关键词:谷子; CIPK基因; 逆境 Expression Analysis of Two CIPK genes under Abiotic Stress in Foxtail Millet YU Ai-Li1,2, ZHAO Jin-Feng1,2, WANG Gao-Hong2, DU Yan-Wei2, LI Yan-Fang2, ZHANG Zheng2, GUO Er-Hu2, LIANG Ai-Hua1,* 1Insitute of Biotechnology, Shanxi University, Taiyuan 030006, China
2 Millet Research Institute, Shanxi Academy of Agricultural Sciences / Shanxi Key Laboratory of Genetic Resources and Breeding in Minor Crops, Changzhi 046011, China
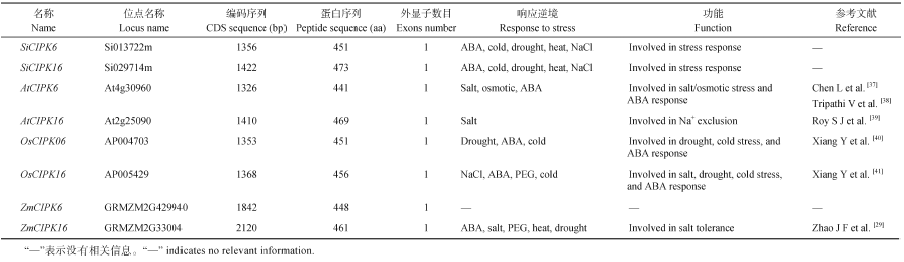
Fund:This study was supported by the grants from Natural Science Foundation of Shanxi Province (2015011071), the Key Laboratory Open Fund Project of Bio-engineering in Shanxi Provincial, and the Enhance Technology Independent Innovation Ability Project in Shanxi Academy of Agricultural Sciences (2015ZZCX-09) AbstractCIPK (CBL interacting protein kinase) is a type of serine or threonine protein kinases, which plays an important role in response to stress. In this study, we identified two CIPKgenes designated as SiCIPK6and SiCIPK16from foxtail millet ( Setaria italica) genome using bioinformatics methods. The sequence analysis showed that SiCIPK6 has a length of 1994 bp in the genome, encoding 513 amino acids residues, and SiCIPK16 is 1885 bp, encoding 473 amino acids residues. These two genes have no alternative splicing and intron. The characters predicted based on the bioinformatics analysis revealed that the protein sequences and structure of the two SiCIPK genes were very conservative just like CIPKgenes in other species. Real-time PCR analysis discovered that the expression of SiCIPK6 and SiCIPK16 was up-regulated by ABA, cold, heat, drought and salt stress, respectively. The expression was strongly induced by ABA, drought and salt treatments for SiCIPK6, and by cold, drought and heat treatments for SiCIPK16. The semi-quantitative PCR analysis showed that SiCIPK6 and SiCIPK16 were expressed at the jointing, booting and filling stages, and induced by drought stress in the corresponding growth period. Foxtail millet CIPKgenes reported in this study would enrich CIPK members in plant kingdom and provides important information for further elucidating the function and mechanisms of the CBL/CIPK network system responsive to stresses in foxtail millet.
Keyword:Foxtail millet; Calcineurin B-like-interacting protein kinase gene; Stress Show Figures Show Figures
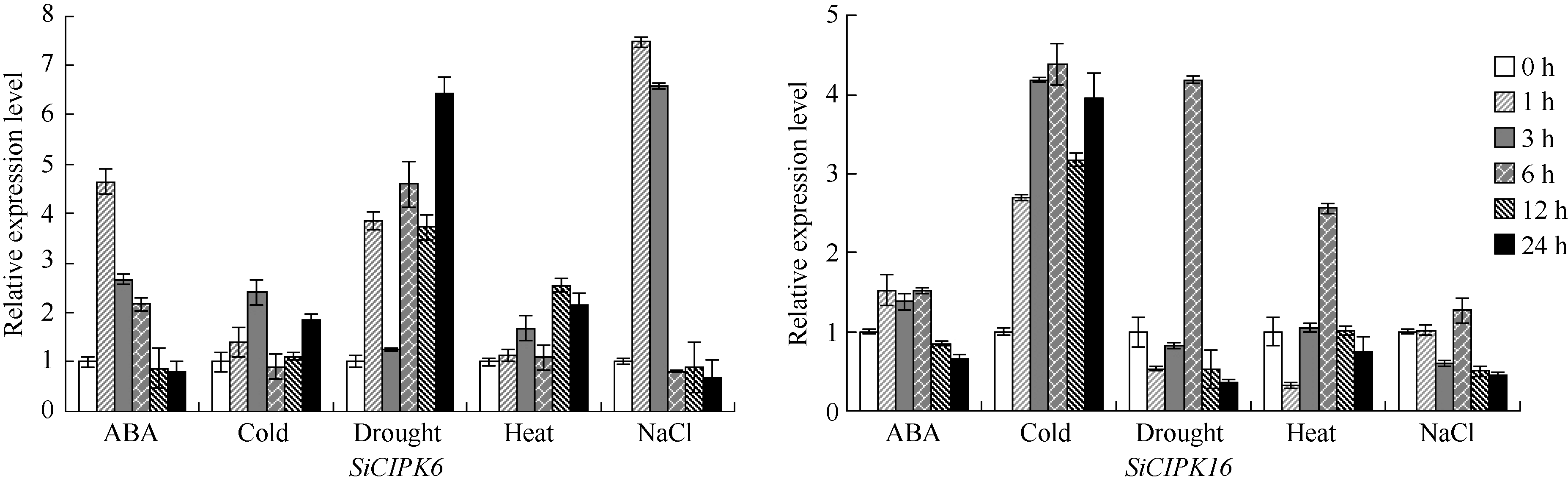
图1SiCIPK6和SiCIPK16在ABA、低温、干旱、高温和盐处理下的Real-time PCR表达分析Fig. 1 Real-time PCR analysis of expression levels of the SiCIPK6andSiCIPK16under ABA, cold, dehydration, heat, and salt stresses treatments
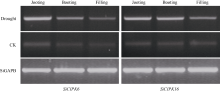
图2SiCIPK6和SiCIPK16基因在不同生育期自然脱水胁迫下的表达分析Fig. 2 Electrophoretogram displaying the loading patterns for expression ofSiCIPK6 and SiCIPK16 under dehydration stresses in different growth periods
