关键词:紫芽茶树; 基因表达分析; 总儿茶素; 花青素; 相关性分析 Correlation Analysis between Total Catechins (or Anthocyanins) and Expression Levels of Genes Involved in Flavonoids Biosynthesis in Tea Plant with Purple Leaf ZHOU Tian-Shan1, WANG Xin-Chao2, YU You-Ben1, XIAO Yao1, QIAN Wen-Jun1, XIAO Bin1,*, YANG Ya-Jun2,* 1College of Horticulture, Northwest A&F University, Yangling 712100, China
2Key Laboratory of Tea Plant Biology and Resource Utilization, Ministry of Agriculture / Tea Research Institute, Chinese Academy of Agricultural Sciences, Hangzhou 310008, China
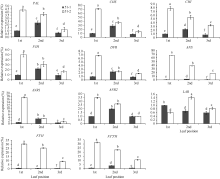
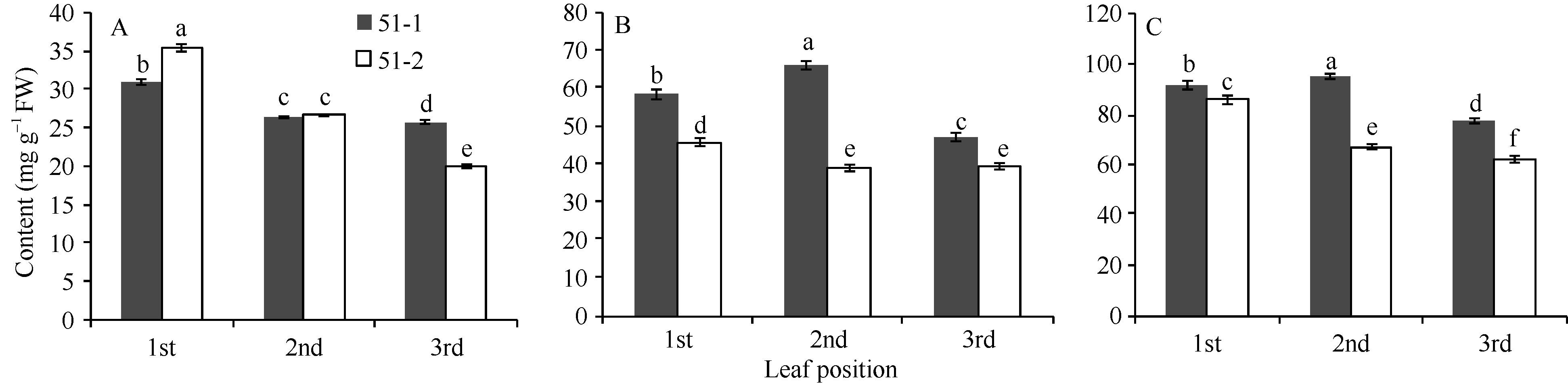
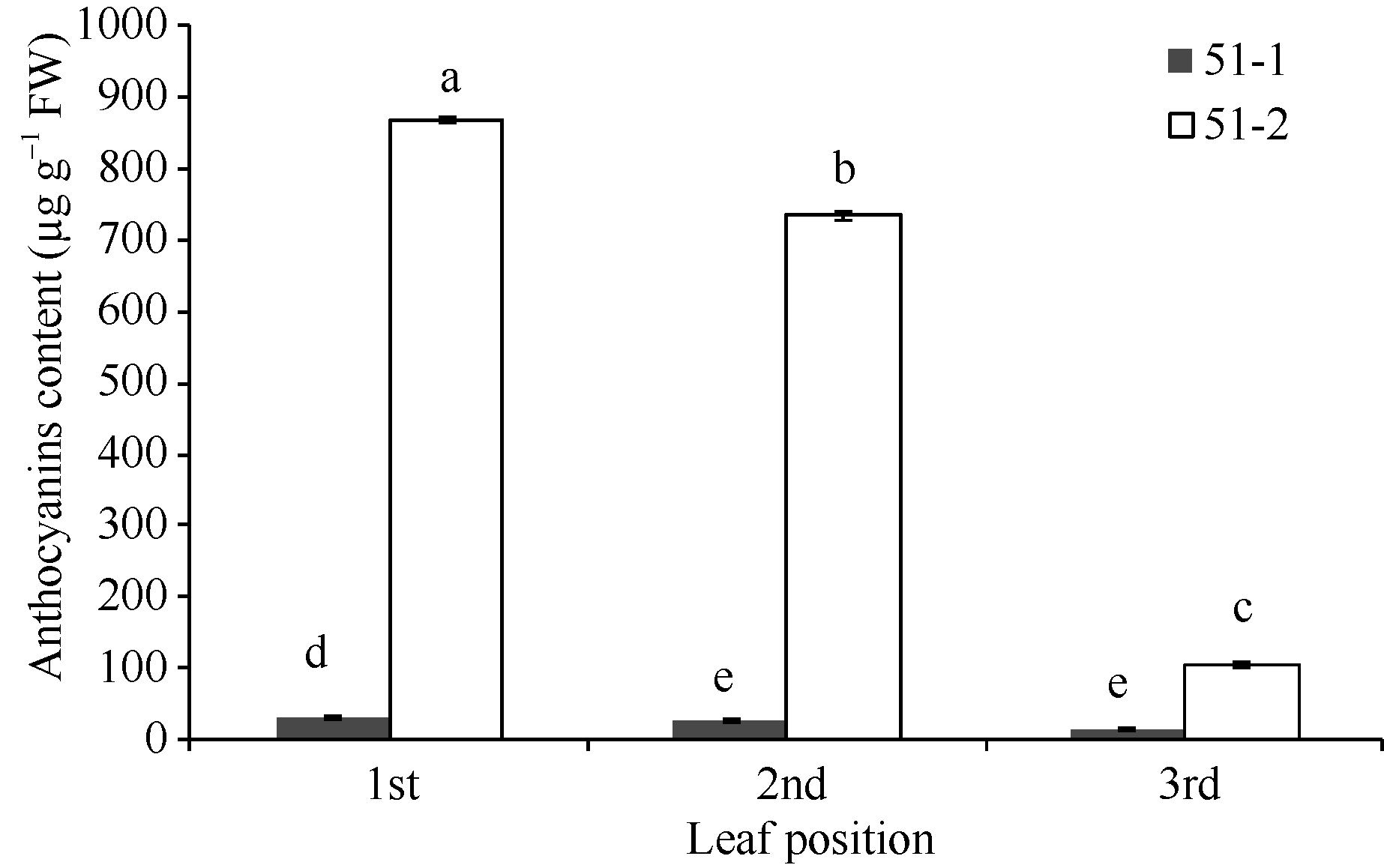
Fund:This study was supported by the China Agriculture Research System (CARS-23) AbstractFlavan-3-ols (aka catechins) and Anthocyanins found in large amounts in tea plant with purple leaf are synthesized through flavonoids metobolic pathway. To investigate the metobolic flux of flavanoids biosynthetic pathway in tea plant with purple leaf, we employed a tea plant with purple leaf and a tea plant with green leaf both from Mei-Tan-Tai-Cha, to examine the expression profiles of related genes involved in flavonoids biosynthesis intensively and determine the concentrations of catechins and anthocynins. The correlation between total catechins (or anthocyanin) and the expression levels of related genes were analyzed. The results indicated that the expression levels of related genes ( PAL, CHS, CHI, F3H, DFR, ANS, ANR1, ANR2, F3’H, and F3’5’H) were up-regulated in purple leaves as compared with those in the green leaves. The purple leaf also had higher concentration of anthocyanins than the green leaf, while the green leaf was richer in total catechins. In purple leaves, the expression levels of related genes (except LAR) were highly correlated with both concentrations of total catechins ( r = 0.84-0.99) and anthocyanins ( r = 0.72-1.00). In contrast, there was only a high correlation between the expression levels of related genes (except LAR and F3’ H) and the concentration of total catechines ( r = 0.64-0.77) in green leaves.
