摘要四川盆地小麦高产育种取得了突破性进展, 但高产生理研究相对滞后。本研究以2011—2015连续5年的田间试验, 比较了高产和一般产量潜力品种花后群体冠层结构、主要生理指标和同化物的转运分配特性, 旨在揭示四川小麦9000 kg hm-2高产品种的生理基础。选择代表性高产潜力品种和一般产量潜力品种各3个, 高产潜力品种产量平均9422 kg hm-2, 比一般产量潜力品种高14.3%, 增产原因是生物量或收获指数的提高。相比一般产量潜力品种, 高产潜力品种旗叶短且宽, 长宽比低于10, 开花初期至灌浆中期叶基角和开角增加明显。开花至灌浆后期, 高产潜力品种顶三叶的SPAD值及花后0 d和20 d的群体光合速率显著高于一般产量潜力品种, 群体光合速率以10:00-12:00的差异最大。此外, 高产潜力品种在开花期茎鞘生物量所占比例较高, 而成熟期籽粒所占比例较一般产量潜力品种高1~4个百分点。籽粒产量与小麦形态、生理参数关系密切, 与灌浆期旗叶基角( r = 0.947, P < 0.01)和倒二叶基角( r = 0.963, P < 0.01)呈正相关, 与旗叶长宽比( r = -0.913, P < 0.01)和倒二叶长宽比( r = -0.911, P < 0.01)呈负相关; 与开花期顶三叶SPAD值呈正相关, r值分别为0.75、0.90和0.82 ( P < 0.01); 与成熟期穗轴干重比例呈负相关( r = -0.956, P < 0.01)。本研究表明, 株高适中、株型紧凑, 花后冠层叶绿素含量和群体光合速率较高, 以及合理的物质分配, 是高产潜力品种获得高产的生理基础。
关键词:小麦; 冠层结构; 群体光合速率; 干物质分配; 籽粒产量 Canopy Architecture, Physiological Characteristics and Assimilate Partitioning in Wheat Cultivars with 9000 kg ha-1 Yield Potential in Sichuan Basin WU Xiao-Li1, LI Chao-Su1, TANG Yong-Lu1,*, LI Jun1, MA Xiao-Ling1, LI Shi-Zhao1, HUANG Ming-Bo2 1Crop Research Institute, Sichuan Academy of Agricultural Sciences, Chengdu 610066, China
2 Guanghan Productivity Promotion Center, Guanghan 618300, China
Fund:This study was supported by the China Agriculture Research System (CARS-3-1-23) and the National Natural Science Foundation of China (31571590). AbstractIn contrast to the outstanding breeding progress of high-yield wheat in Sichuan Basin of China, the physiological basis of high-yield-potential cultivars is unclear due to seldom studies. In this study, a five-year field experiment (2011-2015) was carried out to compare the differences of canopy structure after anthesis, canopy apparent photosynthesis (CAP), chlorophyll content (SPAD), and dry matter partitioning between high-yield and normal-yield potential cultivars. Three typical high-yield and three normal-yield potential cultivars were selected, and the average yield of the high-yield potential cultivars (9422 kg ha-1) was 14.3% higher than that of the normal-yield potential cultivars owing to higher biomass or harvest index. Compared with the normal- yield potential cultivars, the high-yield potential cultivars had shorter and wider flag leaves (length-to-width ratio lower than 10) and showed obvious increases of basal and open angles of the topmost three leaves from early anthesis to mid-filling stage. From anthesis to late-filling stage, SPAD values of the topmost three leaves and CAP values at 0 and 20 days after anthesis were significantly higher in the high-yield potential cultivars than in the normal-yield potential cultivars, with the largest difference of CAP between 10:00 and 12:00 a.m. In addition, the high-yield potential cultivars showed higher biomass proportion of stem and sheath at anthesis and higher (1-4 percentage points) biomass proportion of grain at maturity than the normal-yield potential cultivars. Grain yield was closely related to morphological and physiological parameters in wheat. For example, grain yield was positively correlated with basal angles of flag leaf ( r = 0.947, P < 0.01) and the second leaf from top ( r = 0.963, P < 0.01) at grain-filling stage and negatively correlated with leaf length-to-width ratios of flag leaf ( r = -0.913, P < 0.01) and the second leaf from top ( r = -0.911, P < 0.01). Grain yield was also positively correlated ( P < 0.01) with SPAD values of the topmost three leaves ( r = 0.75, 0.90, and 0.82), but negatively correlated with the proportion of spike rachis at maturity ( r = -0.956, P < 0.01). Our results indicate that moderate plant height, compact plant type, high SPAD and CAP values after anthesis, and proper dry matter partitioning are important factors in high-yielding physiology of wheat.