图3 SiCIPK6、SiCIPK16与其他已知CIPK氨基酸序列比对相同氨基酸残基用黑色表示, 相似氨基酸残基用灰色表示(≥ 60% similarity); 磷酸化或自磷酸化位点用星号表示。保守的激活环和C端调控域的NAF结构域用连字符标出。 The amino acids with an entire homology are shown by a black background, and those shared non-identical conserved identity by a gray background (≥ 60% similarity); Autophosphorylated or phosphorylated sites are marked with asterisk. The activation loop and the conserved NAF motif are marked with hyphens.Fig. 3 Sequences alignment of SiCIPK6, SiCIPK16 and other known CIPKs
智慧, 牛振刚, 贾冠清, 柴杨, 李伟, 王永芳, 李海权, 陆平, 白素兰, 刁现民. 谷子干草饲用品质性状变异及相关性分析. 作物学报, 2012, 38: 800-807ZhiH, Niu ZG, Jia GQ, ChaiY, LiW, Wang YF, Li HQ, LuP, Bai SL, Diao XM. Variation and correlation analysis of hay forage quality traits of foxtail millet [Setaria italica (L. ) Beauv. ]. Acta Agron Sin, 2012, 38: 800-807 (in Chinese with English abstract)[本文引用:1]
[2]
Devos KM, Wang ZM, BealesJ, SasakiT, Gale MD. Comparative genetic maps of foxtail millet (Setaria italica) and rice (Oryza sativa). Theor Appl Genet, 1998, 96: 63-68[本文引用:1]
[3]
JayaramanA, PuranikS, Rai NK, VidapuS, Sahu PP, LataC, Prasad M. cDNA-AFLP analysis reveals differential gene expression in response to salt stress in foxtail millet (Setaria italica L. ). Mol Biotechnol, 2008, 40: 241-251[本文引用:1]
[4]
ZhaoD, OosterhuisD, BednarzC. Influence of potassium deficiency on photosynthesis, chlorophyll content and chloroplast ultrastructure of cotton plants. Photosynthetica, 2001, 9: 103-109[本文引用:1]
[5]
ZhouJ, WangJ, BiY, WangL, TangL, YuX, OhtaniM, DemuraT, ZhugeQ. Overexpression of PtSOS2 enhances salt tolerance in transgenic poplars. Plant Mol Biol Report, 2014, 32: 185-197[本文引用:1]
[6]
Wang WX, VinocurB, AltmanA. Plant responses to drought, salinity and extreme temperature: towards genetic engineering for stress tolerance. Planta, 2003, 2: 1-14[本文引用:1]
Choi WG, ToyotaM, Kim SH, HillearyR, GilroyS. Salt stress-induced Ca2+waves are associated with rapid, longdistance root-to-shoot signaling in plants. Proc Natl Acad Sci USA, 2014, 111: 6497-6502[本文引用:1]
[9]
BatisticO, KudlaJ. Analysis of calcium signaling pathways in plants. Biochim Biophys Acta, 2012, 1820: 1283-293[本文引用:2]
[10]
KolukisaogluU, WeinlS, BlazevicD, BatisticO, KudlaJ. Calcium sensors and their interacting protein kinases: genomics of the Arabidopsis and rice CBL-CIPK signaling networks. Plant Physiol, 2004, 134: 43-58[本文引用:1]
[11]
ZhangH, YangB, Liu WZ, LiH, WangL, WangB, DengM, LiangW, DeyholosMK, Jiang YQ. Identification and characterization of CBL and CIPK gene families in canola (Brassica napus L. ). BMC Plant Biol, 2014, 14: 8[本文引用:1]
[12]
Kleist TJ, Spencley AL, LuanS. Comparative phylogenomics of the CBL-CIPK calcium-decoding network in the moss Physcomitrella, Arabidopsis, and other green lineages. Front Plant Sci, 2014, 5: 1-17[本文引用:2]
[13]
沈金秋, 郑仲仲, 潘伟槐, 潘建伟. 植物CBL-CIPK信号系统的功能及其作用机理. 植物生理学报, 2014, 50: 641-650Shen JQ, Zheng ZZ, Pan WH, Pan JW. Functions and action mechanisms of CBL-CIPK signaling system in plants. Plant Physiol J, 2014, 50: 641-650 (in Chinese with English abstract)[本文引用:2]
[14]
