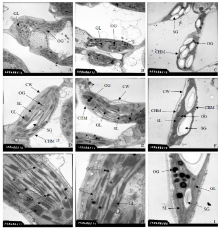
关键词:大豆; 套作; 光合荧光特性; 叶绿体超微结构 Effects of Shading on Leaf Structure and Photosynthetic Fluorescence Characteristics of Soybean Seedlings in Maize-Soybean Relay Intercropping System FAN Yuan-Fang, YANG Feng*, LIU Qin-Lin, CHEN Jun-Xu, WANG Rui, LUO Shi-Ling, YANG Wen-Yu* College of Agronomy, Sichuan Agricultural University / Key Laboratory of Crop Ecophysiology and Farming System in Southwest, Ministry of Agriculture, Chengdu 611130, China. Fund:The study was supported by the National Natural Science Foundation of China (31571615), the National Key Research and development Program of China (2016YFD0300209), and the Innovation Program of Sichuan Agricultural University ( 04060658) AbstractShading in relay intercropping system is a major factor affecting the low growing of crop growth and development. This study attempted to compare leaf structure and photosynthetic characteristics of cultivar Nandou 12 under maize-soybean relay intercropping systems. Three treatments were designed including one row maize to one row soybean with 50 cm of row space between maize and soybean (A1); two rows maize to two rows soybean with 40 cm of row space for maize or soybean and 60 cm of row space between maize and soybean (A2), sole cropping of soybean. The result indicated that shading caused the changes of both anatomical structure of leaf and photosynthetic characteristics in maize/soybean relay strip intercropping systems. During the V5 stage of soybean, the PAR density in treatments (A1 and A2) were lower than that in CK by 91.2% and 66.8%, respectively. The content of Chl a of treatments (A1 and A2) was lower than that in CK by 50.2% and 27.9%. The Pn of treatments (A1 and A2) was lower than that in CK by 63.2% and 37.8%. The Fv/ Fm, qP, NPQ in leaf of treatments (A1 and A2) were lower than there in CK significantly, Fv'/ Fm' and Fq'/ Fm' of treatments (A1 and A2) were higher than that in CK. Under light microscope, compared with CK, A1 and A2 treatments decreased palisade tissue thickness and spongy tissue thickness of leaf. The leaf of treatment CK was 2.15 and 1.69 times as thick as that of treatments A1 and A2. In A1 and A2 treatments leaf chloroplast morphology was normal, the number of chloroplasts, granule lamella and osmophilic globule in chloroplast increased; the structure of chloroplast was not damaged; There were few starch grains in chloroplasts; and the structure of cell organelles was normal. Therefore, the different spatial patterns of maize-soybean intercropping system result in different light environments above the soybean canopy, and directly affect soybean leaf structure and photosynthetic fluorescence characteristics.
Keyword:Soybean; Intercropping; Photosynthetic fluorescence characteristics; Chloroplast ultrastructure Show Figures Show Figures
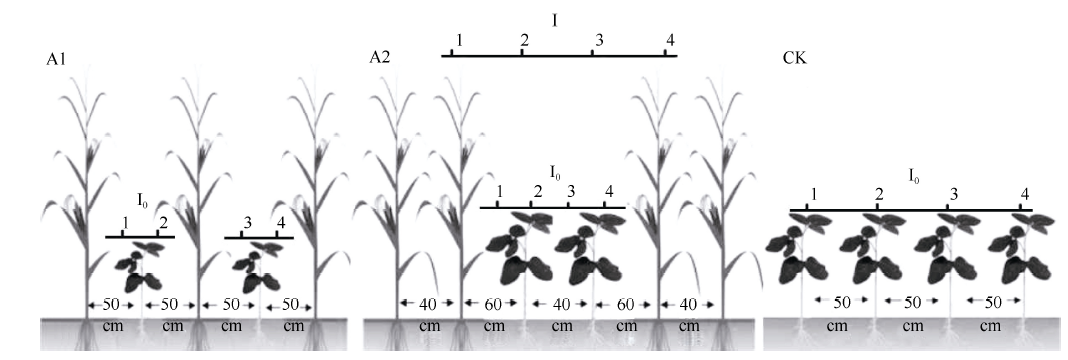
图1 PAR传感器在玉米-大豆带状套作组合中的摆放位置示意图Fig. 1 Diagram of maize-soybean relay strip intercropping patterns and layout of photo synthetically active radiation sensors
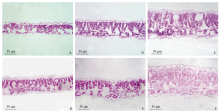
图2 玉米-大豆套作不同行比配置下大豆叶片解剖结构 A~C: A1、A2处理和CK (2014年); D~F: A1, A2处理和CK (2015年) (× 400)。Fig. 2 Anatomical structure of soybean leaf in different maize-soybean relay intercropping systems A-C: A1, A2 treatments and CK (2014); D-F: A1, A2 treatments and CK (2015) (× 400).