Keyword:Wheat; Canopy architecture; Canopy apparent photosynthesis; Dry matter partitioning; Grain yield Show Figures Show Figures
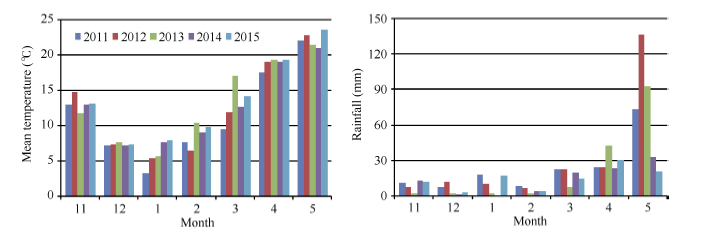
图1 小麦生育期间(11月至翌年5月12日)的月均气温和月降水量Fig. 1 Monthly mean temperature and rainfall amount during wheat growing period (from November to May 12 of the next year)
参试品种6个(表1), 其中典型高产潜力品种3个(川麦42、川麦104和绵麦367, 均多年多点次验收超过9000 kg hm-2), 一般产量潜力品种3个(川麦55、西科麦5号、绵麦37)。3个高产潜力品种均为农业部确定的主推品种。川麦42、绵麦37分别是当前国家区试长江中上游组和四川省区试的对照品种。5年均在10月播种, 具体日期分别为30日、26日、28日、29日和29日; 开花期分别为4月14日至19日、4月3日至7日、3月22日至24日、4月2日至6日和3月26日至30日; 成熟期分别为5月18日至23日、5月10日至15日、5月6日至8日、5月12日至17日和5月6日至9日。随机区组设计, 3次重复, 小区面积3 m × 4 m, 行距20 cm, 基本苗225~266株 m-2, 各品种在同一年份种植密度一致。全年育期施N、P2O5和K2O分别为160、75和75 kg hm-2, 氮肥基施60%, 拔节期追施40%, 磷、钾肥全部作基肥一次性施用。全生育期仅在拔节初期(1月10日左右)灌水一次。喷施杀虫剂和杀菌剂控制病虫害。 表1 Table 1 表1(Table 1)
表1 参试品种信息 Table 1 Information of wheat cultivars
品种 Cultivar
系谱 Pedigree
矮秆基因 Dwarfing gene
审定年份 Year released
区试产量 YD (kg hm-2)
增产率 Increased (%)
验收最高产量 MD (kg hm-2)
川麦42 Chuanmai 42
Syn769/SW3243//川6415
Rht1
2004
6130
16.4
10660
川麦104 Chuanmai 104
川麦42/川农16
Rht1
2012
6120
14.1
10440
绵麦367 Mianmai 367
1275-1/川麦43
Rht1
2010
5690
14.0
9750
川麦55 Chuanmai 55
SW3243/SW8688
Rht2
2009
5420
15.0
8250
西科麦5号 Xikemai 5
贵农21/96II-39
Rht2
2008
5510
10.1
—
绵麦37 Mianmai 37
96EW37/绵阳90-100
Rht2, Rht8
2004
5110
12.6
—
YD stands for yield in the regional trials at national or Sichuan provincial level, and the increased ratio was compared to the control cultivar Chuanmai 42 or Mianmai 37. MD stands for the maximum yield from farmers’ fields. 增产率为在国家或四川省区试中较对照(川麦42或绵麦37)产量的增加百分比。
表3 参试品种的株高、节间构成指数和倒伏情况 Table 3 Differences in plant height, internode index, and lodging amongst genotypes across years
品种 Cultivar
株高 Plant height (cm)
节间构成指数 Internode index
