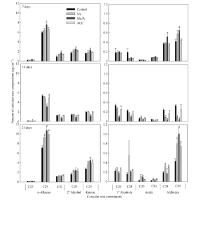
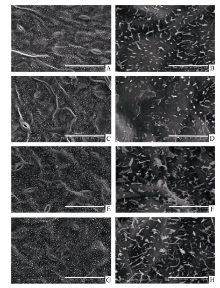
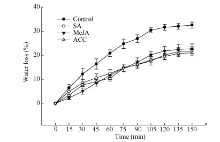
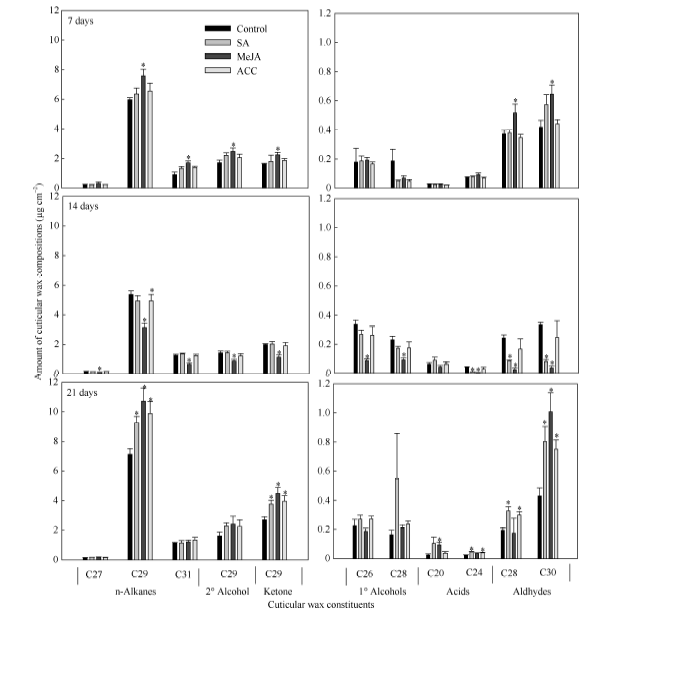
关键词:甘蓝型油菜; 角质层蜡质; 水杨酸(SA); 茉莉酸甲酯(MeJA); 乙烯(ETH) Effects of SA, MeJA, and ACC on Leaf Cuticular Wax Constituents, Structure and Permeability in Brassica napus LI Shuai, ZHAO Qiu-Ling, PENG Yang, XU Yi, LI Jia-Na, NI Yu* College of Agronomy and Biotechnology, Southwest University, Chongqing 400715, China Fund:This study was supported by the Fundamental Research Funds for the Central Universities (XDJK2014B037), the Chongqing Basic and Advanced Research Project (cstc2016jcyjA0170), and the National Natural Science Foundation of China (31000122). AbstractCuticular waxes are related to plant adaptation to environment stress. In the current study, Brassica napus cv. Zhongshuang 11 grown in the soil treated with adding solutions of SA (200 μmol L-1), MeJA (100 μmol L-1), and ACC (200 μmol L-1) wax, were sampled at the five-leaf stage to clarify the effects of SA, MeJA and ETH on adjusting plant cuticular wax deposition. The leaves were used to analyze the amounts of total cuticular wax and wax constituents, wax crystal structure, and cuticular permeability. The amounts of n-alkanes, secondary alcohols, ketones, aldehydes and total cuticular waxes increased significantly when compared with the control at seven days after MeJA treatment, whereas the amounts of all wax constituents and total cuticular waxes significantly decreased at 14 days after MeJA treatment. The application of SA and ACC had no significant influence on cuticular wax deposition at 7 and 14 days after treatments, except for an amount reduction of primary alcohol, aldehyde and unknown constituents for SA treatment at 14 days after treatment. At 21 days after SA, MeJA, and ACC application, the amounts of total cuticular wax, n-alkanes, ketones and aldehyde significantly increased. C29 n-alkane, C29 ketone, and C30 aldehyde were the main cuticular wax constituents induced by SA, MeJA, and ACC application, implying that n-alkane, ketone and aldehyde might be related to the resistance to stresses induced by these signal molecules. Scan electric microscope analysis indicted that the rod-shape structure of cuticular wax in leaf surface reduced and some rods melted under SA treatment. MeJA and ACC application increased the distribution density of wax crystalloids. The cuticular wax deposition and crystal structure alteration reduced the cuticular permeability and delayed the leaf water loss. The specific increase of C29 n-alkane might be the main reason for reducing leaf water loss in B. napus.
