关键词:甘蓝型油菜; 全基因组关联分析; 结角高度; 结角层厚度; 产量 Genome-Wide Association Analysis of Height of Podding and Thickness of Pod Canopy in Brassica napus LU Kun1,**, WANG Teng-Yue1,**, XU Xin-Fu1, TANG Zhang-Lin1, QU Cun-Ming1, HE Bin2, LIANG Ying1, LI Jia-Na1,* 1College of Agronomy and Biotechnology, Southwest University, Chongqing 400715, China
2 Agricultural Technology Extension Stationin Lincang City, Lincang 677000, China
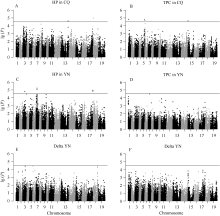
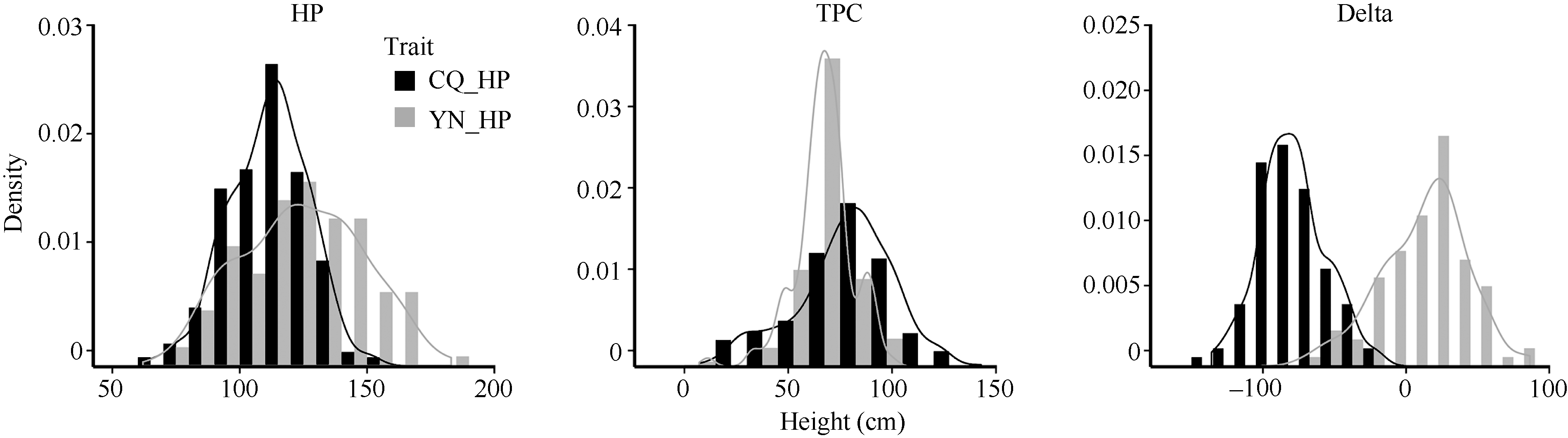
Fund:This study was supported by the National Basic Research Program of China (973 Program) (2015CB150201), the National Science Foundation of China (U1302266 and 31401412), the Program of Introducing International Super Agricultural Science and Technology (948 Program) (2011-G23), the Key Technologies Research and Development Program of China (2013BAD01B03-12) and the 111 Project (B12006) AbstractLayer of pod canopy is an important photosynthetic and seed storage part in rapeseed, providing important contribution to yield. In this study, 412 representative Brassica napus varieties (or lines) were genotyped using the Brassica 60 K Illumina Infinium SNP array by genome-wide association analysis of the height of podding (HP) and thickness of pod canopy (TPC). A total of 16 significant SNPs were identified, including two and four SNPs associated with HP and TPC in Chongqing, each of them explained 5.61-5.69% and 5.94-6.31% of phenotypic variation, respectively. Five and one significant SNPs accounting for 12.66-13.97% and 22.43% of the phenotypic variation for HP and TPC in Yunnan, respectively, were also detected. Three and one significant SNPs associated with the difference of HP and TPC between two environments were detected, explaining 17.33-20.32% and 29.05% of phenotypic variation, respectively. The latter SNP marker was located in the same linkage disequilibrium (LD) interval with one of significant SNPs related to TPC in Chongqing. Functional annotation of genes within the LD intervals containing significant markers showed that several genes involved in regulation of cell organization and biogenesis, floral meristem development, number of silique, and multicellular organismal development existed, such as NSN1, TPST, and SAC1, which might result in the variation of HP and TPC through affecting the growth and development of flower or silique in B. napus. These loci and genes could be regarded as important candidate regions and genes for HP and TPC of B. napus. The results lay the foundation for revealing the genetic basis and molecular mechanism for podding traits, and improving the yield per unit area of B. napus.