Wu SJ, DingL, Zhu JK. SOS1: a genetic locus essential for salt tolerance and potassium acquisition. Plant Cell, 1996, 8: 617-627[本文引用:1]
[15]
LiuJ, Zhu JK. An Arabidopsis mutant that requires increased calcium for potassium nutrition and salt tolerance. Proc Natl Acad Sci USA, 1997, 94: 14960-14964[本文引用:1]
[16]
GongD, GuoY, Jagendorf AT, Zhu JK. Biochemical characterization of the Arabidopsis protein kinase SOS2 that functions in salt tolerance. Plant Physiol, 2002, 130: 256-264[本文引用:1]
[17]
HalfterU, IshitaniM, Zhu JK. The Arabidopsis SOS2 protein kinase physically interacts with and is activated by the calcium-binding protein SOS3. Proc Natl Acad Sci USA, 2000, 97: 3735-3740[本文引用:1]
[18]
Quintero FJ, OhtaM, ShiH, Zhu JK, Pardo JM. Reconstitution in yeast of the Arabidopsis SOS signaling pathway for Na+ homeostasis. Proc Natl Acad Sci USA, 2002, 99: 9061-9066[本文引用:1]
[19]
Qiu QS, GuoY, Quintero FJ, Pardo JM, Schumaker KS, Zhu JK. Regulation of vacuolar Na+/H+ exchange in Arabidopsis thaliana by the salt-overly-sensitive (SOS) pathway. J Biol Chem, 2004, 279: 207-215[本文引用:1]
[20]
XuJ, Li HD, Chen LQ, WangY, Liu LL, HeL, Wu WH. A protein kinase, interacting with two calcineurin B-like proteins, regulates K+ transporter AKT1 in Arabidopsis. Cell, 2006, 125: 1347-1360[本文引用:1]
[21]
Lee SC, Lan WZ, Kim BG, LiL, Cheong YH, Pand ey GK, LuG, Buchanan BB, LuanS. A protein phosphorylation/dephosphorylation network regulates a plant potassium channel. Proc Natl Acad Sci USA, 2007, 104: 15959-15964[本文引用:1]
[22]
Hu HC, Wang YY, Tsay YF. AtCIPK8, a CBL-interacting protein kinase, regulates the low-affinity phase of the primary nitrate response. Plant J, 2009, 57: 264-278[本文引用:1]
[23]
DengX, ZhouS, HuW, FengJ, ZhangF, ChenL, HuangC, LuoQ, HeY, YangG, HeG. Ectopic expression of wheat TaCIPK14, encoding a calcineurin B-like protein-interacting protein kinase, confers salinity and cold tolerance in tobacco. Physiol Plant, 2013, 149: 367-377[本文引用:1]
[24]
DengX, HuW, WeiS, ZhouS, ZhangF, HanJ, ChenL, LiY, FengJ, FangB, LuoQ, LiS, LiuY, YangG, HeG. TaCIPK29, a CBL-interacting protein kinase gene from wheat, confers salt stress tolerance in transgenic tobacco. PLoS One, 2013, 8: e69881[本文引用:1]
[25]
ChenX, HuangQ, ZhangF, WangB, WangJ, ZhengJ. ZmCIPK21, a maize CBL-interacting kinase, enhances salt stress tolerance in Arabidopsis thaliana. Int J Mol Sci, 2014, 15: 14819-14834[本文引用:1]
[26]
LiJ, LongY, Qi GN, Xu ZJ, WuWH, WangY. The Os-AKT1 channel is critical for K+ uptake in rice roots and is modulated by the rice CBL1-CIPK23 complex. Plant Cell, 2014, 26: 3387-3402[本文引用:1]
[27]
CuellarT, PascaudF, Verdeil JL, TorregrosaL, Adam-Blondon A F, Thibaud J B, Sentenac H, Gaillard I. A grapevine Shaker inward K+ channel activated by the calcineurin B-like calcium sensor 1-protein kinase CIPK23 network is expressed in grape berries under drought stress conditions. Plant J, 2010, 61: 58-69[本文引用:1]
[28]
