关键词:分子标记; 粳稻; 食味品质; 遗传基础 Genetic Analysis and Taste Quality of Main Conventional JaponicaRice Varieties Grown in Jiangsu Province GONG Hong-Bing1,2, ZENG Sheng-Yuan2, LI Chuang2, ZUO Shi-Min1, JING De-Dao2, LIN Tian-Zi2, CHEN Zong-Xiang1, ZHANG Ya-Fang1, QIAN Hua-Fei2, YU Bo2, SHENG Sheng-Lan2, PAN Xue-Biao1,* 1 Jiangsu Key Laboratory of Crop Genetics and Physiology / Co-Innovation Center for Modern Production Technology of Grain Crops, Key Laboratory of Plant Functional Genomics of Ministry of Education, Yangzhou University, Yangzhou 225009, China
2 Zhenjiang Agricultural Research Institute, Jurong 212400, China
Fund:This study was supported by the Natural Science Foundation of Jiangsu Province of China (BK20141291), the Jiangsu Province Self-innovation Program [CX (13)5073], the Key Research and Development Program of Jiangsu Province (Modern Agriculture) (BE2015363), and the Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions AbstractFifty indica-japonicaspecific SSR markers were employed to genotype 86 conventional japonica rice cultivars mainly planted in Jiangsu province and 41 loci showed polymorphism among them. After amplifying in 86 rice varieties, a total of 113 alleles were detected at the 41 SSR loci, and the allele number ranged from two to six, with an average of 2.76 in each locus. Polymorphism information content (PIC) value of these 41 SSR markers ranged from 0.02 to 0.84, with an average value of 0.48; Nei’s expected heterozygosity value of the 41 markers ranged from 0.02 to 0.68, and the average value was 0.37. Based on the coefficient index ranged from 0.72 to 1.00, we divided the 86 cultivars into three groups by UPGMA. The analysis of taste quality traits investigated under different environments indicated that the taste quality of conventional japonica rice cultivars in Jiangsu was correlated with the genetic clustering. Under long-day (LD) condition, the amylose content (AC) and protein content (PC) manifested a tendency of group I > group II > group III, while the taste value (TV) was opposites. And under short-day (SD) condition, taste quality traits of group II and group III varied significantly compared with that under LD condition, while the traits of group I had no significant change. The results indicated that the main cultivars grown in Jiangsu have a general taste quality and narrow genetic background, broadening the genetic basis is an important approach to breed japonica rice cultivars with environment- insensitivity and good eating quality.
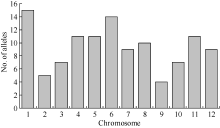
表1 41对有多态的SSR引物的位置以及在86份江淮稻区主栽品种中的遗传多样性信息 Table 1 Location, number of alleles and genetic diversity of 41 SSR loci in 86 Jianghuai-river area major rice varieties
标记座位 Locus
所在位置 Chromosome
等位基因数 na*
有效等位基因数 ne*
Nei’ s指数 Nei’ s* *
多态性信息量 PIC
RM495
1L
3.00
1.92
0.48
0.58
RM583
1L
5.00
2.13
0.53
0.81
RM9
1S
3.00
1.41
0.29
0.66
RM1195
1S
2.00
1.86
0.46
0.46
RM297
1S
2.00
1.10
0.09
0.09
RM525
2L
2.00
2.00
0.50
0.50
RM208
2L
3.00
2.73
0.63
0.70
RM135
3L
3.00
1.12
0.11
0.55
RM22
3L
2.00
1.64
0.39
0.40
RM232
3S
2.00
1.78
0.44
0.44
RM335
4L
4.00
1.97
0.49
0.74
RM255
4S
2.00
1.02
0.02
0.02
RM551
4S
3.00
1.92
0.48
0.63
RM5414
4S
2.00
1.12
0.11
0.11
RM430
5L
6.00
2.37
0.58
0.84
RM169
5S
3.00
1.97
0.49
0.63
RM413
5S
2.00
1.02
0.02
0.02
RM527
6L
3.00
1.98
0.50
0.63
RM340
6L
3.00
1.10
0.09
0.52
RM162
6S
3.00
2.51
0.60
0.68
RM190
6S
2.00
1.49
0.33
0.36
RM253
6S
3.00
1.66
0.40
0.60
RM560
7L
3.00
2.64
0.62
0.69
RM336
7L
4.00
3.14
0.68
0.79
RM18
7L
2.00
2.00
0.50
0.50
RM152
8L
2.00
1.29
0.22
0.22
RM223
8L
3.00
1.88
0.47
0.73
RM72
8S
3.00
2.03
0.51
0.75
RM337
8S
2.00
1.05
0.05
0.05
RM444
9S
2.00
1.93
0.48
0.48
RM219
9S
2.00
1.02
0.02
0.05
RM216
10L
3.00
1.40
0.28
0.62
RM311
10S
4.00
1.61
0.38
0.61
RM332
11L
4.00
2.04
0.51
0.63
RM536
11L
2.00
1.05
0.05
0.05
RM209
11L
2.00
1.10
0.09
0.09
RM224
11S
3.00
1.88
0.47
0.72
RM7003
12L
2.00
1.98
0.50
0.49
RM20a
12L
3.00
2.27
0.56
0.66
RM19
12S
2.00
1.92
0.48
0.48
RM17
12S
2.00
1.20
0.17
0.19
平均Mean
2.76
1.74
0.37
0.48
标准差St. Dev
0.92
0.53
0.20
0.25
L: long arm; S: short arm; na* : observed number of alleles; ne* : effective number of alleles; Nei’ s* * : Nei’ s expected heterozygosity; PIC: polymorphism information content.