Keyword: Brassica napus; Genome-wide association analysis; Height of podding; Thickness of pod canopy; Yield Show Figures Show Figures
官春云. “农业科技创新与服务”笔谈: 依托科技创新与服务加快油菜产业发展方式转变. 湖南农业大学学报(社会科学版), 2010, 11(4): 1-3Guan CY. Pen talk on innovation and service of agricultural science and technology: accelerate change of rapeseed industry development mode based on scientific and technological innovation. J Hunan Agric Uni (Soc Sci), 2010, 11(4): 1-3 (in Chinese with English abstract)[本文引用:1]
[2]
王汉中. 我国油菜产业发展的历史回顾与展望. 中国油料作物学报, 2010, 32: 300-302Wang HZ. Review and future development of rapeseed industry in China. Chin J Oil Crop Sci, 2010, 32: 300-302[本文引用:2]
[3]
ÖzerH, OralE, DogruU. Relationships between yield and yield components on currently improved spring rapeseed cultivars. Turk J Agric For, 1999, 23: 603-608[本文引用:1]
[4]
ChenW, ZhangY, LiuX, ChenB, TuJ, FuT. Detection of QTL for six yield-related traits in oilseed rape (Brassica napus) using DH and immortalized F2 populations. Theor Appl Genet, 2007, 115: 849-858[本文引用:2]
[5]
DiepenbrockW. Yield analysis of winter oilseed rape (Brassica napus L. ): a review. Field Crops Res, 2000, 67: 35-49[本文引用:1]
[6]
ShiJ, LiR, QiuD, JiangC, LongY, MorganC, BancroftI, ZhaoJ, MengJ. Unraveling the complex trait of crop yield with quantitative trait loci mapping in Brassica napus. Genetics, 2009, 182: 851-861[本文引用:1]
[7]
Quijada PA, Udall JA, LambertB, Osborn TC. Quantitative trait analysis of seed yield and other complex traits in hybrid spring rapeseed (Brassica napus L. ): 1. Identification of genomic regions from winter germplasm. Theor Appl Genet, 2006, 113: 549-561[本文引用:2]
[8]
吴建忠. 甘蓝型油菜几个农艺性状对产量的影响. 中国科技论文在线精品论文, 2011, 4: 1737-1741Wu JZ. The effects of several agronomic traits on yield in Brassica napus L. Highlights Sciencepaper Online, 2011, 4: 1737-1741 (in Chinese with English abstract)[本文引用:1]
[9]
漆丽萍. 甘蓝型油菜株型与角果相关性状的QTL分析. 华中农业大学博士学位论文, 湖北武汉, 2014. pp 45-73Qi LP. QTL Analysis for the Traits Associated with Plant Architecture and Silique in Brassica napus L. PhD Disseration of Huazhong Agricultural University, Wuhan, China, 2014. pp 45-73 (in Chinese with English abstract)[本文引用:3]
[10]
王道杰, 杨翠玲, 李艳萍, 王再青. 因子分析和数量分类在油菜种质资源遗传多样性研究中的应用. 植物学研究, 2014, 3: 207-217WangD, YangC, LiY, WangZ. Factor analysis and numerical taxonomy applied in the research of germpasm genetic polymorphism in Brassica napus L. Bot Res, 2014, 3: 207-217 (in Chinese with English abstract)[本文引用:1]
[11]
张书芬, 傅廷栋, 朱家成, 王建平, 文雁成, 马朝芝. 甘蓝型油菜产量及其构成因素的QTL定位与分析. 作物学报, 2006, 32: 1135-1142Zhang SF, Fu TD, Zhu JC, Wang JP, Wen YC, Ma CZ. QTL mapping and epistasis analysis for yield and its components in Brassica napus L. Acta Agron Sin, 2006, 32: 1135-1142 (in Chinese with English abstract)[本文引用:1]
[12]
王峰, 官春云. 甘蓝型油菜遗传图谱的构建及单株产量构成因素的QTL分析. 遗传, 2010, 32: 271-277WangF, Guan CY. Molecular mapping and identification of quantitative trait loci foryield components in rapeseed (Brasscia napus L. ). Hereditas (Beijing), 2010, 32(3): 271-277 (in Chinese with English abstract)[本文引用:1]
[13]
ChenW, ZhangY, YaoJ, MaC, TuJ, FuT. Quantitative trait loci mapping for two seed yield component traits in an oilseed rape (Brassica napus) cross. Plant Breed, 2011, 130: 640-646[本文引用:1]
[14]
