关键词:转基因小麦; NADP苹果酸酶; 净光合速率; 气孔导度 Photosynthetic Characteristics of Transgenic Wheat Expressing Maize C4-Type NADP-ME Gene WANG Yong-Xia1,2, DU Xin-Hua1,2, XU Wei-Gang1,2,*, QI Xue-Li2, LI Yan2, WANG Hui-Wei2, HU Lin2 1Nanjing Key Laboratory of Crop Genetics and Germplasm Enhancement, Nanjing Agricultural University, Nanjing 210095, China
2 Henan Provincial Laboratory of Wheat Biology / Wheat Research Institute, Henan Academy of Agricultural Sciences, Zhengzhou 450002, China
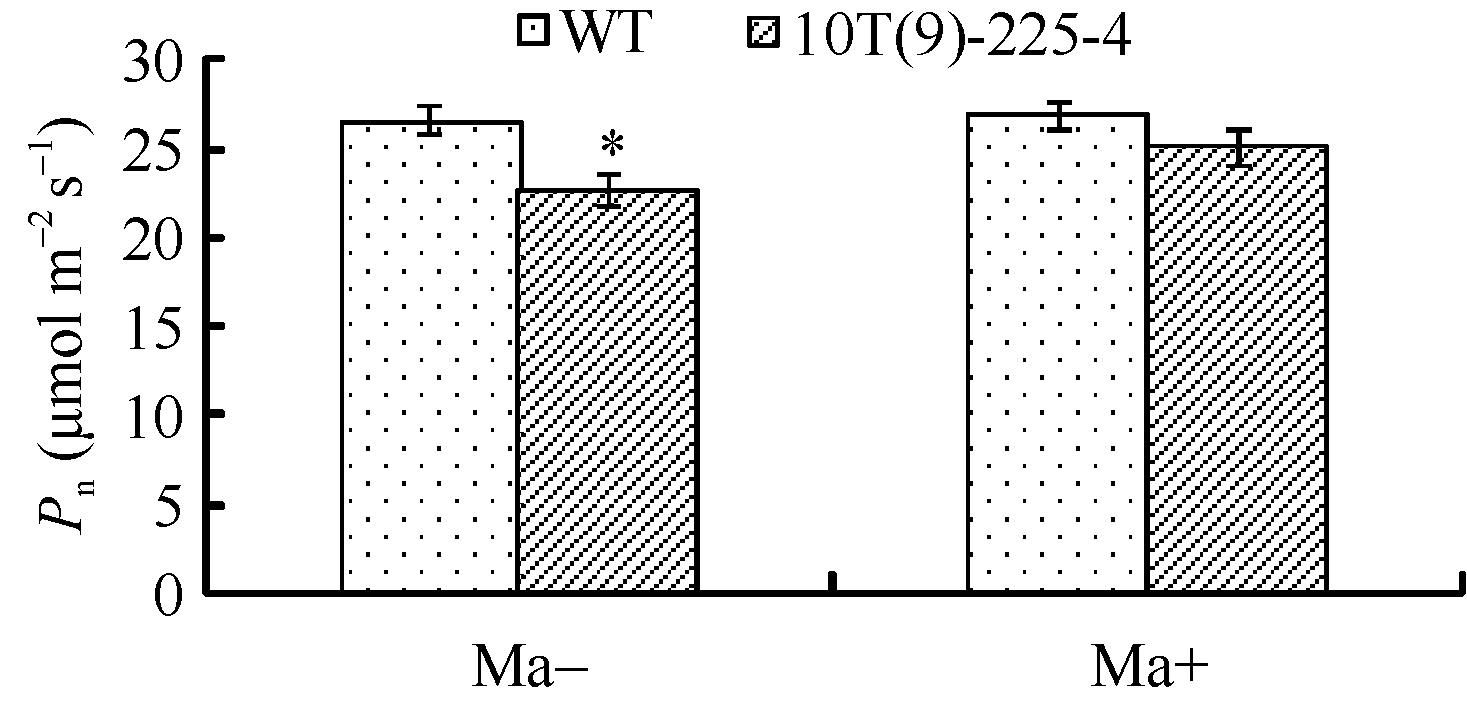
Fund:This study was supported by the National GMO Program of China (2011ZX08002-003) and the China Agriculture Research System (CARS-3-1-9) AbstractTo explore the physiological characteristics of the transgenic wheat expressing maize C4-type NADP-ME, we introduced NADP-ME into the C3 crop wheat by using particle bombardment transformation. Two transgenic wheat lines (10T(9)-1-1, 10T(9)-225-4) and parental control (Zhoumai 23) were used to study molecular characteristics and photosynthesis property, to reveal the mechanism. The results showed that the NADP-ME sequence was integrated into wheat genome, and the transcription and translation were exactly same as expect. The enzyme activity of NADP-ME in flag leaf in transgenic plants were increased significantly than untransformed plants, for instance it was increased 1.33 and 1.13 times on the 7th day after flowering. Net photosynthetic rate ( Pn) of flag leaf in transgenic plants obviously decreased when compared to the untransformed plants. On the 7th day after anthesis, Pn of transgenic wheat decreased by 17.26% and 10.35%. The yield and 1000-grain weight were decreased than the control. Utilization efficiency on strong light