倒伏百分率 Lodging percentage (%)
L1
L2
L3
2011
2012
2013
2014
2015
川麦42 Chuanmai 42
98.2± 3.9
0.63
0.59
0.54
25 /++
75 /++
5 /+
70 /++
0
川麦104 Chuanmai 104
94.6± 1.9
0.61
0.59
0.54
0
20 /+
0
10 /+
0
绵麦367 Mianmai 367
89.6± 2.7
0.64
0.58
0.54
0
45 /++
5 /+
50 /++
0
川麦55 Chuanmai 55
87.6± 3.4
0.65
0.59
0.54
0
0
0
5 /+
0
西科麦5号 Xikemai 5
85.2± 3.6
0.64
0.59
0.55
30 /+
0
0
25 /+
0
绵麦37 Mianmai 37
84.7± 3.6
0.64
0.58
0.54
0
75 /++
20 /+
50 /++
0
平均值 Mean
90.0
0.63
0.59
0.54
LSD0.05
1.300
0.012
0.014
0.015
Plant height was the mean± SD of five years and internode index was the average across 2013-2015. Lodging severity was classified according to the angle between vertical direction and the stem, in which + and ++ represent light (< 40° ) and serious (40-90° ) lodging, respectively. 株高是5年试验数据, 节间构成指数是2013-2015年数据。+为轻度倒伏, 指茎与垂直方向的夹角小于40° ; ++为重度倒伏, 指茎与垂直方向的夹角在40~90° 之间。
表3 参试品种的株高、节间构成指数和倒伏情况 Table 3 Differences in plant height, internode index, and lodging amongst genotypes across years
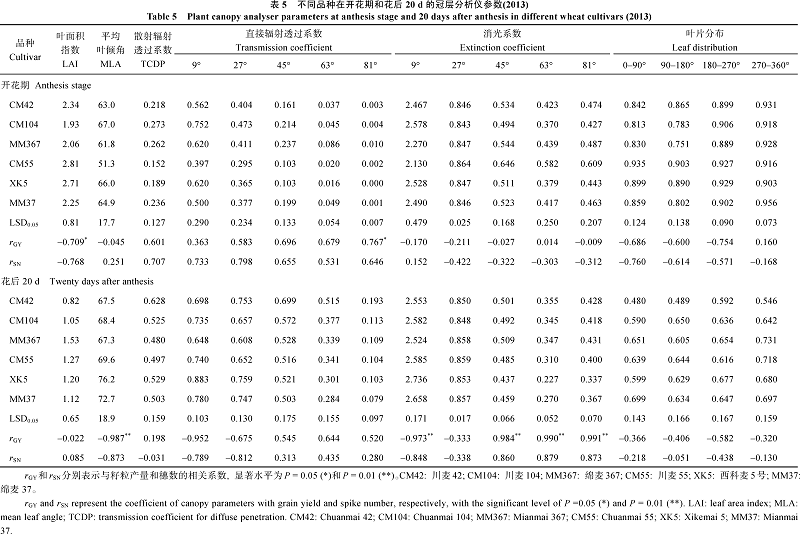
表4 开花期和灌浆中期上三叶的叶长、叶宽、开角、基角 Table 4 Leaf length, width, open, and basal angles for the top three leaves at anthesis and mid-filling stage
品种 Cultivar
开花期 Anthesis stage
灌浆中期 Mid-filling stage
叶长 Leaf length (cm)
叶宽 Leaf width (cm)
长宽比 Length/width ratio
开角 Open angle (° )
基角 Basal angle (° )
开角 Open angle (° )
基角 Basal angle (° )
旗叶 Flag leaf
川麦42 Chuanmai 42
20.4
2.3
8.8
51.7
38.1
120.2
110.8
川麦104 Chuanmai 104
18.3
2.1
8.0
62.5
20.0
129.8
104.2
绵麦367 Mianmai 367
21.1
2.2
9.7
57.9
26.4
101.1
90.4
川麦55 Chuanmai 55
23.4
2.1
11.0
39.4
18.3
31.1
17.0
西科麦5号Xikemai 5
24.7
2.3
10.7
61.2
34.2
111.7
52.5
绵麦37 Mianmai 37
22.6
2.1
10.9
64.2
20.1
42.9
32.6
LSD0.05
1.8
0.1
1.0
30.3
9.4
22.6
36.2
rGY
-0.78*
0.52
-0.82* *
-0.23
0.05
0.60
0.73*
rSN