表5 Table 5 表5(Table 5)
表5 玉米-大豆套作不同行比配置下大豆叶片解剖结构 Table 5 Anatomical structure parameter of soybean leaf in different maize-soybean relay intercropping systems
处理 Treatment
上表皮厚度 Upper epidermis thickness (μ m)
下表皮厚度 Lower epidermis thickness (μ m)
栅栏组织厚度 Palisade tissue thickness (μ m)
海绵组织厚度 Spongy tissue thickness (μ m)
叶片厚度 Leaf thickness (μ m)
栅海比 Palisade tissue/ spongy tissue
2014
A1
9.86± 1.13 a
6.73± 0.41 c
21.46± 1.84 c
24.82± 1.92 b
66.44± 3.46 c
0.87± 0.11 b
A2
9.58± 1.88 a
8.35± 1.11 b
36.20± 2.65 b
23.18± 4.98 b
82.43± 8.45 b
1.60± 0.24 a
CK
11.08± 1.30 a
11.67± 1.71 a
72.47± 2.35 a
45.06± 6.39 a
148.02± 6.84 a
1.64± 0.23 a
2015
A1
10.50± 1.22 b
8.93± 0.31 b
26.36± 1.79 c
26.01± 1.32 b
73.26± 4.85 c
1.02± 0.10 b
A2
12.61± 1.00 a
11.78± 0.69 a
43.36± 1.61 b
26.72± 1.87 b
95.85± 2.26 b
1.63± 0.16 a
CK
13.39± 0.90 a
12.54± 1.00 a
80.04± 2.88 a
50.29± 2.87 a
151.02± 3.81 a
1.60± 0.11 a
Values followed by different lowercases within the same column are significantly different at the 0.05 probability levels. 同一列中标以不同小写字母的值差异达0.05显著水平。
表5 玉米-大豆套作不同行比配置下大豆叶片解剖结构 Table 5 Anatomical structure parameter of soybean leaf in different maize-soybean relay intercropping systems
高小丽, 高金锋, 冯佰利, 王鹏科, 柴岩. 不同绿豆品种的叶片解剖结构. 作物学报, 2012, 38: 181-185Gao XL, Gao JF, Feng BL, Wang PK, ChaiY. Anatomical structure of leaf in different mung bean varieties. Acta Agron Sin, 2012, 38: 181-185 (in Chinese with English abstract)[本文引用:1]
[2]
冯乃杰, 郑殿峰, 赵玖香, 祖伟, 杜吉到, 张玉先, 梁喜龙. 植物生长物质对大豆叶片形态解剖结构及光合特性的影响. 作物学报, 2009, 35: 1691-1697Feng NJ, Zheng DF, Zhao JX, ZuW, Du JD, Zhang YX, Liang XL. Effect of plant growth substances on morphological and anatomical structure of leaf and photosynthetic characteristics in soybean. Acta Agron Sin, 2009, 35: 1691-1697[本文引用:2]
[3]
覃凤飞, 李强, 崔棹茗, 李洪萍, 杨智然. 越冬期遮阴条件下3个不同秋眠型紫花苜蓿品种叶片解剖结构与其光生态适应性. 植物生态学报. 2012, 36: 333-345Qin FF, LiQ, Cui ZM, Li HP, Yang ZR. Leaf anatomical structures and ecological adaptabilities to light of three alfalfa cultivars with different fall dormancies under shading during overwintering. Chin J Plant Ecol, 2012, 36: 333-345 (in Chinese with English abstract)[本文引用:2]
[4]
甄伟, 张福墁. 弱光对黄瓜功能叶片光合特性及超微结构的影响. 园艺学报, 2000, 27: 290-292ZhenW, Zhang FM. The effects of low light intensity on photosynthetic characteristics and ultrastructure of cucumber functional leaves. Acta Hort Sin, 2000, 27: 290-292[本文引用:2]
[5]