ChalhoubB, DenoeudF, LiuS, Parkin I A P, Tang H, Wang X, Chiquet J, Belcram H, Tong C, Samans B, Correa M, Da Silva C, Just J, Falentin C, Koh C S, Le Clainche I, Bernard M, Bento P, Noel B, Labadie K, Alberti A, Charles M, Arnaud D, Guo H, Daviaud C, Alamery S, Jabbari K, Zhao M, Edger P P, Chelaifa H, Tack D, Lassalle G, Mestiri I, Schnel N, Le Paslier M C, Fan G, Renault V, Bayer P E, Golicz A A, Manoli S, Lee T H, Thi V H D, Chalabi S, Hu Q, Fan C, Tollenaere R, Lu Y, Battail C, Shen J, Sidebottom C H D, Wang X, Canaguier A, Chauveau A, Berard A, Deniot G, Guan M, Liu Z, Sun F, Lim Y P, Lyons E, Town C D, Bancroft I, Wang X, Meng J, Ma J, Pires J C, King G J, Brunel D, Delourme R, Renard M, Aury J M, Adams K L, Batley J, Snowdon R J, Tost J, Edwards D, Zhou Y, Hua W, Sharpe A G, Paterson A H, Guan C, Wincker P. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science, 2014, 345: 950-953[本文引用:1]
[15]
LiF, ChenB, XuK, GaoG, YanG, QiaoJ, LiJ, LiH, LiL, XiaoX. A genome-wide association study of plant height and primary branch number in rapeseed (Brassica napus). Plant Sci, 2015, DOI: DOI:10.1016/j.plantsci.2015.05.012[本文引用:3]
[16]
LuoX, MaC, YueY, HuK, LiY, DuanZ, WuM, TuJ, ShenJ, YiB, FuT. Unravelling the complex trait of harvest index in rapeseed (Brassica napus L. ) with association mapping. BMC Genom, 2015, 16: 379[本文引用:1]
[17]
LiF, ChenB, XuK, WuJ, SongW, BancroftI, Harper AL, TrickM, LiuS, GaoG, WangN, YanG, QiaoJ, LiJ, LiH, XiaoX, ZhangT, WuX. Genome-wide association study dissects the genetic architecture of seed weight and seed quality in rapeseed (Brassica napus L. ). DNA Res, 2014, 21: 355-367[本文引用:1]
[18]
Hatzig SV, FrischM, BreuerF, NesiN, DucournauS, Wagner MH, Leckband G, AbbadiA, Snowdon RJ. Genome-wide association mapping unravels the genetic control of seed germination and vigor in Brassica napus. Front Plant Sci, 2015, 6: 221[本文引用:2]
[19]
Pritchard JK, StephensM, DonnellyP. Inference of population structure using multilocus genotype data. Genetics, 2000, 155: 945-959[本文引用:1]
[20]
Earl DA, von Holdt BM. STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv Genet Resour, 2012, 4: 359-361[本文引用:1]
[21]
Bradbury PJ, ZhangZ, Kroon DE, Casstevens TM, RamdossY, Buckler ES. TASSEL: software for association mapping of complex traits in diverse samples. Bioinformatics, 2007, 23: 2633-2635[本文引用:1]
[22]
GinestetC. ggplot2: elegant graphics for data analysis. J Roy Stat Soc A, 2011, 174: 245-246[本文引用:1]
[23]
Turner SD. qqman: an R package for visualizing GWAS results using QQ and manhattan plots. BioRxiv, 2014: 005165[本文引用:1]
[24]
Barrett JC, FryB, MallerJ, Daly MJ. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics, 2005, 21: 263-265[本文引用:1]
[25]
杨光. 油菜高效结角层结构的研究, 扬州大学硕士学位论文, 江苏扬州, 2002. p 11YangG. The Study of High-efficient Pod Canopy of Oilseed Rape. MS Thesis of Yangzhou University, Yangzhou, China, 2002. p 11 (in Chinese with English abstract)[本文引用:1]
[26]
浦惠明, 戚存扣, 傅寿仲. 油菜角果的生长特性及其源库效应. 江苏农业科学, 1993, (3): 22-25Pu HM, Qi CK, Fu SZ. Growth characteristic of pod and source-sink response in oilseed. Jiangsu Agric Sci, 1993, (3): 22-25 (in Chinese with English abstract)[本文引用:1]
[27]
冷锁虎, 朱耕如, 邓秀兰. 油菜籽粒干物质来源的研究. 作物学报, 1992, 18: 250-257Leng SH, Zhu GR, Deng XL. Studies on the sources of the dry matter in the seed of rapeseed. Acta Agron Sin, 1992, 18: 250-257 (in Chinese with English abstract)[本文引用:1]
[28]
JeonY, Park YJ, Cho HK, Jung HJ, Ahn TK, KangH, Pai HS. The nucleolar GTPase nucleostemin-like 1 plays a role in plant growth and senescence by modulating ribosome biogenesis. J Exp Bot, 2015: erv337[本文引用:1]
[29]
KomoriR, AmanoY, Ogawa-OhnishiM, MatsubayashiY. Identification of tyrosylprotein sulfotransferase in Arabidopsis. Proc Natl Acad Sci USA, 2009, 106: 15067-15072[本文引用:1]
[30]
ZhongR, Burk DH, Nairn CJ, Wood-JonesA, Morrison WH, Ye ZH. Mutation of SAC1, an Arabidopsis SAC domain phosphoinositide phosphatase, causes alterations in cell morphogenesis, cell wall synthesis, and actin organization. Plant Cell, 2005, 17: 1449-1466[本文引用:1]
 , 王腾岳
, 王腾岳





{kind=link}
{kind=link}
{kind=link}
{kind=link}