-0.79*
0.78*
-0.91* *
0.22
0.57
0.86* *
0.95* *
倒二叶 The second leaf from the top
川麦42 Chuanmai 42
26.8
2.1
12.9
81.4
34.4
132.8
99.7
川麦104 Chuanmai 104
28.3
1.9
15.1
110.1
34.7
134.9
100.3
绵麦367 Mianmai 367
29.4
2.0
15.0
91.7
31.9
132.3
77.8
川麦55 Chuanmai 55
33.9
1.9
17.6
64.4
18.9
95.2
43.0
西科麦5号Xikemai 5
30.8
2.0
15.1
75.8
29.8
121.4
60.8
绵麦37 Mianmai 37
31.1
1.9
16.2
85.6
26.1
101.4
41.4
LSD0.05
3.0
0.1
1.9
22.2
8.3
32.8
34.0
rGY
-0.53
-0.15
-0.39
0.52
0.52
0.68*
0.80* *
rSN
-0.96* *
0.32
-0.91* *
0.61
0.90* *
0.88* *
0.96* *
倒三叶 The third leaf from the top
川麦42 Chuanmai 42
26.0
1.7
15.1
105.0
55.3
141.9
73.9
川麦104 Chuanmai 104
24.2
1.7
14.6
99.3
51.7
139.4
101.9
绵麦367 Mianmai 367
25.8
1.7
15.2
106.6
61.3
158.1
121.8
川麦55 Chuanmai 55
31.5
1.7
18.7
61.4
28.2
117.9
44.0
西科麦5号Xikemai 5
26.3
1.8
14.9
96.3
59.9
139.7
134.7
绵麦37 Mianmai 37
28.6
1.7
16.6
90.3
53.9
149.2
113.9
LSD0.05
2.5
0.1
1.7
35.2
36.4
42.8
38.9
rGY
-0.51
-0.67*
-0.37
0.30
-0.03
-0.02
-0.31
rSN
-0.81* *
-0.14
-0.77*
0.75*
0.48
0.35
0.01
Data are the means over 2014 and 2015. rGY and rSN represent the coefficient of leaf parameters with grain yield and spike number, respectively, with the significant level of P =0.05 (* ) and P = 0.01 (* * ). 数据为2014年和2015年的平均值。rGY和rSN分别表示与籽粒产量和穗数的相关系数, 显著水平为P= 0.05 (* )和P = 0.01 (* * )。
表4 开花期和灌浆中期上三叶的叶长、叶宽、开角、基角 Table 4 Leaf length, width, open, and basal angles for the top three leaves at anthesis and mid-filling stage
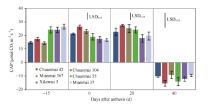
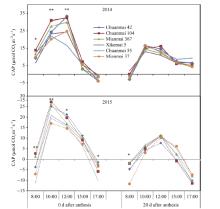
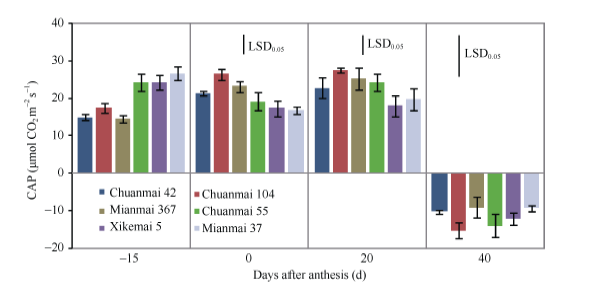
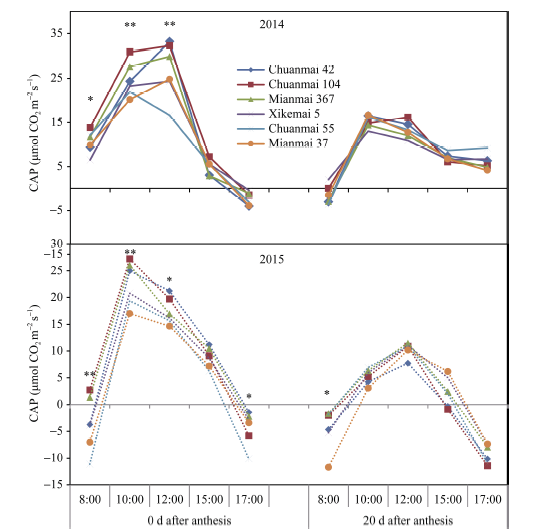
图3 花后0 d和20 d各品种群体光合速率(CAP)的日变化Fig. 3 Daily changes in canopy apparent photosynthesis (CAP) in different cultivars at 0 and 20 days after anthesis * P< 0.05, * * P< 0.01.