马慧丽, 吕德国. 光照条件对“寒富”苹果叶片结构和光合特性的影响. 应用生态学报, 2014, 25: 1927-1932Ma HL, Lyu DG. Effects of light condition on structure and photosynthetic characteristics of leaves in “Hanfu” apple. Chin J Appl Ecol, 2014, 25: 1927-1932 (in Chinese with English abstract)[本文引用:2]
[6]
黄俊, 郭世荣, 吴震, 李式军. 弱光对不结球白菜光合特性与叶绿体超微结构的影响. 应用生态学报, 2007, 18: 352-358HuangJ, Guo SR, WuZ, Li SJ. Effects of weak light on photosynthetic characteristics and chloroplast ultrastructure of nonheading Chinese cabbage. Chin J Appl Ecol, 2007, 18: 352-358 (in Chinese with English abstract)[本文引用:2]
[7]
李芳兰, 包维楷. 植物叶片形态解剖结构对环境变化的响应与适应. 植物学通报, 2005, (增刊1): 118-127Li FL, Bao WK. Responses of the morphological and anatomical structure of the plant leaf to environmental change. Chin Bull Bot, 2005, (suupl-1): 118-127 (in Chinese with English abstract)[本文引用:1]
[8]
姚允聪, 王绍辉, 孔云. 弱光条件下桃叶片结构及光合特性与叶绿体超微结构变化. 中国农业科学, 2007, 40: 855-863Yao YC, Wang SH, KongY. Characteristics of photosynthesis mechanism in different peach species under low light intensity. Sci Agric Sin, 2007: 40: 855-863 (in Chinese with English abstract)[本文引用:3]
[9]
吴涛, 耿云芬, 柴勇, 郝佳波, 袁春明. 三叶爬山虎叶片解剖结构和光合生理特性对3种生境的响应. 生态环境学报, 2014, 23: 1586-1592WuT, Geng YF, ChaiY, Hao JB, Yuan CM. Response of leaf anatomical structure and photosynthesis characteristics of Parthenocissus himalayana to three habitat types. Ecol Environ Sci, 2014, 23: 1586-1592 (in Chinese with English abstract)[本文引用:2]
[10]
FanX, XuZ, LiuX, Liu XY, Tang CM, Wang LW, Han XL. Effects of light intensity on the growth and leaf development of young tomato plants grown under a combination of red and blue light. Sci Hort (Amsterdam), 2013, 153: 50-55[本文引用:2]
[11]
Jiang CD, WangX, Gao HY, ShiL, Chow WS. Systemic regulation of leaf anatomical structure, photosynthetic performance, and high-light tolerance in sorghum. Plant Physiol, 2011, 155: 1416-1424[本文引用:1]
[12]
赵团结, 盖钧镒, 李海旺, 邢邯, 邱家驯. 超高产大豆育种研究的进展与讨论. 中国农业科学, 2006, 39: 29-37Zhao TJ, Gai JY, Li HW, XingH, Qiu JX. Advances in breeding for super high-yielding soybean cultivars. Sci Agric Sin, 2006, 39: 29-37 (in Chinese with English abstract)[本文引用:1]
[13]
Fehr WR, Caviness CE. Stages of Soybean Development. Special Report 80, Cooperative Extension Service, Agriculture and Home Economic Experiment Station. Ames, Iowa: Iowa State University, 1977. pp 1-11[本文引用:1]
[14]
