关键词:小麦; 簇毛麦; 白粉病; 小片段易位; 配子传递率 Transmission and Genetic Stability of No-homoeologous Small Fragment Wheat- Haynaldia villosa Translocation Chromosomes with Pm21in Various Cultivar Backgrounds of Common Wheat WANG Hai-Yan, XIAO Jin, YUAN Chun-Xia, XU Tao, YU Chun-Yan, SUN Hao-Jie, CHEN Pei-Du, WANG Xiu-E* State Key Laboratory of Crop Genetics and Germplasm Enhancement / Cytogenetics Institute, Nanjing Agricultural University / Jiangsu Collaborative Innovation Center for Modern Crop Production, Nanjing 210095, China Fund:This study was supported by the National High Technology Research Program (863 Program) of China (2011AA100101, 2011AA10010201), the National Natural Science Foundation of China (31201204), the Fundamental Research Funds for the Central Universities (KYZ201403) and the Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD) AbstractThe powdery mildew resistance gene Pm21comes from a diploid wheat related species, Haynaldia villosa. Two Pm21-carrying small fragment translocation lines, the terminal translocation line NAU418 and the small interstitial translocation line NAU419, have been developed. Both lines are highly resistant to powdery mildew and serve as new genetic resources for improvement of disease resistance. For understanding the transmission rate of the translocation chromosomes through male and female gametes and the genetic stabilities in different wheat genetic backgrounds, the two translocations were crossed to 12 common wheat varieties from different wheat growing areas of China. The F1 hybrids were then backcrossed as reciprocally. Chromosome configurations of the obtained F1s were analyzed by fluorescence in situhybridization (FISH) of the PMC at MI. It was found that the translocation chromosomes formed rod bivalents with their corresponding wheat chromosomes. Test crosses showed that the translocation chromosomes T1AS•1AL-6VS and T4BS•4BL-6VS-4BL could be transmitted to their offspring. The transmission frequency of T1AS•1AL-6VS was higher through female gametes an average of 33.20%, ranging from 0.08% to 50.98% than through male gametes an average of 23.75%, ranging from 0.14% to 45.07%. Similarly, the transmission frequency of T4BS•4BL-6VS-4BL was higher through female gametes an average of 42.90%, ranging from 29.17% to 52.38% than through male gametes an average of 21.45%, ranging from 7.69% to 47.06%. These results show that the translocated chromosomes could be transmitted through male and female gametes, while genetic background has influences on the transmission rate, especially through male gametes.
Keyword:Wheat; Haynaldia villosa; Powdery mildew; Small fragment translocation; Gamete transmission frequency Show Figures Show Figures