utilizing and ability of CO2 assimilation in transgenic line 10T(9)-225-4 were significantly declined, photosynthesis rate was also decreased. Stomatal opening rate and stomatal conductance were significant decrease, malic acid content of transgenic wheat reducing 5.6% while pyruvate level is raised by 17.1%, and Pn of transgenic wheat can be restored by feeding with exogenous malate. Those results indicated that the transgenic wheat expressing maize NADP-ME gene showed lower photosynthetic characteristics than the control, the reason was maybe the decrease of stomatal aperture caused by decline of malic acid content.
Keyword:Transgenic wheat; NADP-dependent malic enzyme; Net photosynthetic rate; Stomata conductance Show Figures Show Figures
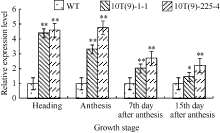
图2 转基因小麦及其对照(WT)不同生育期旗叶NADP-ME基因的相对表达量 内参基因为β -actin。* * 表示转基因植株与对照之间差异显著(P< 0.01)。Fig. 2 Relative expressions of NADP-MEin flag leaves of transgenic wheat lines and wild type (WT) at different stages Gene β -actinwas used as the internal reference. * * indicates significant difference between the transgenic plant and WT (P< 0.01).
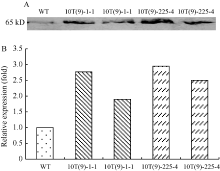
图3 T3代转基因株系Western杂交(A)和杂交图片灰度分析(B)结果 以WT的表达蛋白含量为1。Fig. 3 Western blot (A) and gray level analysis (B) of T3 transgenic lines The expression protein content of WT was 1.
图4 不同生育期转基因小麦及对照(WT)旗叶NADP-ME酶活性(A)和净光合速率(B)的变化 * 和* * 分别表示转基因植株与对照在P< 0.05和P< 0.01水平差异显著。Fig. 4 Dynamic changes of NADP-ME enzyme activity(A) and photosynthetic rate (B) in flag leaves of transgenic wheat lines and the wild type (WT) at different stages * and * * indicate significant difference between the transgenic plant and WT at P< 0.05 and P< 0.01 level, respectively.
