize, metabolize and/or radially transport cytokinin (CTK) and abscisic acid (ABA), while that of root-dominated pattern (CCRI41 and CCRI49 combination) mainly delivers CTK and ABA from roots. In the present study, we observed the anatomy of scion hypocotyls of both patterns under K deficiency (0.03 mmol L-1), and evaluated if it is associated with the flux of CTK and ABA in xylem. In terms of I type grafts (one scion grafted onto one rootstock) of shoot-dominated pattern, the anatomy of scion hypocotyl was clearly influenced by rootstocks, which is inconsistent with the fact that the delivery rate of CTK and ABA in scion xylem was mainly affected by shoot. For Y type grafts (two scions grafted onto one rootstock) of shoot-dominated pattern, the scion with greater CTK flux and lower ABA flux showed growth superiority as compared with its counterpart. However, the two processes referring to flux of phytohormone in scion xylem and scion growth likely occurred concomitantly but not causally. As the results of CTK and ABA flux in scion xylem, the anatomy of scion hypocotyls of root-dominated pattern was obviously altered by rootstock in I grafts. Nevertheless, they might be also concomitant processes rather than the relationship of cause and effect. Additionally, there were significant differences in anatomy between two different scions of Y grafts of root-dominated pattern, which was disagreement with the similar CTK and ABA flux in xylem of two scions. In conclusion, the scion hypocotyl anatomy of either shoot- or root-dominated pattern regarding cotton leaf premature senescence induced by K deficiency is independent of CTK and ABA flux in its xylem. In addition, the differences in scion hypocotyl anatomy between two cultivars in I graft type were inconsistent with those in Y graft type, which is possibly due to shoot-shoot interaction in Y graft.
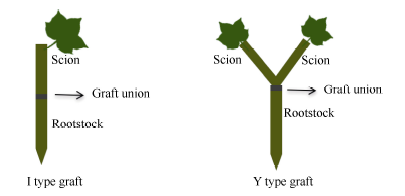
图1 I型(接穗/砧木)和Y型(接穗+接穗/砧木)嫁接示意图Fig. 1 Schema of I type graft (one scion grafted onto one rootstock) and Y type graft (two scions grafted onto one rootstock)
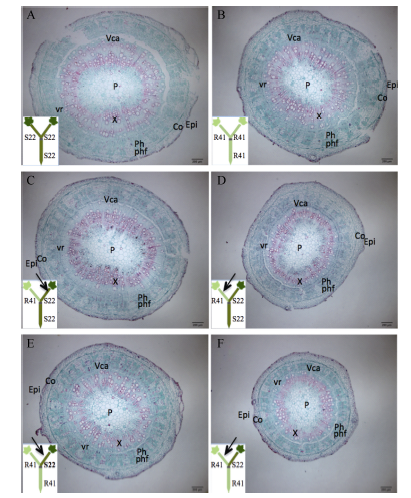
表1 棉花叶片早衰地上部主导型(中棉所41与鲁棉研22组合) I型嫁接接穗下胚轴各部分厚度及下胚轴半径 Table 1 Thickness of each tissue in cross section of scion hypocotyl and radius of scion hypocotyl in I type grafts (one scion grafted onto one rootstock, scion/rootstock) of shoot-dominated pattern (CCRI41 and SCRC22 combination) regarding cotton leaf premature senescence (μ m)
嫁接组合 Scion/rootstock
髓 Pith
木质部 Xylem
韧皮部 Phloem
表皮+皮层 Epidermis+cortex
下胚轴半径 Radius
S22/S22
330.0 b
473.9 a
249.2 a
208.5 a
1261.6 a
S22/C41
327.4 b
473.5 a
269.5 a
157.4 b
1227.8 a
C41/S22
319.3 b
447.4 a
270.1 a
166.4 ab
1203.2 a
C41/C41