Keyword: Brassica napus L; Cuticular wax; Salicylic acid; Methyl jasmonic acid; Ethylene Show Figures Show Figures
图1 SA、MeJA和ACC处理对油菜叶角质层蜡质主要组分含量的影响数据柱上* 表示在P < 0.05水平上差异显著。Fig. 1 Effects of SA, MeJA, and ACC treatments on contents of major individual leaf cuticular wax constituents in B. napusBars with asterisks are significantly different at P < 0.05 according to Student’ s t-test.
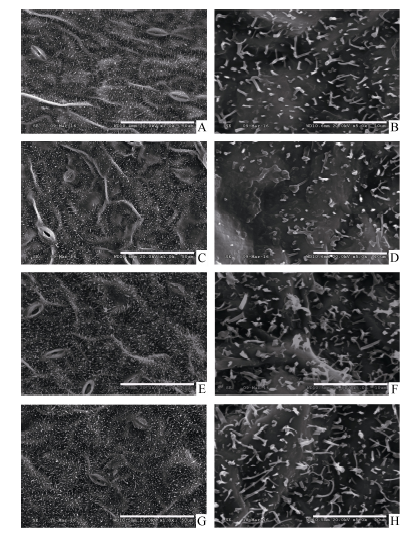
图2 SA、MeJA和ACC处理对油菜叶表皮蜡质晶体结构的影响A~B: 对照; C~D: SA处理; E~F: MeJA处理; G~H: ACC处理; A、C、E、G: 标尺=100 μ m; B、D、F、H: 标尺=10 μ m。Fig. 2 Effects of SA, MeJA, and ACC treatment on the crystalloid structures of leaf cuticular wax in B. napusA-B: control; C-D: SA; E-F: MeJA; G-H: ACC; A, C, E, and G: bar=100 μ m; B, D, F, and H: bar=10 μ m.
4 结论外源SA、MeJA及ACC处理能够诱导甘蓝型油菜中双11叶片角质层蜡质的沉积, 降低角质层渗透性, 减缓叶片的水分散失, 其中C29烷的特异性增加是造成叶片失水率降低的主要原因。蜡质组分中烷类、酮类以及醛类可能与SA、JA和ETH所介导的抗(耐)性反应密切相关。 The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
Kosma DK, BourdenxB, BernardA, Parsons EP, LüS, JoubèsJ, Jenks MA. The impact of water deficiency on leaf cuticle lipids of Arabidopsis. Plant Physiol, 2009, 151: 1918-1929[本文引用:2]
[2]
NiY, Xia RE, Li JN. Changes of epicuticular wax induced by enhanced UV-B radiation impact on gas exchange in Brassica napus. Acta Physiol Plant, 2014, 36: 2481-2490[本文引用:2]
Cajustea JF, González-Cand elasaL, VeyratA, García-Breijo F J, Reig-Arminana J, Lafuente M T. Epicuticular wax content and morphology as related to ethylene and storage performance of ‘Navelate’ orange fruit. Postharvest Biol Tec, 2010, 55: 29-35[本文引用:2]
[6]
CurryE. Effects of 1-MCP applied postharvest on epicuticular wax of apples ( Malus domesticaBorkh. ) during storage. J Sci Food Agric, 2008, 88: 996-1006[本文引用:1]
[7]
MackováJ, VaškováM, MacekP, HronkováM, SchreiberL, ŠantrůčekJ. Plant response to drought stress simulated by ABA application: changes in chemical composition of cuticular waxes. Environ Exp Bot, 2013, 86: 70-75[本文引用:1]
[8]
Seo PJ, Lee SB, Suh MC, Park MJ, Go YS, Park CM. The MYB96 transcription factor regulates cuticular wax biosynthesis under drought conditions in Arabidopsis. Plant Cell, 2011, 23: 1138-1152[本文引用:2]
[9]
Baker EA. The influence of environment on leaf wax development in Brassica oleracea var. gemmifera. New Phytol, 1974, 73: 955-966[本文引用:1]
[10]
Grant RH, Heisler GM, GaoW, JenksM. Ultraviolet leaf reflectance of common urban trees and the prediction of reflectance from leaf surface characteristics. Agric For Meteorol, 2003, 120: 127-139[本文引用:1]
[11]
Khan MI, FatmaM, Per TS. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front Plant Sci, 2015, 6: 462-478[本文引用:1]
[12]
Cheong JJ, Yang DC. Methyl jasmonate as a vital substance in plants. Trends Genet, 2003, 19: 409-413[本文引用:1]
[13]
Morgan PW, Drew MC. Ethylene and plant responses to stress. Physiol Plant, 1997, 100: 620-630[本文引用:1]
[14]
LeideJ, Hildebrand tU, VoggG, RiedererM. The positional sterile ( ps) mutation affects cuticular transpiration and wax biosynthesis of tomato fruits. J Plant Physiol, 2011, 168: 871-877[本文引用:1]
[15]
SinghK, Foley RC, Onate-SanchezL. Transcription factors in plant defense and stress responses. Curr Opins Plant Biol, 2002, 5: 430-436[本文引用:1]
[16]