表2 转NADP-ME基因小麦和非转基因对照的产量及农艺性状比较 Table 2 Comparison of yield related traits betweenNADP-ME transgenic wheat lines and WT
株系 Line
株高 Plant height (cm)
分蘖数 Spike number per plant
小穗数 Number per spike
单株产量 Grain yield per plant (g)
单穗粒数 Grain number per spike
千粒重 1000-grain weight (g)
WT
67.05± 3.03
8.60± 1.31
19.50± 1.28
19.03± 3.72
42.49± 2.62
50.06± 2.76
10T(9)-1-1
68.12± 3.11
8.60± 2.06
18.50± 1.15
16.92± 2.35*
40.62± 4.54
45.03± 2.43*
10T(9)-225-4
67.17± 1.97
8.86± 2.00
18.64± 0.98
17.03± 2.27*
40.26± 3.97
45.10± 2.17*
Data are shown as mean ± SD of 30 individual plants. * indicates significant difference between the transgenic lines and WT at P < 0.05. WT: wild. 数据为30个单株重复的平均值± 标准差。* 表示转基因株系与对照差异显著(P< 0.05)。
表2 转NADP-ME基因小麦和非转基因对照的产量及农艺性状比较 Table 2 Comparison of yield related traits betweenNADP-ME transgenic wheat lines and WT
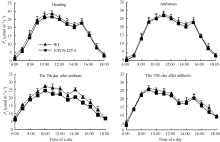
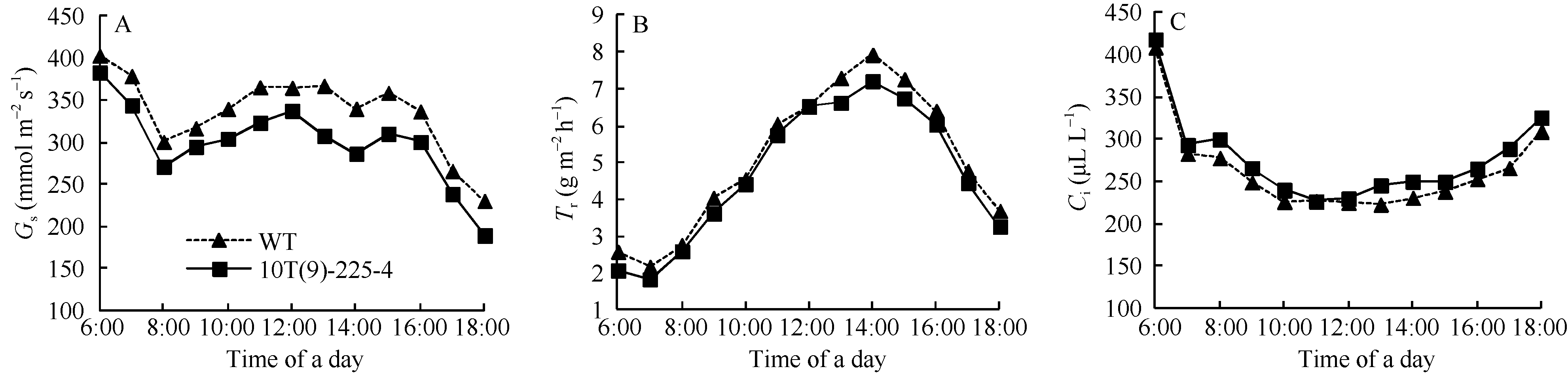
图5 转基因株系和对照(WT)不同时期旗叶净光合速率日变化曲线Fig. 5 Diurnal variation of photosynthetic rate of flag leaves in transgenic line and wild type (WT) at different stages
图6 转基因株系和对照(WT)不同时期旗叶Gs、Tr和Ci平均日变化曲线 图中数值均为4个生育期测定数值的平均值。Fig. 6 Average diurnal variation of Gs, Tr, and Ci of flag leaves in transgenic line and wild type (WT) at different stages Data are the mean of four growth stages.