428.0 a
468.1 a
243.7 a
161.5 ab
1301.3 a
S22: SCRC22; C41: CCRI41. Data within the same column followed by the same letter are not significantly different at P < 0.05 according to Duncan’ s multiple range test. S22: 鲁棉研22; C41: 中棉所41。同列数据后的不同字母表示在0.05水平差异显著。
表1 棉花叶片早衰地上部主导型(中棉所41与鲁棉研22组合) I型嫁接接穗下胚轴各部分厚度及下胚轴半径 Table 1 Thickness of each tissue in cross section of scion hypocotyl and radius of scion hypocotyl in I type grafts (one scion grafted onto one rootstock, scion/rootstock) of shoot-dominated pattern (CCRI41 and SCRC22 combination) regarding cotton leaf premature senescence (μ m)
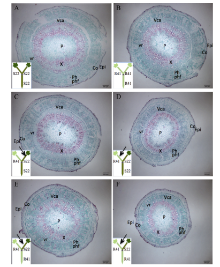
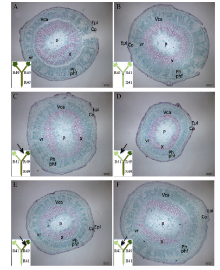
图3 棉花叶片早衰根系主导型(中棉所41与中棉所49组合) I型嫁接(接穗/砧木)接穗下胚轴解剖结构因C49/C49(A)和C41/C49(C)接穗下胚轴较粗, 为了显示全图, 标尺较其他图缩小。A, E: C49/C49; B, F: C49/C41; C, G: C41/C49; D, H: C41/C41。C49: 中棉所49; C41: 中棉所41。Epi: 表皮; Co: 皮层; P: 髓; Ph: 韧皮部; X: 木质部; vr: 维管射线; Vca: 维管形成层; phf: 韧皮纤维。Fig. 3 Anatomy of scion hypocotyl in I type grafts (one scion grafted onto one rootstock, scion/rootstock) of root-dominated pattern (CCRI41 and CCRI49 combination) regarding cotton leaf premature senescence The scale for A and C is smaller than B and D since the scion of C49/C49 (A) and C41/C49 (C) is wider in diameter. C49: CCRI49; C41: CCRI41. Epi: epidermis; Co: cortex; P: pith; Ph: phloem; X: xylem; vr: vascular rays; Vca: vascular cambium; phf: phloem fiber.
表2 棉花叶片早衰根系主导型(中棉所41与中棉所49组合)I型嫁接接穗下胚轴各部分厚度及下胚轴半径 Table 2 Thickness of each tissue in cross section of scion hypocotyl and radius of scion hypocotyl in I type grafts (one scion grafted onto one rootstock, scion/rootstock) of root-dominated pattern (CCRI41 and CCRI49 combination) regarding cotton leaf premature senescence (μ m)
嫁接组合 Scion/rootstock
髓 Pith
木质部 Xylem
韧皮部 Phloem
表皮+皮层 Epidermis+cortex
下胚轴半径 Radius
C41/C41
428.0 a
468.1 b
243.7 b
161.5 a
1301.3 a
C41/C49
372.6 ab
754.1 ab
335.9 ab
150.8 a
1613.4 a
C49/C41
316.6 b
500.6 b
258.1 b
184.7 a
1260.0 a
C49/C49
323.2 b
818.2 a
405.7 a
140.7 a
1687.8 a
C49: CCRI49; C41: CCRI41. Data within the same column followed by the same letter are not significantly different at P < 0.05 according to Duncan’ s multiple range test. C49: 中棉所49; C41: 中棉所41。同列数据后的不同字母表示在0.05水平差异显著。
表2 棉花叶片早衰根系主导型(中棉所41与中棉所49组合)I型嫁接接穗下胚轴各部分厚度及下胚轴半径 Table 2 Thickness of each tissue in cross section of scion hypocotyl and radius of scion hypocotyl in I type grafts (one scion grafted onto one rootstock, scion/rootstock) of root-dominated pattern (CCRI41 and CCRI49 combination) regarding cotton leaf premature senescence (μ m)
