关键词:花生; AhDGAT2a基因; 启动子; 功能验证 Cloning and Functional Analysis of Peanut AhDGAT2a Promoter ZHENG Ling1,2, SHI Ling-Min1,3, TIAN Hai-Ying1,2, SHAN Lei1,2, BIAN Fei1, GUO Feng1, XUAN Ning1, WAN Shuo-Bo1,2,*, PENG Zhen-Ying1,2,3,* 1 Biotechnology Research Center, Shandong Academy of Agricultural Sciences / Shandong Provincial Key Laboratory of Crop Genetic Improvement, Ecology and Physiology, Jinan 250100, China
2College of Life Science, Shandong University, Jinan 250100, China
3College of Agronomy, Xinjiang Agricultural University, Urumqi 830000, China
Fund:This study was supported by Natural Science Foundation of Shandong (ZR2013CM036) and Shandong Province Germplasm Innovation and Utilization Project (2014-2017) AbstractDiacylglycerol acyltransferase (DGAT) is a rate-limiting enzyme in triacylglycerol (TAG) biosynthesis pathway. In this study, GenomeWalking method was used for cloning the promoter sequence of AhDGAT2a gene from Luhua 14, and finally a 1200 bp fragment flanking 5′-upstream of AhDGAT2a was obtained and named as pAhDGAT2a. The crucial regulatory elements in pAhDGAT2a were further analyzed with software PlantCARE. There were many TATA-box, CAAT-box, light regulation, stress and defense response and hormone response elements. To assess the activity of pAhDGAT2a, we constructed pAhDGAT2a:GUS cassettes and introduced it into the tobacco SR1 genome by Agrobacterium-mediated transformation. Expression pattern was monitored by histochemical staining. Results showed that GUS activity driven by the pAhDGAT2a was detected in almost all vegetative and reproductive tissues, with a higher expression level in stigma, anther and young seeds than in the other organs, indicating that pAhDGAT2a has a constitutive promoter activity.
Keyword: Arachis hypogaeaL .; AhDGAT2a gene; Promoter; Function analysis Show Figures Show Figures
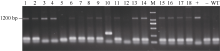
图1 pAhDGAT2a的两轮扩增结果 M: DL2000; A: 第2轮扩增结果; B: 第3轮扩增结果。Fig. 1 PCR products of pAhDGAT2a promoter M: DL2000; A: products of the second round PCR; B: products of the third round PCR.
图3 转基因烟草的GUS染色图 A~D: 野生型烟草; E~H: 转基因烟草; A, E: 烟草幼苗; B, F: 花; C, G: 花药和柱头; D, H: 未成熟的种子。A~C, E~G比例尺为1 cm; D, H比例尺为1 mm。Fig. 3 GUS staining of the transformed tobacco lines A-D, wild type tobacco; E-H, transgenic tobacco. A and E: seedlings; B and F: flowers; C and G: anthers and stigmas; D and H: immature seeds. Bars = 1 cm in A-C and E-G; bars = 1 mm in D, H.
目前关于植物DGAT1基因组织特异性表达的研究有较多报道, 而关于植物DGAT2基因的组织器官特异性表达的研究较少。普遍认为大多数高等植物的DGAT1基因在各组织器官中均表达, 而DGAT2则与植物种子中特殊脂肪酸的积累有关。拟南芥DGAT1在发育中的种子、花瓣、花芽中表达量高, 而在叶和茎中表达量低[19], 大多数双子叶植物例如大豆、斑鸠菊、油菜的表达模式与拟南芥相似[14, 17, 40], 而旱金莲DGAT1只在发育的种子中表达[41]。桐树DGAT2基因在种子发育中期被较强诱导表达, 而在其他器官中表达量很低, 表明桐树DGAT2与种子中脂肪酸积累直接相关[28]。斑鸠菊、大戟、琉璃苣和蓖麻的DGAT2基因在种子发育早期高表达, 而大豆DGAT2基因在种子中的表达量很低。相反, 大豆DGAT1基因在种子中表达量较高[42]。我们前期关于AhDGAT2基因组织器官特异性表达模式研究表明, 该基因在花生的各个器官都表达, 但是在叶、花以及种子发育前期表达量较高[25]。本研究中将pAhDGAT2a构建GUS植物表达载体并转化烟草, 结果发现该序列不仅能正确启动GUS基因表达, 而且在转基因烟草的各个器官中都表达, 与我们前期研究结果一致, 但是与桐树DGAT2和大豆DGAT2的表达特点显著不同。本研究中GUS活性在转基因烟草柱头和花药中较强表达, 显示出AhDGAT2a基因在花生授粉中的重要性, 这一点在其他文献中未见报道。这些研究均表明, DGAT1与DGAT2基因的组织器官特异性表达模式因物种而异。二者不仅在植物种子脂肪酸合成中具有重要作用, 同时对于其他组织器官内的脂肪酸合成(比如维持细胞膜脂类的动态平衡), 甚至对于植物个体的整个发育过程都具有十分重要的作用。 The authors have declared that no competing interests exist.