表3 Table 3 表3(Table 3)
表3 转NADP-ME小麦不同生育时期的单位日光合总量 Table 3 Diurnal photosynthesis cumulative Pn in NADP-ME transgenic wheat lines at different growth stages (× 105 μ mol m-2)
材料 Line
抽穗期 Heading
开花期 Anthesis
花后第7天 7th day after anthesis
花后第15天 15th day after anthesis
平均 Average
WT
8.30± 0.63
7.83± 0.89
9.07± 0.57
8.48± 0.68
8.42± 0.66
10T(9)-225-4
7.63± 6.28
7.38± 0.75
7.80± 0.55*
7.88± 0.89
7.68± 0.73
Data are shown as Mean ± SD of 10 individual plants. * indicates significant difference between the transgenic lines and WT at P < 0.05. WT: wild type (Zhoumai 23). 数据为10个单株重复的平均值± 标准差。* 表示转基因株系与对照差异显著(P< 0.05)。
表3 转NADP-ME小麦不同生育时期的单位日光合总量 Table 3 Diurnal photosynthesis cumulative Pn in NADP-ME transgenic wheat lines at different growth stages (× 105 μ mol m-2)
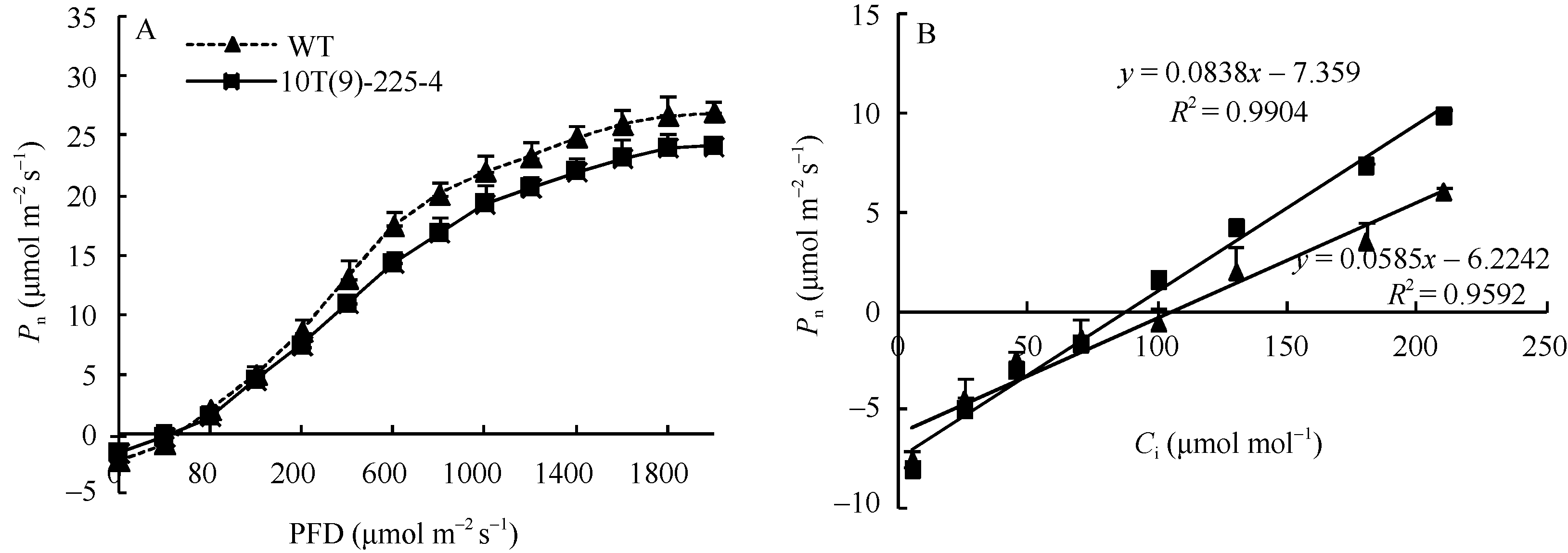
图7 转基因小麦旗叶净光合速率对光照强度(A)和胞间CO2浓度(C)的响应曲线Fig. 7 Response curves of photosynthesis to PPDF (A) and Ci (C) in transgenic wheat
表4 Table 4 表4(Table 4)
表4 转NADP-ME小麦和非转基因对照苹果酸、丙酮酸含量比较 Table 4 Comparison of malic acid and pyruvate acid content betweenNADP-ME transgenic wheat and WT
株系 Line
苹果酸含量 Malic acid content (mg g-1)
丙酮酸含量 Pyruvate acid content (mg g-1)
WT
0.45± 0.07
0.12± 0.02
10T(9)-225-4
0.41± 0.07*
0.15± 0.01*
Data are shown as mean ± SD of 5 individual flag leaves. * indicates significant difference between the transgenic lines and WT at P < 0.05. WT: wild type (Zhoumai 23). 数据为5片旗叶重复的平均值± 标准差。* 表示转基因株系与对照差异显著(P< 0.05)。WT: 受体对照(周麦23).