表3 棉花叶片早衰地上部主导型(中棉所41与鲁棉研22组合)Y型嫁接接穗下胚轴各部分厚度及下胚轴半径 Table 3 Thickness of each tissue in cross section of scion hypocotyl and radius of scion hypocotyl in Y type grafts (two scions grafted onto one rootstock, scion+scion/rootstock) of shoot-dominated pattern (CCRI41 and SCRC22 combination) regarding cotton leaf premature senescence (μ m)
嫁接组合 Scion+scion/rootstock
髓 Pith
木质部 Xylem
韧皮部 Phloem
表皮+皮层 Epidermis+cortex
下胚轴半径 Radius
(S22)+(S22)/S22
345.8 a
526.2 a
292.0 b
152.8 ab
1316.8 a
(S22)+C41/S22
333.0 ab
474.2 a
264.3 bc
172.5 a
1244.1 a
S22+(C41)/S22
295.9 abc
463.0 a
277.7 b
143.1 ab
1179.6 a
(S22)+C41/C41
266.8 c
469.2 a
285.1 b
149.0 ab
1170.2 a
S22+(C41)/C41
276.3 bc
320.9 b
211.6 c
125.3 b
934.2 b
(C41)+(C41)/C41
306.1 abc
521.3 a
354.5 a
148.1 ab
1330.0 a
S22: SCRC22; C41: CCRI41. The scion in parentheses is the measured. Data within the same column followed by the same letter are not significantly different at P< 0.05 according to Duncan’ s multiple range test. S22: 鲁棉研22; C41: 中棉所41。括号内接穗为测量接穗。同列数据后的不同字母表示在0.05水平差异显著。
表3 棉花叶片早衰地上部主导型(中棉所41与鲁棉研22组合)Y型嫁接接穗下胚轴各部分厚度及下胚轴半径 Table 3 Thickness of each tissue in cross section of scion hypocotyl and radius of scion hypocotyl in Y type grafts (two scions grafted onto one rootstock, scion+scion/rootstock) of shoot-dominated pattern (CCRI41 and SCRC22 combination) regarding cotton leaf premature senescence (μ m)
表4 Table 4 表4(Table 4)
表4 棉花叶片早衰根系主导型(中棉所41与中棉所49组合)Y型嫁接接穗下胚轴各部分厚度及下胚轴半径 Table 4 Thickness of each tissue in cross section of scion hypocotyl and radius of scion hypocotyl in Y type grafts (two scions grafted onto one rootstock, scion+scion/rootstock) of root-dominated pattern (CCRI41 and CCRI49 combination) regarding cotton leaf premature senescence (μ m)
嫁接组合 Scion+scion/rootstock
髓 Pith
木质部 Xylem
韧皮部 Phloem
表皮+皮层 Epidermis+cortex
下胚轴半径 Radius
(C41)+(C41)/C41
306.1 ab
521.3 a
354.5 a
148.1 ab
1330.0 a
(C41)+C49/C41
342.1 a
445.5 ab
288.4 b
206.7 a
1282.7 a
C41+(C49)/C41
237.1 b
372.3 b
242.0 b
146.8 ab
998.2 b
(C41)+C49/C49
272.6 ab
365.4 b
263.0 b
157.7 ab
1058.7 b
C41+(C49)/C49
293.9 ab
420.3 ab
272.3 b
196.2 a
1182.7 ab
(C49)+(C49)/C49
300.0 ab
510.5 a
353.0 a
125.5 b
1289.0 a
C49: CCRI49; C41: CCRI41. The scion in parentheses is the measured. Data within the same column followed by the same letter are not significantly different at P < 0.05 according to Duncan’ s multiple range test. C49: 中棉所49; C41: 中棉所41。括号内接穗为测量接穗。同列数据后的不同字母表示在0.05水平差异显著。
表4 棉花叶片早衰根系主导型(中棉所41与中棉所49组合)Y型嫁接接穗下胚轴各部分厚度及下胚轴半径 Table 4 Thickness of each tissue in cross section of scion hypocotyl and radius of scion hypocotyl in Y type grafts (two scions grafted onto one rootstock, scion+scion/rootstock) of root-dominated pattern (CCRI41 and CCRI49 combination) regarding cotton leaf premature senescence (μ m)