Keyword:Tea plant with purple leaf; Gene expression analysis; Total catechins; Anthocyanins; Correlation analysis Show Figures Show Figures
表2 紫色芽叶与对照色泽 Table 2 Values of L, a, and b for purple leaves and control
样品 Sample
L
a
b
绿色芽叶(对照) Green leaf (control) (51-1)
第一叶 1st leaf
45.72± 0.24 e
-14.99± 0.37 b
28.11± 0.44 e
第二叶 2nd leaf
46.62± 0.09 e
-15.66± 0.24 a
28.85± 0.49 f
第三叶 3rd leaf
42.42± 0.40 d
-14.97± 0.54 b
23.92± 0.63 d
紫色芽叶 Purple leaf (51-2)
第一叶 1st leaf
30.30± 0.38 a
4.71± 0.33 e
3.91± 0.29 a
第二叶 2nd leaf
33.71± 0.26 b
2.47± 0.09 d
10.44± 0.28 b
第三叶 3rd leaf
34.92± 0.36 c
-5.64± 0.24 c
15.15± 0.31 c
The lightness, L, represents the darkest black at L = 0 and the brightest white at L = 100. The red/green opponent colors are represented along the aaxis, with green at negative avalues and red at positive avalues. The yellow/blue opponent colors are represented along the b axis, with blue at negative b values and yellow at positive b values. Duncan’ s multiple range test (P< 0.05, n= 20). L表示明暗度, L = 0时表示黑色, L = 0时表示白色; a表示红绿, a为正值表示红色, a为负值表示绿色; b表示黄蓝, b为正值表示黄色, b为负值表示蓝色。采用Duncan’ s 多重比较方法分析(P< 0.05, n= 20)。
表2 紫色芽叶与对照色泽 Table 2 Values of L, a, and b for purple leaves and control
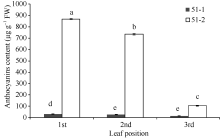
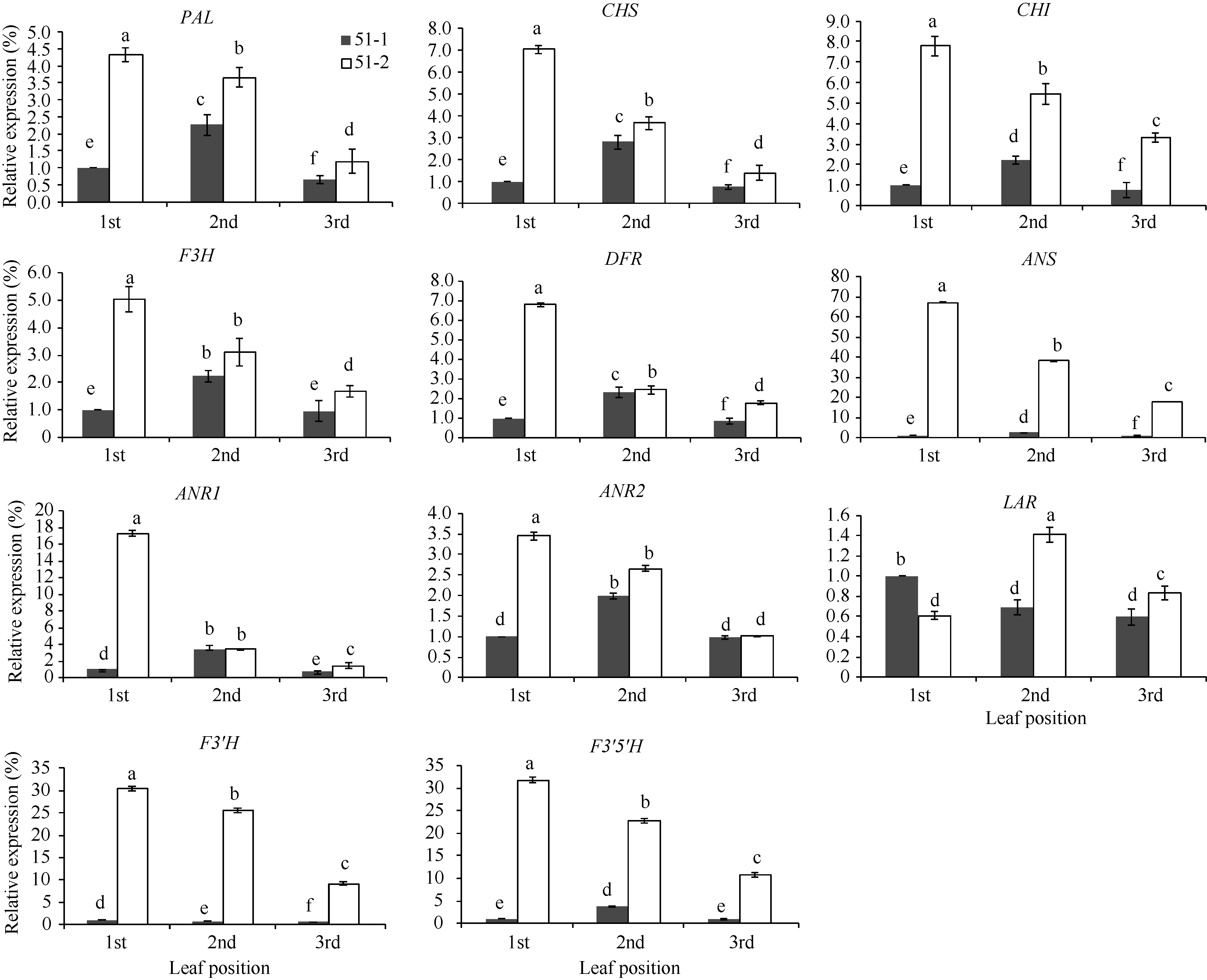
图4 紫色芽叶中类黄酮生物合成相关基因相对表达量 51-1: 对照; 51-2: 紫色芽叶。采用Duncan’ s 多重比较方法分析(P< 0.05, n= 3)。Fig. 4 Expression levels of related genes involved in flavonoids biosynthesis in tea plant with purple leaf 51-1: control; 51-2: purple leaf. Duncan’ s multiple range test (P< 0.05, n= 3).