崔亮, 苏本营, 杨峰, 杨文钰. 不同玉米-大豆带状套作组合条件下光合有效辐射强度分布特征对大豆光合特性和产量的影响. 中国农业科学, 2013, 47: 1489-1501CuiL, Su BY, YangF, Yang WY. Effects of photo-synthetically active radiation on photosynthetic characteristics and yield of soybean in different maize/soybean relay strip intercropping systems. Sci Agric Sin, 2013, 47: 1489-1501 (in Chinese with English abstract)[本文引用:3]
[15]
李艳大, 汤亮, 张玉屏, 刘蕾蕾, 曹卫星, 朱艳. 水稻冠层光合有效辐射的时空分布特征. 应用生态学报, 2010, 21: 952-958Li YD, TangL, Zhang YP, Liu LL, Cao WX, ZhuY. Spatiotemporal distribution of photosynthetically active radiation in rice canopy. Chin J Appl Ecol, 2010, 21: 952-958 (in Chinese with English abstract)[本文引用:1]
[16]
吕丽华, 赵明, 赵久然, 陶洪斌, 王璞. 不同施氮量下夏玉米冠层结构及光合特性的变化. 中国农业科学, 2008, 41: 2624-2632Lyu LH, ZhaoM, Zhao JR, Tao HB, WangP. Canopy structure and photosynthesis of summer maize under different nitrogen fertilizer application rates. Sci Agric Sin, 2008, 41: 2624-2632 (in Chinese with English abstract)[本文引用:1]
[17]
王锐, 杨峰, 张勇, 黄山, 雍太文, 刘卫国, 杨文钰. 套作大豆后期叶片叶绿素荧光特性及光谱特征分析. 核农学报, 2015, 29: 1182-1189WangR, YangF, ZhangY, HuangS, Yong TW, Liu WG, Yang WY. The analysis of chlorophyll fluorescence parameters and hyperspectral characteristics of soybean after maize harvest under relay intercropping systems. Acta Agric Nucl Sin, 2015, 29: 1182-1189 (in Chinese with English abstract)[本文引用:1]
[18]
Arnon DI. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol, 1949, 24: 1-15[本文引用:1]
[19]
杨虎彪, 李晓霞, 罗丽娟. 植物石蜡制片中透明和脱蜡技术的改良. 植物学报, 2009, 44: 230-235Yang HB, Li XX, Luo LJ. An improved clearing and de-waxing method for plant paraffin sectioning. Chin Bull Bot, 2009, 44: 230-235 (in Chinese with English abstract)[本文引用:1]
[20]
苏本营, 宋艳霞, 陈圣宾, 杨文钰. 大豆幼苗对套作玉米遮阴环境的光合生理生态响. 生态学报, 2015, 35: 3298-3308Su BY, Song YX, Chen SB, Yang WY. Photosynthetic responses of soybean (Glycine max) seedlings to shading caused by maize in an intercropping system. Acta Ecol Sin, 2015, 35: 3298-3308 (in Chinese with English abstract)[本文引用:2]
[21]
李瑞, 文涛, 唐艳萍, 孙歆, 夏超. 遮阴对大豆幼苗光合和荧光特性的影响. 草业学报, 2014: 198-206LiR, WenT, Tang YP, SunX, XiaC. Effect of shading on photosynthetic and chlorophyll fluorescence characteristics of soybean. Acta Prat Sin, 2014: 198-206[本文引用:1]
[22]
Demmig-AdamsB, Adams WW. The role of xanthophyll cycle carotenoids in the protection of photosynthesis. Trends Plant Sci, 1996, 1: 21-26[本文引用:1]
[23]
Hu WH, Yu JQ. Effects of chilling under low light on photosynthesis and chlorophyll fluorescence characteristic in tomato leaves. Acta Hort Sin, 2001, 28: 41-46[本文引用:1]
[24]
Zhang GR, Du WG, Man WQ, Li GQ, Gui MZ, Wang XD, Ge QY, Hao NB. Study on leaf comparative anatomy of different genotypes of soybean. Chin Bull Bot, 2002, 19: 208-214[本文引用:1]