LiB, WangY, Zhang ZY, Wang BM, Eneji AE, Duan LS, LiZH, Tian XL. Cotton shoot plays a major role in mediating senescence induced by potassium deficiency. J Plant Physiol, 2012, 169: 327-335[本文引用:1]
[2]
WangY, LiB, Du MW, Eneji AE, Wang BM, Duan LS, Li ZH, Tian XL. Mechanism of phytohormone involvement in feedback regulation of cotton leaf senescence induced by potassium deficiency. J Exp Bot, 2012, 63: 5887-5901[本文引用:7]
[3]
张巧玉, 王逸茹, 安静, 王保民, 田晓莉. 根-冠互作对棉花叶片衰老的影响. 作物学报, 2017, 43: 226-237Zhang QY, Wang YR, AnJ, Wang BM, Tian XL. Effect of root-shoot interaction on cotton leaf senescence. Acta Agron Sin, 2017, 43: 226-237 (in Chinese with English abstract)[本文引用:8]
[4]
Bauerle TL, CentinariM, Bauerle WL. Shifts in xylem vessel diameter and embolisms in grafted apple trees of differing rootstock growth potential in response to drought. Planta, 2011, 234: 1045-1054[本文引用:1]
[5]
TombesiS, AlmehdiA, DeJong T M. Phenotyping vigour control capacity of new peach rootstocks by xylem vesselanalysis. Sci Hort, 2011, 127: 353-357[本文引用:1]
[6]
Olmstead MA, Lang NS, Ewers FW, Owens SA. Xylem vessel anatomy of sweet cherries grafted onto dwarfing and nondwarfing rootstocks. J Am Soc Hort Sci, 2006, 131: 577-585[本文引用:1]
[7]
SantarosaE, Souza P V D D, Mariath J E D A, Lourosa G V. Physiological Interaction between rootstock-scion: effects on xylem vessels in cabernet sauvignon and merlot grapevines. Am J Enol Viticult, 2016, 67: 65-76[本文引用:1]
[8]
RodríguezgamirJ, Intrigliolo DS, PrimomilloE, Fornerginer MA. Relationships between xylem anatomy, root hydraulic conductivity, leaf/root ratio and transpiration in citrus trees on different rootstocks. Physiol Plant, 2010, 139: 159-169[本文引用:1]
[9]
杨志威. 嫁接对棉花吸收和运输14C-ZR的影响. 中国农业大学硕士学位论文, 北京, 2016Yang ZW. Effect of Grafting on the Uptake and Transportion of 14C-ZR in Cotton. MS Thesis of China Agricultural University, Beijing, China, 2016 (in Chinese with English abstract)[本文引用:2]
[10]
MiyawakiK, Matsumoto-KitanoM, KakimotoT. Expression of cytokinin biosynthetic isopentenyltransferase genes in Arabidopsis: tissue specificity and regulation by auxin, cytokinin, and nitrate. Plant J, 2004, 37: 128-138[本文引用:1]
[11]
KoD, KangJ, KibaT, ParkJ, KojimaM, DoJ, Kim KY, KwonM, EndlerA, Song WY, MartinoiaE, SakakibaraH, LeeY. Arabidopsis ABCG14 is essential for the root-to-shoot translocation of cytokinin. Proc Natl Acad Sci USA, 2014, 111: 7150-7155[本文引用:2]
[12]
Zhang KW, Novak OR, Wei ZY, Gou MY, Zhang XB, YuY, Yang HJ, Cai YH, StrnadM, Liu CJ. Arabidopsis ABCG14 protein controls the acropetal translocation of root-synthesized cytokinins. Nat Commun, 2014, 5: 3274[本文引用:2]