宛晓春. 茶叶生物化学(第3版). 北京: 中国农业出版社, 2003. pp 9-30Wan X C. Tea Biochemistry, 3rd Edn. Beijing: China Agriculture Press, 2003. pp 9-30(in Chinese)[本文引用:1]
[2]
Shirley BW. Flaconoid biosynthesis: a colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol, 2001, 126: 485-493[本文引用:4]
[3]
SeitzC, EderC, DeimlB, KellnerS, MartensS, ForkmannG. Cloning, functional identification and sequence analysis of flavonoid 3’-hydroxylase and flavonoid 3’, 5’-hydroxylase cDNAs reveals independent evolution of flavonoid 3’, 5’-hydroxylase in the Asteraceae family. Plant Mol Biol, 2006, 61: 365-381[本文引用:4]
[4]
AshiharaH, Deng WW, MullenW, CrozierA. Distibution and biosynthesis of flavan-3-ols in Camellia sinensis seedlings and expression of genes encoding biosynthetic enzymes. Phytochemistry, 2010, 71: 559-566[本文引用:4]
[5]
吕海鹏, 费旭元, 梁名志, 王立波, 林智. 茶树特异品种“紫娟”中的花青素组分分析. 食品科学, 2012, 33(22): 203-206Lü HP, Fei XY, Liang MZ, Wang LB, LinZ. Analysis of anthocyanins constituents in the special tea germplasms of Zijuan. Food Sci, 2012, 33(22): 203-206 (in Chinese with English abstract)[本文引用:1]
KowalczykE, KrzesinskiP, KuraM, SzmigielB, BlaszczykJ. Anthocyanins in medicine. Pol J Pharmacol, 2003, 55: 699-702[本文引用:1]
[8]
NakaishiH, MatsumotoH, TominagaS, HirayamaM. Effects of black currant anthocyanoside intake on dark adaptation and VDT work-induced transient refractive alteration in healthy humans. Altern Med Rev, 2000, 5: 553-562[本文引用:1]
[9]
TeraharaN, TakedaY, NesumiA, HondaT. Anthocyanins from red flower tea (Benibana-cha), Camellia sinensis. Phytochemistry, 2001, 56: 359-361[本文引用:3]
[10]
SaitoT, HonmaD, TagashiraM, Kand aT, NesumiA, Maeda-YamamotoM. Anthocyanins from new red leaf tea “Sunrouge”. J Agric Food Chem, 2011, 59: 4779-4782[本文引用:3]
[11]
包云秀, 夏丽飞, 李友勇, 梁名志. 茶树新品种“紫娟”. 园艺学报, 2008, 35: 934Bao YX, Xia LF, Li YY, Liang MZ. A new tea tree cultivar ‘Zijuan’. Acta Hort Sin, 2008, 35: 934 (in Chinese with English abstract)[本文引用:1]
[12]
Meada-YamamotoM, SaitoT, NesumiA, TokudaY, EmaK, HonmaD, OginoA, MonobeM, MurakamiA, MurakamiA, TachibanaH. Chemical analysis and acetylcholinesterase inhibitory effect of anthocyanin-rich red leaf tea (cv. Sunrouge). J Sci Food Agric, 2012, 92: 2379-2386[本文引用:3]
[13]
蔡丽, 梁名志, 夏丽飞, 陈林波, 孙云南. “紫娟”茶外观表象差异研究. 西南农业学报, 2010, 23: 700-703CaiL, Liang MZ, Xia LF, Chen LB, Sun YN. Study on exterior appearance difference of ‘Zijuan’. Southwest China J Agric Sci, 2010, 23: 700-703 (in Chinese with English abstract)[本文引用:1]
[14]