Fischer RA. Wheat physiology: a review of recent developments. Crop Pasture Sci, 2011, 62: 95-114[本文引用:4]
[2]
张法全, 王小燕, 于振文, 王西芝, 白洪立. 公顷产10000 kg小麦氮素和干物质积累与分配特性. 作物学报, 2009, 35: 1086-1096Zhang FQ, Wang XY, Yu ZW, Wang XZ, Bai HL. Characteristics of accumulation and distribution of nitrogen and dry matter in wheat at yield level of ten thousand kilograms per hectare. Acta Agron Sin, 2009, 35: 1086-1096 (in Chinese with English abstract)[本文引用:3]
[3]
Jaradat AA. Modeling biomass allocation and grain yield in bread and durum wheat under abiotic stress. Aust J Crop Sci, 2009, 3: 237-248[本文引用:2]
[4]
MadaniA, Rad AS, PazokiA, NourmohammadiG, ZarghamiR. Wheat (Triticum aestrivum) grain filling and dry matter partitioning responses to source: sink modifications under post anthesis water and nitrogen deficiency . Acta Sci Agron, 2010, 32: 145-151[本文引用:1]
[5]
Tian ZW, JingQ, Dai TB, JiangD, Cao WX. Effects of genetic improvements on grain yield and agronomic traits of winter wheat in the Yangtze River Basin of China. Field Crops Res, 2011, 124: 417-425[本文引用:3]
[6]
Xiao YG, QianZG, WuK, Liu JJ, Xia XC, Ji WQ, He ZH. Genetic gains in grain yield and physiological traits of winter wheat in Shand ong province, China, from 1969 to 2006. Crop Sci, 2012, 52: 44-56[本文引用:4]
[7]
Miralles DJ, Slafer GA. Radiation interception and radiation use efficiency of near-isogenic wheat lines with different height. Euphytica, 1997, 97: 201-208[本文引用:1]
[8]
雷振生, 林作揖. 黄淮麦区冬小麦合理株型结构研究. 华北农学报, 1994, 9(4): 27-32Lei ZS, Lin ZJ. Studies on ideotype characters of winter wheat in Huang-Huai wheat region. Acta Agric Boreal-Sin, 1994, 9(4): 27-32 (in Chinese with English abstract)[本文引用:3]
[9]
Hansen KA, Martin JM, Lanning SP, Talbert LE. Correlation of genotype for agronomic and physiological traits in space-planted versus densely seeded conditions. Crop Sci, 2005, 45: 1023-1028[本文引用:1]
[10]
Blake NK, Lanning SP, Martin JM, Sherman JD, Talbert LE. Relationship of flag characteristics to economically important traits in two spring wheat crosses. Crop Sci, 2007, 47: 491-496[本文引用:3]
Zhao CJ, Wang JH, Huang WJ, Zhou QF. Spectral indices sensitively discriminating wheat genotypes of different canopy architectures. Precis Agric, 2010, 11, 557-567[本文引用:3]
[13]
RajaramS, van GinkelM. Yield potential debate: germplasm vs. methodology, or both. In: Reynolds M P, Rajaram S, McNab A, eds. Increasing Yield Potential in Wheat: Breaking the Barriers. Mexico D. F. : CIMMYT, 1996. pp 101-133[本文引用:4]
[14]
Liu YK, Li MJ, Li JY, Li XJ, Yang XH, Tong YP, Zhang AM, LiB, Lin JX, Kuang TY, Li Z S. Dynamic changes in flag leaf angle contribute to high photosynthetic capacity. Chin Sci Bull, 2009, 54: 3045-3052[本文引用:3]
[15]