CuellarT, AzeemF, AndrianteranagnaM, PascaudF, Verdeil JL, SentenacH, ZimmermannS, GaillardI. Potassium transport in developing fleshy fruits: the grapevine inward K+ channel VvK1. 2 is activated by CIPK-CBL complexes and induced in ripening berry flesh cells. Plant J, 2013, 73: 1006-1018[本文引用:1]
[29]
Zhao JF, Sun ZF, ZhengJ, Guo XY, Dong ZG, Huai JL, Gou MY, He JG, Jin YS, Wang JH, Wang GY. Cloning and characterization of a novel CBL-interacting protein kinase from maize. Plant Mol Biol, 2009, 69: 661-674[本文引用:2]
[30]
赵晋锋, 余爱丽, 田岗, 杜艳伟, 郭二虎, 刁现民. 谷子CBL基因鉴定及其在干旱、高盐胁迫下的表达分析. 作物学报, 2013, 39: 360-367Zhao JF, Yu AL, TianG, Du YW, Guo EH, Diao XM. Identification of CBL genes from foxtail millet (Setaria italica [L. ] Beauv. ) and its expression under drought and salt stresses. Acta Agron Sin, 2013, 39: 360-367 (in Chinese with English abstract)[本文引用:1]
[31]
ZhengJ, Zhao JF, Tao YZ, Wang JH, Liu YJ, Fu JJ, JinY, GaoP, Zhang JP, Bai YF, Wang GY. Isolation and analysis of water stress induced genes in maize seedlings by subtractive PCR and cDNA macroarray. Plant Mol Biol, 2004, 55: 807-823[本文引用:1]
[32]
余琴鸯, 尹恒, 安利佳, 李文利. 一个快速响应干旱的F-box基因的克隆和表达分析. 作物学报, 2014, 40: 1531-1539Yu QY, YinH, An LJ, Li WL. Cloning and expression analysis of a calcium-dependent protein kinase gene SiCDPK1 in Setaria italica. Acta Agron Sin, 2014, 40: 1531-1539 (in Chinese with English abstrac)[本文引用:1]
[33]
Zhang JP, Liu TS, Fu JJ, ZhuY, Jia JP, ZhengJ, Zhao YH, ZhangY, Wang GY. Construction and application of EST library from Setaria italica in response to dehydration stress. Genomics, 2007, 90: 121-131[本文引用:1]
[34]
SambrookJ, RussellD. Molecular Cloning: A Laboratory Manual, 3rd Edn. New York: Cold Spring Harbor Laboratory Press, 2001. pp 581-585[本文引用:1]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using Real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods, 2001, 25: 402-408[本文引用:1]
[37]
ChenL, Wang QQ, ZhouL, RenF, Li DD, Li XB. Arabidopsis CBL-interacting protein kinase (CIPK6) is involved in plant response to salt/osmotic stress and ABA. Mol Biol Rep, 2013, 40: 4759-4767[本文引用:1]
[38]
TripathiV, ParasuramanB, LaxmiA, ChattopadhyayD. CIPK6, a CBL-interacting protein kinase is required for development and salt tolerance in plants. Plant J, 2009, 58: 778-790[本文引用:1]
[39]
Roy SJ, HuangW, Wang XJ, EvrardA, Schmöckel SM, Zafar ZU, TesterM. A novel protein kinase involved in Na (+) exclusion revealed from positional cloning. Plant Cell Environ, 2013, 36: 553-568[本文引用:1]
[40]
XiangY, HuangY, XiongL. Characterization of stressresponsive CIPK genes in rice for stress tolerance improvement. Plant Physiol, 2007, 144: 1416-1428[本文引用:1]
[41]
赵晋锋, 余爱丽, 王高鸿, 田岗, 王寒玉, 杜艳伟, 常海霞. 植物CBL/CIPK网络系统逆境应答研究进展. 中国农业科技导报, 2011, 13(4): 32-38Zhao JF, Yu AL, Wang GH, TianG, Wang HY, Du YW, Chang HX. Progress of CBL/CIPK signal system in response to stresses in plant. J Agric Sci Technol, 2011, 13(4): 32-38 (in Chinese with English abstract)[本文引用:2]






{kind=link}
{kind=link}
{kind=link}