表4 转NADP-ME小麦和非转基因对照苹果酸、丙酮酸含量比较 Table 4 Comparison of malic acid and pyruvate acid content betweenNADP-ME transgenic wheat and WT
MiyaoM, MasumotoC, Miyazawa SI, FukayamaH. Lessons from engineering a single-cell C4 photosynthetic pathway into rice. J Exp Bot, 2011, 62: 3021-3029[本文引用:1]
[4]
Ruan CJ, Shao HB, Teixeira da Silva J A. A critical review on the improvement of photosynthetic carbon assimilation in C3 plants using genetic engineering. Crit Rev Biotechnol, 2012, 32: 1-21[本文引用:1]
[5]
Ku MS, AgarieS, NomuraM, FukayamaH, TsuchidaH, OnoK, MatsuokaM. High-level expression of maize phosphoenolpyruvate carboxylase in transgenic rice plants. Nat Biotechnol, 1999, 17: 76-80[本文引用:3]
FukayamaH, Hatch MD, TamaiT, TsuchidaH, SudohS, Furbank RT, MiyaoM. Activity regulation and physiological impacts of maize C4-specific phosphoenolpyruvate carboxylase overproduced in transgenic rice plants. Photosyn Res, 2003, 77: 227-239[本文引用:2]
[8]
陈绪清, 张晓东, 梁荣奇, 张立全, 杨凤萍, 曹鸣庆. 玉米C4型pepc基因的分子克隆及其在小麦的转基因研究. 科学通报, 2005, 49: 1976-1982Chen XQ, Zhang XD, Liang RQ, Zhang LQ, Yang FP, Cao MQ. Cloning maize C4 phosphoenolpyruvate carboxylase gene and transformation in wheat. Chin Sci Bull, 2005, 49: 1976-1982 (in Chinese with English abstract)[本文引用:1]
[9]
Band yopadhyayA, DattaK, ZhangJ, YangW, RaychaudhuriS, MiyaoM, Datta SK. Enhanced photosynthesis rate in genetically engineered indica rice expressing pepc gene cloned from maize. Plant Sci, 2007, 172: 1204-1209[本文引用:1]
[10]
张彬, 丁在松, 张桂芳, 石云鹭, 王金明, 方立锋, 郭志江, 赵明. 根癌农杆菌介导获得稗草 Ecppc转基因小麦的研究. 作物学报, 2007, 33: 356-362ZhangB, Ding ZS, Zhang GF, Shi YL, Wang JM, Fang LF, Guo ZJ, ZhaoM. Introduction of phosphoenolpyruvate carboxylase gene from Echinochloa crusgalli into wheat mediated by Agrobacterium tumefaciens. Acta Agron Sin, 2007, 33: 356-362 (in Chinese with English abstract)[本文引用:1]
[11]
张庆琛, 许为钢, 胡琳, 李艳, 张磊, 齐学礼. 玉米C4型全长pepc基因导入普通小麦的研究. 麦类作物学报, 2010, 30: 194-197Zhang QC, Xu WG, HuL, LiY, ZhangL, Qi XL. Development of transgenic wheat plants with maize C4-specific pepc gene by particle bombardment. J Triticeae Crops, 2010, 30: 194-197 (in Chinese with English abstract)[本文引用:1]
[12]
李艳, 许为钢, 胡琳, 张磊, 齐学礼, 张庆琛, 王根松. 玉米磷酸烯醇式丙酮酸羧化酶基因高效表达载体构建及其导入小麦的研究. 麦类作物学报, 2009, 29: 741-746LiY, Xu WG, HuL, ZhangL, Qi XL, Zhang QC, Wang GS. Construction of a high efficient expression vector for maize phosphoenolpyruvate carboxylase gene and its transformation in wheat. J Triticeae Crops, 2009, 29: 741-746 (in Chinese with English abstract)[本文引用:1]
[13]