表1 NAU418和NAU419与不同品种的杂种F1 PMC MI染色体构型及自交结实率 Table 1 Chromosome configuration and seed-setting rates of F1 hybrids from crosses of NAU418 and NAU419 with different wheat varieties
母本 Female
父本 Male
观察细胞数 No. of PMCs observed
F1染色体配对构型 Chromosome configuration of F1
自交结实率Seed-setting rate (%)
单价体 Univalent
棒状二价体 Ring II
环状二价体 Rod II
二价体合计 Total II
NAU418
郑麦9023 Zhengmai 9023
37
0.00
1.78
19.22
21.00
95.74
周麦9823 Zhoumai 9823
61
0.03
2.08
18.90
20.98
99.08
绵阳26 Mianyang 26
33
0.06
3.33
17.63
20.96
96.71
石4185 Shi 4185
41
0.15
3.61
17.32
20.93
94.83
扬麦15 Yangmai 15
—
—
—
—
—
95.38
宁麦13 Ningmai 13
50
0.08
3.48
17.48
20.96
94.73
济麦22 Jimai 22
51
0.20
3.90
17.00
20.90
93.03
远丰175 Yuanfeng 175
23
0.17
4.52
16.39
20.91
92.30
川麦54 Chuanmai 54
48
0.25
2.98
17.90
20.88
98.81
南农0686 Nannong 0686
63
0.13
2.71
18.22
20.93
96.51
平均值 Mean
45
0.12
3.15
17.78
20.94
95.71
NAU419
郑麦9023 Zhengmai 9023
44
0.18
3.30
17.61
20.91
91.43
周麦9823 Zhoumai 9823
—
—
—
—
—
95.06
石4185 Shi 4185
79
0.33
3.86
16.97
20.84
92.49
扬麦15 Yangmai 15
66
0.12
3.32
17.62
20.94
91.47
宁麦13 Ningmai 13
—
—
—
—
—
90.87
济麦22 Jimai 22
54
0.11
3.83
17.11
20.94
90.20
远丰175 Yuanfeng 175
48
0.44
3.46
17.32
20.78
91.47
扬麦11 Yangmai 11
31
0.19
3.77
17.13
20.90
93.10
川麦54 Chuanmai 54
39
0.00
2.92
18.08
21.00
94.73
烟农19 Yannong 19
63
0.13
2.71
18.23
20.94
92.71
南农0686 Nannong 0686
55
0.00
2.11
18.89
21.00
93.70
平均值 Mean
53
0.17
3.25
17.66
20.92
92.48
The ploidy of all parents and their F1 is 2n = 42. Symbol — indicates data not available. PMC: pollen mother cell. 所有亲本和F1的倍性均为2n = 42。— 表示数据缺失。
表1 NAU418和NAU419与不同品种的杂种F1 PMC MI染色体构型及自交结实率 Table 1 Chromosome configuration and seed-setting rates of F1 hybrids from crosses of NAU418 and NAU419 with different wheat varieties
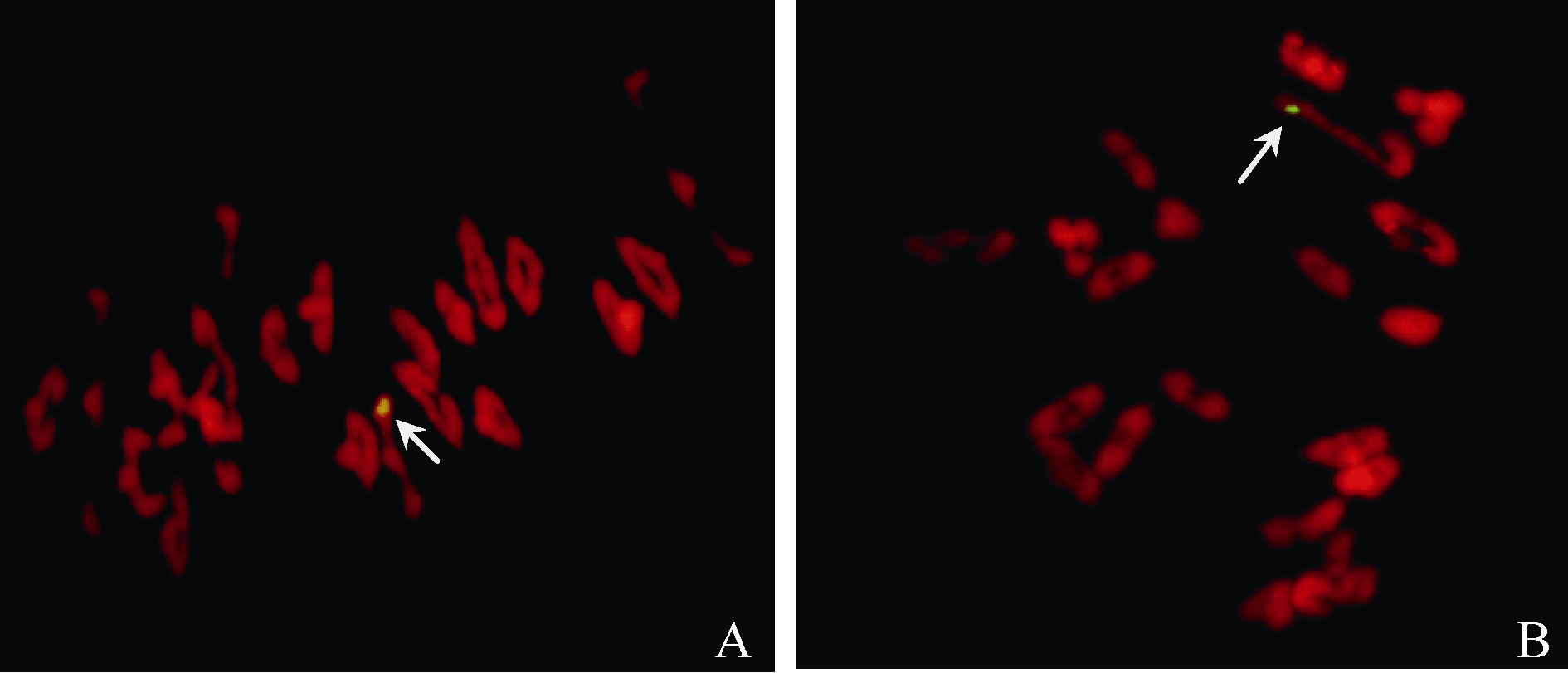
图2 与Pm21基因紧密连锁的EST标记CINAU15在NAU418和NAU419回交群体部分单株的扩增结果Fig. 2 Electrophoresis of Pm21-specific EST marker CINAU15 in partial plants from the NAU418 and NAU419 backcross populations
表2 Table 2 表2(Table 2)
表2 T1AS• 1AL-6VS (NAU418)和T4BS• 4BL-6VS-4BL (NAU419)易位染色体在不同小麦背景中通过雌雄配子的传递率 Table 2 Gamete transmission frequencies of T1AS• 1AL-6VS (NAU418) and T4BS• 4BL-6VS-4BL (NAU419) in different wheat backgrounds
杂交、回交的小麦品种 Wheat variety in hybrid and backcross
雌配子传递 Female gamete transmission
雄配子传递 Male gamete transmission
Pm21株 Plants with Pm21
观察株数 Observed plants
传递率 Transmission frequency (%)
χ 2
Pm21株 Plants with Pm21
观察株数 Observed plants
传递率 Transmission frequency (%)
χ 2
NAU418
烟农19 Yannong 19
26
51
50.98
0.00
3
38
7.89* *
25.28
远丰175 Yuanfeng 175
35
73
47.95
0.05
32
71
45.07
0.51
扬麦15 Yangmai 15
42
97
43.30
1.48
20
52
38.46
2.33
周麦9823 Zhoumai 9823