Long SP, Zhu XG, Naidu SL, Ort DR. Can improvement in photosynthesis increase crop yields? Plant Cell Environ, 2006, 29: 315-330[本文引用:4]
[16]
魏燮中. 小麦株型结构分析法与消光系数分布的基因型间差异. 南京农业大学学报, 1987, (3): 21-28Wei XZ. Plant type architecture analysis (PTAA) and the differences of K vertical distribution in canopy among wheat genotypes. J Nanjing Agric Univ, 1987, (3): 21-28 (in Chinese with English abstract)[本文引用:1]
[17]
Leihner DE, Guillermo OF. Improvement of durum wheat-plant type, yield potential, and adaptation. Euphytica, 1978, 27: 785-799[本文引用:3]
[18]
Fischer RA. Understand ing the physiological basis of yield potential in wheat. J Agric Sci, 2007, 2007, 145: 99-113[本文引用:2]
[19]
Sharma-NatuP, Ghildiyal MC. Potential targets for improving photosynthesis and crop yield. Curr Sci India, 2005, 88: 1918-1928[本文引用:3]
[20]
Evans LT, Dunstone RL. Some physiological aspects of evolution in wheat. Aust J Biol Sci, 1970, 23: 725-741[本文引用:2]
[21]
Reynolds MP, Delgado B M I, Gutiérrez-Rodríguez, Larqué- Saavedra A. Photosynthesis of wheat in warm, irrigated environment: I. Genetic diversity and crop productivity. Field Crops Res, 2000, 66: 37-50[本文引用:4]
[22]
董树亭. 高产冬小麦群体光合能力与产量关系的研究. 作物学报, 1991, 17: 461-469Dong ST. Studies on the relationship between canopy apparent photosynthesis and grain yield in high-yielding winter wheat. Acta Agron Sin, 1991, 17: 461-469 (in Chinese with English abstract)[本文引用:3]
[23]
Tang YL, Rosewarne GM, Li CS, Wu XL, Yang WY, WuC. Physiological factors underpinning grain yield improvements of synthetic-derived wheat in southwestern China. Crop Sci, 2015, 55: 98-112[本文引用:4]
[24]
HafsiM, HadjiA, GuendouzA, MaamariK. Relationship between flag leaf senescence and grain yield in durum wheat growth under drought conditions. J Agron, 2013, 12: 69-77[本文引用:1]
[25]
KumariM, Pudake RN, Singh VP, Joshi AK. Association of staygreen trait with canopy temperature depression and yield traits under terminal heat stress in wheat ( Triticum aestrivum L. ). Euphytica, 2013, 190: 87-97[本文引用:1]
[26]
Foulkes MJ, Slafer GA, Davies WJ, Berry PM, Sylvester-BradleyR, MartreP, Callerini DF, GriffithsS, Reynolds MP. Raising yield potential of wheat: III. Optimizing partitioning to grain while maintaining lodging resistance. J Exp Bot, 2011, 62: 469-486[本文引用:4]
GiuntaF, MotzoR, PrunedduG. Has long-term selection for yield in durum wheat also induced changes in leaf and canopy traits? Field Crops Res, 2008, 106: 68-76[本文引用:1]
[29]
Acreche MM, Briceńo-FélixG, Sánchez J A M, Slafer G A. Physiological bases of genetic gains in Mediterranean bread wheat yield in Spain. Eur J Agron, 2008, 28: 162-170[本文引用:2]
[30]