HuL, LiY, XuW, ZhangQ, ZhangL, QiX, DongH. Improvement of the photosynthetic characteristics of transgenic wheat plants by transformation with the maize C4 phosphoenolpyruvate carboxylase gene. Plant Breed, 2012, 131: 385-391[本文引用:2]
[14]
SheriffA, MeyerH, RiedelE, Schmitt JM, LapkeC. The influence of plant pyruvate, orthophosphate dikinase on a C3 plant with respect to the intracellular location of the enzyme. Plant Sci, 1998: 136: 43-57[本文引用:1]
[15]
Ding ZS, Huang SH, Zhou BY, Sun XF, ZhaoM. Overexpression of phosphoenolpyruvate carboxylase cDNA from C4 millet (Seteria italica) increase rice photosynthesis and yield under upland condition but not in wetland fields. Plant Biotechnol Rep, 2013, 7: 155-163[本文引用:2]
[16]
Zhang HF, Xu WG, Wang HW, HuL, LiY, Qi XL, ZhangL, Li CX, HuaX. Pyramiding expression of maize genes encoding phosphoenolpyruvate carboxylase (PEPC) and pyruvate orthophosphate dikinase (PPDK) synergistically improve the photosynthetic characteristics of transgenic wheat. Protoplasma, 2014, 251: 1163-1173[本文引用:1]
[17]
Ku MS, WuJ, DaiZ, Scott RA, ChuC, Edwards GE. Photosynthetic and photorespiratory characteristics of Flaveria species. Plant Physiol, 1991, 96: 518-528[本文引用:2]
[18]
Häusler RE, RademacherT, LiJ, LipkaV, Fischer KL, SchubertS, KreuzalerF, Hirsch HJ. Single and double overexpression of C4-cycle genes had differential effects on the pattern of endogenous enzymes, attenuation of photorespiration and on contents of UV protectants in transgenic potato and tobacco plants. J Exp Bot, 2001, 52: 1785-1803[本文引用:1]
[19]
Huang XQ, Jiao DM, ChiW, Ku M S B. Characteristics of CO2 exchange and chlorophyll fluorescence of transgenic rice with C4 genes. Acta Bot Sin, 2002, 44: 405-412[本文引用:1]
[20]
TakeuchiY, AkagiH, KamasawaN, OsumiM, HondaH. Aberrant chloroplasts in transgenic rice plants expressing a high level of maize NADP-dependent malic enzyme. Planta, 2000, 211: 265-274[本文引用:2]
[21]
TsuchidaH, TamaiT, FukayamaH, AgarieS, NomuraM, OnoderaH, OnoK, NishizawaY, LeeB, HiroseS. High level expression of C4-specific NADP-malic enzyme in leaves and impairment of photoautotrophic growth in a C3 plant, rice. Plant Cell Physiol, 2001, 42: 138-145[本文引用:3]
[22]
ChiW, ZhouJ, ZhangF, WuN. Photosynthetic features of transgenic rice expressing sorghum C4 type NADP-ME. Acta Bot Sin, 2004, 46: 873-882[本文引用:3]
[23]
Laporte MM, ShenB, Tarczynski MC. Engineering for drought avoidance: expression of maize NADP-malic enzyme in tobacco results in altered stomatal function. J Exp Bot, 2002, 53: 699-705[本文引用:2]
[24]