8
21
38.10
0.76
12
54
22.22* *
15.57
石4185 Shi 4185
24
63
38.10
3.10
9
27
33.33
2.37
郑麦9023 Zhengmai 9023
20
60
33.33*
6.02
21
51
41.18
1.25
绵阳26 Mianyang 26
24
80
30.00* *
12.00
31
78
39.74
2.88
南农0686 Nannong 0686
13
49
26.53* *
9.88
4
50
8.00* *
33.62
宁麦13 Ningmai 13
17
72
23.61* *
19.01
15
95
15.79* *
42.11
川麦54 Chuanmai 54
4
50
8.00* *
33.62
8
46
17.39* *
18.28
济麦22 Jimai 22
—
—
—
—
7
50
14.00* *
24.50
NAU419
扬麦11 Yangmai 11
22
42
52.38
0.02
9
35
25.71* *
7.31
石4185 Shi 4185
22
47
46.81
0.07
8
17
47.06
0.00
扬麦15 Yangmai 15
16
36
44.44
0.25
5
48
10.42* *
28.52
郑麦9023 Zhengmai 9023
6
15
40.00
0.27
9
38
23.68* *
9.50
济麦22 Jimai 22
31
60
51.67
0.02
3
39
7.69* *
26.26
远丰175 Yuanfeng 175
32
76
42.11
1.59
14
74
18.92* *
27.36
南农0686 Nannong 0686
6
18
33.33
1.39
8
48
16.67*
20.02
川麦54 Chuanmai 54
6
13
46.15
0.00
—
—
—
—
宁麦13 Ningmai 13
7
24
29.17
3.38
—
—
—
—
χ 20.05 = 3.84; χ 20.01 = 6.64. * and * * indicate significant difference between the observed transmission frequency and the expected value (50%) at the 0.05 and 0.01 probability levels, respectively. — indicates data not available. χ 20.05 = 3.84; χ 20.01 = 6.64。* 和* * 分别表示传递率与理论值(50%)在0.05和0.01水平差异显著。— 表示数据缺失。
表2 T1AS• 1AL-6VS (NAU418)和T4BS• 4BL-6VS-4BL (NAU419)易位染色体在不同小麦背景中通过雌雄配子的传递率 Table 2 Gamete transmission frequencies of T1AS• 1AL-6VS (NAU418) and T4BS• 4BL-6VS-4BL (NAU419) in different wheat backgrounds
谢超杰, 杨作民. 孙其信. 小麦抗白粉病基因. 西北植物学报, 2003, 23: 822-829Xie CJ, Yang ZM, Sun QX. Resistance genes to powdery mildew in wheat. Acta Bot Boreali-Occident, 2003, 23: 822-829 (in Chinese with English abstract)[本文引用:1]
[2]
GradzielewskaA. The genus Dasypyrum: 2. Dasypyrum villosum, a wild species used in wheat improvement. Euphytica, 2006, 152: 441-454[本文引用:1]
[3]
Chen PD, Qi LL, ZhouB, Zhang SZ, Liu DJ. Development and molecular cytogenetic analysis of wheat-Haynaldia villosa 6VS/6AL translocation lines specifying resistance to powdery mildew. Theor Appl Genet, 1995, 91: 1125-1128[本文引用:1]
[4]
Qi LL, Cao MS, Chen PD, Li WL, Liu DJ. Identification, mapping, and application of polymorphic DNA associated with resistance gene Pm21 of wheat. Genome, 1996, 39: 191-197[本文引用:1]
[5]
Cao AZ, Xing LP, Wang XY, Yang XM, WangW, Sun YL, QianC, Ni JL, Chen YP, Liu DJ, Wang XE, Chen PD. Serine/threonine kinase gene Stpk-V, a key member of powdery mildew resistance gene Pm21, confers powdery mildew resistance in wheat. Proc Natl Acad Sci USA, 108: 7727-7732[本文引用:1]
[6]
Chen PD, You CF, HuY, Chen SW, ZhouB, Cao AZ, Wang XE. Radiation-induced translocations with reduced Haynaldia villosa chromatin at the Pm21 locus for powdery mildew resistance in wheat. Mol Breed, 2013, 31: 477-484[本文引用:2]