[25]
GroverA, MohantyP. Leaf senescence-induced alterations in structure and function of higher plant chloroplasts. In: Abrol Y P, Mohanty P, Govindjee, eds. Photosynthesis: Photoreactions to Plant Productivity. Dordrecht: Kluwer Academic Publishers, 1992. pp 225-255[本文引用:1]
[26]
HuangD, WuL, Chen JR, DongL. Morphological plasticity, photosynthesis and chlorophyll fluorescence of Athyrium pachyphlebium at different shade levels. Photosynthetica, 2011, 49: 611-618[本文引用:2]
[27]
吴正锋, 孙学武, 王才斌, 郑亚萍, 万书波, 刘俊华, 郑永美, 吴菊香, 冯昊, 于天一. 弱光胁迫对花生功能叶片RuBP羧化酶活性及叶绿体超微结构的影响. 植物生态学报, 2014, 38: 740-748Wu ZF, Sun XW, Wang CB, Zheng YP, Wan SB, Liu JH, Zheng YM, Wu JX, FengH, Yu TY. Effects of low light stress on rubisco activity and the ultrastructure of chloroplast in functional leaves of peanut. Acta Ecol Sin, 2014, 38: 740-748 (in Chinese with English abstract)[本文引用:2]
[28]
艾希珍, 郭延奎, 马兴庄, 邢禹贤. 弱光条件下日光温室黄瓜需光特性及叶绿体超微结构. 中国农业科学, 2004, 37: 268-273Ai XZ, Guo YK, Ma XZ, Xing YX. Photosynthetic characteristics and ultrastructure of chloroplast of cucumber under low light intensity in solar greenhouse. Sci Agric Sin, 2004, 37: 268-273 (in Chinese with English abstract)[本文引用:3]
[29]
刘凡值, 苟兴红, 傅生华, 梁慕勤. 大豆耐阴性的研究: V. 大豆叶片形态特征及解剖结构与耐阴性的关系. 贵州农业科学, 1990, (3): 9-16Liu FZ, Gou XH, Fu SH, Liang MQ. Studies on shading-endurance of soybean [Glycing max (L. ) Merr. ]: V. A relation between the morphogical and anatomical characters of soybean leaf and the shade-endurance. Guizhou Agric Sci, 1990, (3): 9-16 (in Chinese with English abstract)[本文引用:1]
[30]
徐克章, 张治安, 王英典. 光强对人参叶片显微和超微结构的影响. 植物学报, 1994, 11(增刊): 23-27Xu KZ, Zhang ZA, Wang YD. Effect of light density on microstructure and ultrastructure of ginseng leaves. Chin Bull Bot, 1994, 11(suppl): 23-27 (in Chinese with English abstract)[本文引用:2]
[31]
张振贤, 郭延奎, 邹琦. 遮阴对生姜叶片显微结构及叶绿体超微结构的影响. 园艺学报, 1999, 26: 96-100Zhang ZX, Guo YK, ZouQ. Effect of shading on ultrastructure of chloroplast and microstructure of ginger leaves. Acta Hort Sin, 1999, 26: 96-100 (in Chinese with English abstract)[本文引用:2]
[32]
魏珉, 邢禹贤, 王秀峰, 马红. CO2加富对黄瓜叶片显微和亚显微结构的影响. 园艺学报, 2002, 29: 30-34WeiM, Xing YX, Wang XF, MaH. Effect of CO2 enrichment on the microstructure and ultrastructure of leaves in cucumber. Acta Hort Sin, 2002, 29: 30-34 (in Chinese with English abstract)[本文引用:1]
 , 杨峰
, 杨峰



{kind=link}
{kind=link}
{kind=link}