王玉民. 玉米C4途径关键酶基因(PPDK、NADP-ME)的克隆及PPDK、PEPC在拟南芥中的表达分析. 河南农业大学博士学位论文, 河南郑州, 2011. pp 22-47Wang YM. Molecular Cloning of Maize C4 Key Enzyme Genes (PPDK and NADP-ME) and Expression Analysis of PPDK and PEPC in Arabidopsis. PhD Dissertation of Henan Agricultural University, Zhengzhou, China, 2011. pp 22-47 (in Chinese with English abstract)[本文引用:2]
[25]
杜西河. 玉米PEPC、PPDK和NADP-ME基因在拟南芥中的表达分析. 河南农业大学硕士学位论文, 2013. pp 17-21Du XH. Expression Analysis of Maize C4 Key Enzyme (Phosphoenolpyruvate Carboxylase, Pyruvate Orthophosphate Dikinase and NADP-Malic Enzyme) Genes in Arabidopsis. MS Thesis of Henan Agricultural University, Zhengzhou, China, 2013. pp 17-21 (in Chinese with English abstract)[本文引用:1]
[26]
陈昆松, 李方, 徐昌杰, 张上隆, 傅承新. 改良CTAB法用于多年生植物组织基因组DNA的大量提取. 遗传, 2004, 26: 529-531Chen KS, LiF, Xu CJ, Zhang SL, Fu CX. An efficient macro-method of genomic DNA isolation from Actinidia chinensis leaves. Hereditas (Beijing), 2004, 26: 529-531 (in Chinese with English abstract)[本文引用:1]
[27]
王兰兰, 何兴元, 陈玮, 李雪梅. 大气中O3、CO2浓度升高对蒙古栎叶片生长的影响. 中国环境科学, 2011, 31: 340-345Wang LL, He XY, ChenW, Li XM. Effects of elevated O3 or/and CO2 on growth in leaves of Quercus mongolica. China Environ Sci, 2011, 31: 340-345 (in Chinese with English abstract)[本文引用:1]
[28]
Boehringer MG. Methods of biochemical analysis and food analysis. Boehringer Mannheim GmbH, Biochemica, Mannheim, 1989[本文引用:1]
[29]
李琳, 李光兴, 代庆伟. 比色法快速分析苯丙酮酸含量的研究. 化学试剂, 2002, 24(1): 22-23LiL, Li GX, Dai QW. Analysis of the content of phenylpyruvic acid in mixture by colorimetry. Chem Reagents, 2002, 24(1): 22-23[本文引用:1]
[30]
JiB, ZhuS, JiaoD. A limited photosynthetic C4-microcycle and its physiological function in transgenic rice plant expressing the maize PEPC gene. Acta Bot Sin, 2004, 46: 542-551[本文引用:1]
[31]
SarahC, Julian MH. Integrating C4 photosynthesis into C3 crops to increase yield potential. Plant Biotechnol, 2012, 6: 1004[本文引用:1]
[32]
JiaoD, HuangX, LiX, ChiW, KuangT, ZhangQ, Ku MS, ChoD. Photosynthetic characteristics and tolerance to photo-oxidation of transgenic rice expressing C4 photosynthesis enzymes. Photosyn Res, 2002, 72: 85-93[本文引用:1]
[33]
Chen GY, Ye JY. Effects of oxaloacetate and malate on photosynthesis in leaves and in intact chloroplasts from spinach. Acta Phytophysiol Sin, 2001, 27: 478-482[本文引用:1]
[34]
朱素琴, 季本华, 焦德茂. 外源C4二羧酸对转玉米PEPC基因水稻C4光合途径的促进作用. 中国水稻科学, 2004, 18: 326-332Zhu SQ, Ji BH, Jiao DM. Promotive effect of exogenous C4-bicarboxylate on photosynthetic C4 pathway in transgenic rice plant expressing maize specific PEPC gene. Chin J Rice Sci, 2004, 18: 326-332[本文引用:1]
 , 杜新华
, 杜新华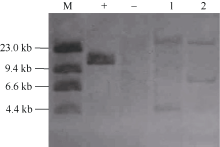









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}