表1 41对有多态的SSR引物的位置以及在86份江淮稻区主栽品种中的遗传多样性信息 Table 1 Location, number of alleles and genetic diversity of 41 SSR loci in 86 Jianghuai-river area major rice varieties
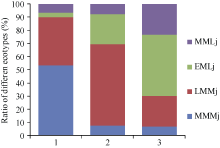
图2 江苏主栽粳稻品种的遗传聚类图 MMMj: 中熟中粳; LMMj: 迟熟中粳; EMLj: 早熟晚粳; MMLj: 中熟晚粳。Fig. 2 Molecular coefficient index and the clustering of main cultivars grown in Jiangsu province MMMj: medium maturing mid-season japonica rice; LMMj: late maturing mid-season japonicarice; EMLj: early maturing late japonica rice; MMLj: medium maturing late japonica rice.
表2 不同遗传类群江苏主栽粳稻品种的食味品质特征 Table 2 Characteristics of taste quality between different genetic groups of japonica rice varieties
类群 Group
2012年江苏 2012 in Jiangsu
2013年江苏 2013 in Jiangsu
2012年海南 2012 in Hainan
AC (%)
PC (%)
TV
AC (%)
PC (%)
TV
AC (%)
PC (%)
TV
I
20.43± 0.97 a
9.60± 1.52 Aa
61.13± 8.26 a
22.26± 1.66 a
11.25± 2.28 a
52.67± 6.83 a
20.71± 1.09 a
8.90± 1.38 a
59.53± 6.69 A
II
20.13± 0.46 a
9.05± 0.72 Aab
62.69± 4.70 a
21.62± 1.10 a
10.40± 1.40 a
53.85± 6.88 a
21.29± 0.88 a
9.63± 1.12 a
55.00± 3.89 B
III
19.96± 0.66 a
8.78± 0.95Ab
64.79± 5.91 a
21.45± 1.84 a
10.18± 2.45 a
56.00± 6.88 a
20.92± 1.10 a
9.13± 1.21 a
57.88± 4.81 AB
AC: amylose content; PC: protein content; TV: taste value. Values followed by different letters are significantly different at 1% (capital) and 5% (lowercase) probability levels, respectively. AC: 直链淀粉含量; PC: 蛋白质含量; TV: 食味分。标以不同字母的值分别在1% (大写字母)和5% (小写字母)水平差异显著。
表2 不同遗传类群江苏主栽粳稻品种的食味品质特征 Table 2 Characteristics of taste quality between different genetic groups of japonica rice varieties
陈温福, 潘文博, 徐正进. 我国粳稻生产现状及发展趋势. 沈阳农业大学学报, 2006, 37: 801-805Chen WF, Pan WB, Xu ZJ. Current situation and trends in production of japonica rice in China. J Shenyang Agric Univ, 2006, 37: 801-805 (in Chinese with English abstract)[本文引用:1]
[2]
王才林, 朱镇, 张亚东, 赵凌. 江苏省粳稻品质改良的成就、问题与对策. 江苏农业学报, 2008, 24: 199-203Wang CL, ZhuZ, Zhang YD, ZhaoL. Achievement and consideration on improving of grain quality for japonica rice in Jiangsu, China. Jiangsu J Agric Sci, 2008, 24: 199-203 (in Chinese with English abstract)[本文引用:1]
[3]
王才林, 张亚东, 朱镇, 陈涛, 赵庆勇, 赵凌, 周丽慧, 姚姝. 优良食味粳稻新品种南粳5055的选育及利用. 农业科技通讯, 2012, (2): 84-88Wang CL, Zhang YD, ZhuZ, ChenT, Zhao QY, ZhaoL, Zhou LH, YaoS. Breeding and application of new good eating quality variety Nanjing5055. Bull Agric Sci & Technol, 2012, (2): 84-88 (in Chinese)[本文引用:1]
[4]
王才林, 陈涛, 张亚东, 朱镇, 赵凌, 林静. 通过分子标记辅助选择培育优良食味水稻新品种. 中国水稻科学, 2009, 23: 25-30Wang CL, ChenT, Zhang YD, ZhuZ, ZhaoL, LinJ. Breeding of a new rice variety with good eating quality by marker assisted selection. Chin J Rice Sci, 2009, 23: 25-30 (in Chinese with English abstract)[本文引用:1]
[5]
潘学彪, 陈宗祥, 左示敏, 张亚芳, 吴旭江, 马宁, 江祺祥, 阙金华, 周春和. 以分子标记辅助选择育成抗条纹叶枯病水稻新品种“武陵粳1号”. 作物学报, 2009, 35: 1851-1857Pan XB, Chen ZX, Zuo SM, Zhang YF, Wu XJ, MaN, Jiang QX, Que JH, Zhou CH. A new rice cultivar “Wulingjing 1” with resistance to rice stripe virus bred by marker assisted selection. Acta Agron Sin, 2009, 35: 1851-1857 (in Chinese with English abstract)[本文引用:1]
[6]