Cheng WH, EndoA, ZhouL, PennyJ, Chen HC, ArroyoA, LeonP, NambaraE, AsamiT, SeoM, KoshibaT, SheenJ. A unique short-chain dehydrogenase/reductase in Arabidopsis glucose signaling and abscisic acid biosynthesis and functions. Plant Cell, 2002, 14: 2723-2743[本文引用:2]
[15]
KoiwaiH, NakaminamiK, SeoM, MitsuhashiW, ToyomasuT, KoshibaT. Tissue-pecific localization of an abscisic acid biosynthetic enzyme, AAO3, in Arabidopsis. Plant Physiol, 2004, 134: 1697-1707[本文引用:3]
[16]
KuromoriT, SugimotoE, ShinozakiK. Intertissue signal transfer of abscisic acid from vascular cells to guard cells. Plant Physiol, 2014, 164: 1587-1592[本文引用:1]
[17]
王逸茹. 根冠互作对棉花下胚轴细胞分裂素和脱落酸相关基因表达的影响. 中国农业大学硕士学位论文, 北京, 2015Wang YR. Effect of Rootstock on The Expression of Cytokinin and Abscisic Acid Related Gene in Cotton Hypocotyl. MS Thesis of China Agricultural University, Beijing, China, 2015 (in Chinese with English abstract)[本文引用:2]
[18]
李博, 王春霞, 张志勇, 段留生, 李召虎, 田晓莉. 适用于低钾条件下棉花苗期根冠通讯研究的三种嫁接方法. 作物学报, 2009, 35: 363-369LiB, Wang CX, Zhang ZY, Duan LS, Li ZH, Tian XL. Three types of grafting techniques available for research of root-shoot communication in cotton (Gossypium hirsutum ) seedlings under low-potassium condition. Acta Agron Sin, 2009, 35: 363-369 (in Chinese with English abstract)[本文引用:1]
[19]
MilhinhosA, Miguel CM. Hormone interactions in xylem development: a matter of signals. Plant Cell Rep, 2013, 32: 867-883[本文引用:1]
[20]
OlesV, PanchenkoA, SmertenkoA. Modeling hormonal control of cambium proliferation. PLoS One, 2017, 12: e0171927[本文引用:1]
[21]
BiemeltS, TschierschH, SonnewaldU. Impact of altered gibberellin metabolism on biomass accumulation, lignin biosynthesis, and photosynthesis in transgenic tobacco plants. Plant Physiol, 2004, 135: 254-265[本文引用:1]
[22]
MauriatM, MoritzT. Analyses of GA20ox- and GID1-over- expressing aspen suggest that gibberellins play two distinct roles in wood formation. Plant J, 2009, 58: 989-1003[本文引用:1]
Guo HY, Wang YC, Liu HZ, HuP, Jia YY, Zhang CR, Wang YM, GuS, Yang CP, Wang C. ExogenousGA(3) application enhances xylem development and induces the expression of secondary wall biosynthesis related genes in betula platyphylla. Internl J Mol Sci, 2015, 16: 22960-22975[本文引用:1]
[25]
Ohashi-ltoK, FukudaH. HD-zip III homeobox genes that include a novel member, ZeHB-13 (Zinnia)/ATHB-15 (Arabidopsis), are involved in procambium and xylem cell differentiation. Plant Cell Physiol, 2003, 44: 1350-1358[本文引用:1]
[26]
Cano-DelgadoA, Yin YH, YuC, VafeadosD, Mora-GarciaS, Cheng JC, Nam KH, Li JM, ChoryJ. BRL1 and BRL3 are novel brassinosteroid receptors that function in vascular differentiation in Arabidopsis. Development, 2004, 131: 5341-5351[本文引用:1]
 , 张睿
, 张睿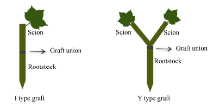




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}