季鹏章, 梁名志, 宋维希, 蒋会兵, 马琳, 王丽, 矣兵. 茶树珍稀品种“紫娟”的叶片色素含量与叶色变化的关系研究. 西南农业学报, 2010, 23: 1860-1863Ji PZ, Liang MZ, Song WX, Jiang HB, MaL, YiB. Relationship between changes of pigments content and leaf color changing in ‘Zijuan’ (Camellia sinensis var. assamica). Southwest China J Agric Sci, 2010, 23: 1860-1863 (in Chinese with English abstract)[本文引用:1]
[15]
Jiang LH, Shen XJ, ShojiT, Kand aT, Zhou JC, Zhao LM. Characterization and activity of anthocyanins in Zijuan tea (Camellia sinensis var. kitamura). J Agric Food Chem, 2013, 61: 3306-3310[本文引用:2]
[16]
陈林波, 夏丽飞, 孙云南, 梁明志, 张正竹, 李叶云, 宛晓春. 特异茶树品种“紫娟”叶色转变的基因表达差异分析. 茶叶科学, 2012, 32: 59-65Chen LB, Xia LF, Sun YN, Liang MZ, Zhang ZZ, Li YY, Wan XC. Analysis of differential gene expression on specific tea cultivar ‘Zijuan’ for leaf color changing. J Tea Sci, 2012, 32: 59-65 (in Chinese with English abstract)[本文引用:1]
[17]
Livaka KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 2001, 25: 402-408[本文引用:1]
[18]
尹军峰, 许勇泉, 袁海波, 余书平, 韦坤坤, 陈建新, 汪芳, 吴荣梅. 名优绿茶鲜叶摊放过程中主要生化成分的动态变化. 茶叶科学, 2009, 19: 102-110Yin JF, Xu YQ, Yuan HB, Yu SP, Wei KK, Chen JX, WangF, Wu RM. Dynamic change of main biochemical components of premium green tea fresh leaves during spreading. J Tea Sci, 2009, 19: 102-110 (in Chinese with English abstract)[本文引用:1]
[19]
Kerio LC, Wachira FN, Wanyoko JK, Rotich MK. Characterization of anthocyanins in Kenyan teas: extraction and identification. Food Chem, 2012, 131: 31-38[本文引用:1]
[20]
MatsumotoS, TakeuchiA, HayatsuM, KondoS. Molecular cloning of phenylalanine ammonium-lyase cDNA and classification of varieties and cultivars of tea plants (Camellia sinensis) using a tea PAL cDNA probe. Theor Appl Genet, 1994, 89: 671-675[本文引用:1]
[21]
Olsen KM, Lea US, SlimestadR, VerheulM, LilloC. Differential expression of four Arabidopsis PAL genes: PAL1 and PAL2 have functional specialization in abiotic environmental-triggered flavonoid synthesis. J Plant Physiol, 2008, 165: 1491-1499[本文引用:1]
[22]
SinghK, KumarS, RaniA, GulatiA, Ahuja PS. Phenylalanine ammonia-lyase (PAL) and cinnamate 4-hydroxylase (C4H) and catechins (flavan-3-ols) accumulation in tea. Funct Integr Genomic, 2009, 9: 125-134[本文引用:1]
[23]
Xiong LG, LiJ, Li YH, YuanL, Liu SQ, Huang JA, Liu ZH. Dynamic changes in catechin levels and catechin biosynthesis-related gene expression in albino tea plants (Camellia sinensis L. ). Plant Physiol Biochem, 2013, 71: 132-143[本文引用:1]
[24]
PangY, Abeysinghe IS, HeJ, HeX, HuhmanD, Mewan KM, Sumner LW, YunJ, Dixon RA. Functional characterization of proanthocyanidin pathway enzymes from tea and their application for metabolic engineering. Plant Physiol, 2013, 161: 1103-1116[本文引用:1]
 , 王新超
, 王新超



{kind=link}
{kind=link}
{kind=link}
{kind=link}