[7]
李桂萍, 陈佩度, 张守忠, 赵和. 小麦-簇毛麦6VS/6AL易位染色体对小麦农艺性状的影响. 植物遗传资源学报, 2011, 12: 744-749Li GP, Chen PD, Zhang SZ, ZhaoH. Effects of the 6VS/6AL translocation chromosome on agronomic characteristics of wheat. J Plant Genet Resour, 2011, 12: 744-749 (in Chinese with English abstract)[本文引用:4]
[8]
Smith JD. The effect of chromosome number on competitive ability of hexaploid wheat gametophytes. Can J Genet Cytol, 1963, 5: 200-226[本文引用:1]
[9]
Efremova TT, Maystrenko OI, Laykova LI. Development of alien substitution lines of wheat with rye chromosome 5R. Cereal Res Commun, 1996, 24: 33-39[本文引用:1]
[10]
Whelan ED, Conner P R L, Thomas J B. Transmission of a wheat alien chromosome translocation with resistance to the wheat curl mite in common wheat, Triticum aestivum L. Can J Genet Cytol, 1986, 28: 294-297[本文引用:1]
[11]
张文俊, 景健康, 胡含. 黑麦6R染色体在小麦背景中的传递. 遗传学报, 1995, 22: 211-216Zhang WJ, Jing JK, HuH. The transmission of rye chromosome 6R in wheat background. Acta Genet Sin, 1995, 22: 211-216 (in Chinese with English abstract)[本文引用:2]
[12]
王海燕, 赵仁慧, 袁春霞, 张守忠, 肖进, 王秀娥. 小麦-簇毛麦T4DL•4VS易位染色体在不同背景中的遗传稳定性及其在配子中的传递. 麦类作物学报, 2013, 33: 13-17Wang HY, Zhao RH, Yuan CX, Zhang SZ, XiaoJ, Wang XE. Transmission of T4DL•4VS chromosome through gametes and its genetic stability in different genetic background. J Triticeae Crops, 2013, 33: 13-17 (in Chinese with English abstract)[本文引用:2]
[13]
ZhangP, LiW, FriebeB, Gill BS. Simultaneous painting of three genomes in hexploid wheat by BAC-FISH. Genome, 2004, 47: 979-987[本文引用:1]
[14]
Sharp PJ, CaoS, DesaiS, Gale MD. The isolation characterization and application in the Triticeae of a set of wheat RFLP probes identifying each homoeologous chromosome arm. Theor Appl Genet, 1989, 78: 342-348[本文引用:1]
[15]
Cao AZ, Wang XE, Chen YP, Zou XW, Chen PD. A sequence-specific PCR marker linked with Pm21 distinguishes chromosomes 6AS, 6BS, 6DS of Triticum aestivum and 6VS of Haynaldia villosa. Plant Breed, 2006, 125: 201-205[本文引用:1]
[16]
张悦, 林志珊, 曹保久, 郭义强, 王美蛟, 叶兴国, 辛志勇, 徐琼芳, 郭世华. 2Ai-2染色体在小麦部分同源染色体代换背景中的遗传. 作物学报, 2009, 35: 424-431ZhangY, Lin ZS, Cao BJ, Guo YQ, Wang MJ, Ye XG, Xin ZY, Xu QF, Guo SH. Genetic behaviour of Thinopyrum intermedium chromosome 2Ai-2 in different wheat chromosome substitution backgrounds of group 2. Acta Agron Sin, 2009, 35: 424-431 (in Chinese with English abstract)[本文引用:1]
[17]
刘琳, 邓光兵, 易玲, 李林, 赵柳笛, 龙海, 潘志芬, 余懋群. 偏凸山羊草6Mv染色体在不同四川小麦品种中的传递. 应用与环境生物学报, 2010, 16: 50-53LiuL, Deng GB, YiL, LiL, Zhao LD, LongH, Pan ZF, Yu MQ. Transmission of chromosome 6Mv from Aegilops ventricosa in Sichuan wheat varieties. Chin J Appl Environ Biol, 2010, 16: 50-53 (in Chinese with English abstract)[本文引用:1]





{kind=link}
{kind=link}