BrounP, PoindexterP, OsborneE, Jiang CZ, Riechmann JL. WIN1, a transcriptional activator of epidermal wax accumulation in Arabidopsis. Proc Natl Acad Sci USA, 2004, 101: 4706-4711[本文引用:1]
[17]
AharoniA, DixitS, JetterR, Thoenes E, van Arkel G, Pereira A. The SHINE clade of AP2 domain transcription factors activates wax biosynthesis, alters cuticle properties, and confers drought tolerance when overexpressed in Arabidopsis. Plant Cell, 2004, 16: 2463-2480[本文引用:1]
[18]
NiY, Guo YJ, WangJ, Xia RE, Wang XQ, AshG, Li JN. Responses of physiological indexes and leaf epicuticular waxes of Brassica napus to Sclerotinia sclerotiorum infection. Plant Pathol, 2014, 63: 174-184[本文引用:1]
[19]
Kosma DK, Nemacheck JA, Jenks MA, Williams CE. Changes in properties of wheat leaf cuticle during interactions with Hessian fly. Plant J, 2010, 63: 31-43[本文引用:1]
[20]
GarbayB, Tautu MT, CostaglioliP. Low level of pathogenesis-related protein 1 mRNA expression in 15-day-old Arabidopsis cer6-2 and cer2 eceriferum mutants. Plant Sci, 2007, 172: 299-305[本文引用:1]
Boughton AJ, HooverK, Felton GW. Methyl jasmonate application induces increased densities of gland ular trichomes on tomato, Lycopersicon esculentum. J Chem Ecol, 2005, 31: 2211-2216[本文引用:1]
[24]
Holloway PJ, Brown GA, Baker EA, Macey M J K. Chemical composition and ultrastructure of the epicuticular wax in three lines of Brassica napus(L). Chem Phys Lipids, 1977, 19: 114-127[本文引用:1]
[25]
WengH, MolinaI, ShockeyJ, BrowseJ. Organ fusion and defective cuticle function in a lacs1 lacs2 double mutant of Arabidopsis. Planta, 2010, 231: 1089-1100[本文引用:1]
[26]
BourdenxB, BernardA, DomergueF, PascalS, LegerA, RobyD, PerventM, VileD, Haslam RP, Napier JA, LessireR, JoubesJ. Overexpression of Arabidopsis ECERIFERUM1 promotes wax very-long-chain alkane biosynthesis and influences plant response to biotic and abiotic stresses. Plant Physiol, 2011, 156: 29-45[本文引用:1]
[27]
Zhang JY, Broeckling CD, Sumner LW, Wang ZY. Heterologous expression of two Medicago truncatula putative ERF transcription factor genes, WXP1 and WXP2, in Arabidopsis led to increased leaf wax accumulation and improved drought tolerance, but differential response in freezing tolerance. Plant Mol Biol, 2007, 64: 265-278[本文引用:1]
[28]
Oliveira A FM, Meirelles ST, SalatinoA. Epicuticular waxes from caatinga and cerrado species and their efficiency against water loss. An Acad Bras Cienc, 2003, 75: 431-439[本文引用:1]
[29]
YangM, Yang QY, Fu TD, Zhou YM. Overexpression of the Brassica napus BnLAS gene in Arabidopsis affects plant development and increases drought tolerance. Plant Cell Rep, 2011, 30: 373-388[本文引用:1]
[30]
VoggG, FischerS, LeideJ, EmmanuelE, JetterR, Levy AA, RiedererM. Tomato fruit cuticular waxes and their effects on transpiration barrier properties: functional characterization of a mutant deficient in a very-long-chain fatty acid beta-ketoacyl- CoA synthase. J Exp Bot, 2004, 55: 1401-1410[本文引用:1]
[31]
GlazebrookJ. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu Rev Phytopathol, 2005, 43: 205-227[本文引用:1]
[32]
Thomma B P H J, EggermontK, Penninckx I A M A, Mauch-ManiB, VogelsangR, Cammue B PA, BroeckaertW. Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Arabidopsis are essential for resistance to distinct microbial pathogens. Proc Natl Acad Sci USA, 1998, 95: 15107-15111[本文引用:1]



{kind=link}
{kind=link}
{kind=link}