Austin RB, BinghamJ, Blackwell RD, Evans LT, Ford MA, Morgan CL, TaylorM. Genetic improvement in winter wheat yield since 1900 and associated physiological changes. J Agric Sci(Camb), 1980, 94: 675-689[本文引用:3]
[31]
ZhouY, Zhu HZ, Cai SB, He ZH, Zhang XK, Xia XC, Zhang GS. Genetic improvement of grain yield and associated traits in the southern China winter wheat region: 1949-2000. Euphitica, 2007, 157: 465-473[本文引用:1]
[32]
LiJ, Wan HS, Yang WY. Synthetic hexaploid wheat enhances variation and adaptive evolution of bread wheat in breeding processes. J Syst Evol, 2014, 52: 735-742[本文引用:1]
[33]
魏燮中, 吴兆苏. 小麦植株高度的结构分析. 南京农学院学报, 1983, (1): 14-21Wei XZ, Wu ZS. Analysis of plant height and structure. J Nanjing Agric Coll, 1983, (1): 14-21 (in Chinese with English abstract)[本文引用:2]
[34]
Parry M AJ, ReynoldsM, Salvucci ME, RainesC, Andralojc PJ, Zhu XG, Price GD, Condon AG, Furbank RT. Raising yield potential of wheat: II. Increasing photosynthetic capacity and efficiency. J Exp Bot, 2011, 62: 453-467[本文引用:1]
[35]
Aliyev JA. Photosynthesis, photorespiration and productivity of wheat and soybean genotypes. Physiol Plant, 2012, 145: 369-383[本文引用:1]
[36]
胡延吉, 兰进好, 赵坦方, 高法振. 不同穗型的两个冬小麦品种冠层结构及光合特性的研究. 作物学报, 2000, 26: 905-912Hu YJ, Lan JH, Zhao TF, Gao FZ. Canopy architecture and photosynthetic characteristics in two winter wheat cultivars with different spike type. Acta Agron Sin, 2000, 26: 905-912 (in Chinese with English abstract)[本文引用:1]
[37]
李朴芳, 程正国, 赵鸿, 张小丰, 李冀南, 王绍明, 熊友才. 旱地小麦理想株型研究进展. 生态学报, 2011, 31: 2631-2640Li PF, Cheng ZG, ZhaoH, Zhang XF, Li JN, Wang SM, Xiong YC. Current progress in plant ideotype research of dry land wheat ( Triticum aestivum L. ). Acta Ecol Sin, 2011, 31: 2631-2640 (in Chinese with English abstract)[本文引用:2]
[38]
YildirimM, KoçM, AkinciC, BarutçularC. Variations in morphological and physiological traits of bread wheat diallel crosses under timely and late sowing conditions. Field Crops Res, 2013, 140: 9-17[本文引用:1]
[39]
ReesD, Ibarra LR, AcevedoE, Mujeeb-KaziA, Villareal RL. Photosynthetic Characteristics of Synthetic Bread Wheats. Wheat Special Report No. 28. Mexico, D. F. : CIMMYT, 1994[本文引用:1]
[40]
Reynolds MP, PellegrineschiA, Skovmand B. Sink-limitation to yield and biomass: a summary of some investigations in spring wheat. Ann Appl Biol, 2005, 146: 39-49[本文引用:2]
Underdahl JL, MergoumM, Ransom JK, Schatz BG. Agronomic traits improvement and associations in hard red spring wheat cultivars released in North Dakota from 1968 to 2006. Crop Sci, 2008, 48: 158-166[本文引用:1]
[43]
Tang YL, Li CS, Yang WY, Wu YQ, Wu XL, WuC, Ma XL, Li SZ, Rosewarne GM. Quality potential of synthetic-derived commercial wheat cultivars in southwestern China. Crop Pasture Sci, 2016, 67: 583-593[本文引用:1]
 , 李俊
, 李俊

{kind=link}
{kind=link}
{kind=link}