UpadhyayP, Neeraja CN, KoleC, Singh VK. Population structure and genetic diversity in popular rice varieties of India as evidenced from SSR analysis. Biochem Genet, 2012, 50: 770-783[本文引用:1]
[7]
Sun LH, Wang CM, Su CC, Liu YQ, Zhai HQ, Wan JM. Mapping and marker-assisted selection of a brown planthopper resistance gene bph2 in rice (Oryza sativa L. ). J Genet Genomics, 2006, 33: 717-723[本文引用:1]
[8]
McCouch SR, TeytelmanL, XuY, Lobos KB, ClareK, WaltonM, FuB, MaghirangR, LiZ, Xing Y. Development and mappingof2240 new SSR markers for rice (Oryza sativa L. ). DNA Res, 2002, 9: 199-207[本文引用:1]
[9]
McCouch S, Kochert G, Yu Z, Wang Z, Khush G, Coffman W, Tanksley S. Molecular mapping of rice chromosomes. Theor Appl Genet, 1988, 76: 815-829[本文引用:1]
[10]
SheteS, TiwariH, Elston RC. On estimating the heterozygosity and polymorphism information content value. Theor Popul Biol, 2000, 57: 265-271[本文引用:1]
[11]
YehF, YangR, BoyleT. POPGENE, Version 1. 31. Microsoft Window-based Freeware for Population Genetic Analysis. University of Alberta, Edmonton, Canada. In. , 1999[本文引用:1]
[12]
Rohlf FJ. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System. New York: Applied Biostatistics Inc, 1998[本文引用:1]
[13]
徐大勇, 杜永, 方兆伟, 潘启民, 杨建昌, 朱庆森. 江淮稻区不同穗型粳稻品种主要农艺和品质特性的比较分析. 作物学报, 2006, 32: 379-384Xu DY, DuY, Fang ZW, Pan QM, Yang JC, Zhu QS. Comparison on main agronomical and quality characters between japonica cultivars with diferent panicle types in Jiang-Huai area. Acta Agron Sin, 2006, 32: 379-384 (in Chinese with English abstract)[本文引用:1]
[14]
周振玲, 王宝祥, 樊继伟, 卢百关, 赵志刚, 江玲, 秦德荣, 万建民, 徐大勇. 江淮稻区不同生态型粳稻品种的籼粳分化度和遗传多样性. 中国水稻科学, 2012, 26: 431-437Zhou ZL, Wang BX, Fan JW, Lu BG, Zhao ZG, JiangL, Qin DR, Wan JM, Xu DY. Indica-japonica differentiation degree and genetic diversity of japonica cultivars belonging to different ecotypes from the Yangtze-Huaihe region. Chin J Rice Sci, 2012, 26: 431-437 (in Chinese with English abstract)[本文引用:2]
[15]
MartinC, Smith AM. Starch biosynthesis. Plant Cell, 1995, 7: 971[本文引用:1]
[16]
Juliano BO. Criteria and test for rice grain quality. In: Juliano B O, ed. Rice Chemistry and Technology. St. Paul, Minn. American Association of Cereal Chemists Inc. , 1985. pp 443-513[本文引用:1]
[17]
TanY, ZhangQ. Correlation of Simple Sequence Repeat (SSR) variants in the leader sequence of the waxy gene with amylose content of the grain in rice. Acta Bot Sin, 2000, 43: 146-150[本文引用:1]
[18]
万映秀, 邓其明, 王世全, 刘明伟, 周华强, 李平. 水稻Wx基因的遗传多态性及其与主要米质指标的相关性分析. 中国水稻科学, 2006, 20: 603-609Wan YX, Deng QM, Wang SQ, Liu MW, Zhou HQ, Li P. Genetic polymorphism of Wx gene and its correlation with major grain quality traits in rice. Chin J Rice Sci, 2006, 20: 603-609 (in Chinese with English abstract)[本文引用:2]
[19]
张巧凤, 吉健安, 张亚东, 赵凌, 朱镇, 王才林. 粳稻食味仪测定值与食味品尝综合值的相关性分析. 江苏农业学报, 2007, 23: 161-165Zhang QF, Ji JA, Zhang YD, ZhaoL, ZhuZ, Wang CL. Correlation analysis between tested value and comprehensive taste evaluation of japonica rice. Jiangsu J Agric Sci, 2007, 23: 161-165 (in Chinese with English abstract)[本文引用:1]





{kind=link}
{kind=link}
{kind=link}