BrounP, GettnerS, SomervilleC. Genetic engineering of plant lipids. Annu Rev Nutr, 1999, 19: 197-216[本文引用:1]
[2]
JungS, SwiftD, SengokuE, PatelM, TeuléF, PowellG, MooreK, AbbottA. The high oleate trait in the cultivated peanut [Arachis hypogaea L. ]: I. Isolation and characterization of two genes encoding microsomal oleyl-PC desaturases. Mol Gen Genet, 2000, 263: 796-805[本文引用:1]
[3]
Yu YH, Ginsberg HN. The role of acyl-CoA: diacylglycerol acyilransferase (DGAT) in energy metabolism. Ann Med, 2004, 36: 252-261[本文引用:1]
[4]
ZouJ, Wei YD, JakoC, KumarA, SelvarajG, Taylor DC. The Arabidopsis thaliana TAG1 mutant has a mutation in a diacylglycerol acyltransferase gene. Plant J, 1999, 19: 645-653[本文引用:1]
Lu CF, Hills MJ. Arabidopsis mutants deficient in diacylglycerol acyltransferase display increased sensitivity to abscisic acid, sugars and osmotic stress during germination and seedling development. Plant Physiol, 2002, 129: 1352-1358[本文引用:1]
[7]
HeX, Chen GQ, Lin JT, Mckeon TA. Diacylglycerol acyltransferase activity and triacylglycerol synthesis in germinating castor seed cotyledons. Lipids, 2006, 41: 281-285[本文引用:1]
[8]
Weiss SB, Kennedy EP, Kiyasu JY. The enzymatic synthesis of triglycerides. J Biol Chem, 1960, 235: 40-44[本文引用:1]
[9]
SahaS, EnuguttiB, RajakumariS, RajadekharanR. Cytosolic triacylglycerol biosynthetic pathway in oilseeds. Molecular cloning and expression of peanut cytosolic diacylglycerol acyltransferase. Plant Physiol, 2006, 141: 1533-1543[本文引用:4]
HofmannK. A superfamily of membrane-bound O-acyltransferases with implications for WNT signaling. Trends Biochem Sci, 2000, 25: 111-112[本文引用:1]
[12]
CasesS, Smith SJ, Zheng YW. Identification of a gene encoding an acyl CoA: diacylglycerol acyltransferase, a key enzyme in triacylglycerol synthesis. Proc Natl Acad Sci USA, 1998, 95: 13018-13023[本文引用:1]
[13]
BouvierP, BenvenisteP, OelkersP, Sturley SL, SchallerH. Expression in yeast and tobacco of plant cDNAs encoding acyl CoA: diacylglycerol acyltransferase. Eur J Biochem, 2000, 267: 85-96[本文引用:1]
StoneS, LevinM, Farese RV. Membrane topology and identification of key functional amino acid residues of murine acyl-CoA: diacylglycerol acyltransferase-2. J Biol Chem, 2006, 281: 40273-40282[本文引用:1]
[16]
ShockeyJ, Gidda SD, Chapital, KuanJ, Dhanoa P, Bland J, Rothstein S, Mullen R, Dyer J. Tung tree DGAT1 and DGAT2 have nonredundant functions in triacylglycerol biosynthesis and are localized to different subdomains of the endoplasmic reticulum. Plant Cell, 2006, 18: 2294-2313[本文引用:1]
[17]
Hobbs DH, LuC, Hills MJ. Cloning of a cDNA encoding diacylglycerol acyltransferase from Arabidopsis thaliana and its functional expression. FEBS Lett, 1999, 452: 145-149[本文引用:4]
[18]
Lu CL, Noyer SB, Hobbs DH, KangJ, WenY, KratchusD, Hills MJ. Expression pattern of diacyl-glycerol acyltransferase-1, an enzyme involved in triacylglycerol biosynthesis, in Arabidopsis thaliana. Plant Mol Biol, 2003, 52: 31-41[本文引用:2]
[19]
JakoC, KumarA, Wei YD, ZouJ, Barton DL, Giblin EM, Covello PS, Taylor DC. Seed-specific over-expression of an Arabidopsis cDNA encoding a diacylglycerol acyltransferase enhance seed oil content and seed weight. Plant Physiol, 2001, 126: 861-874[本文引用:3]
[20]
ZhengP, AllenW, RoeslerK, WilliamsM, ZhangS, LiJ, GlassmanK, RanchJ, NubelD, SolawetzW, BhattramakkiD, LlacaV, DeschampsS, Zhong GY, Tarczynski, MC, Shen B. A phenylalanine in DGAT is a key determinant of oil content and composition in maize. Nat Genet, 2008, 40: 367-372[本文引用:1]
[21]
LardizabalK, EffertzR, LeveringC, JenniferM, Pedroso MC, TomJ, EricA, KenG, KristenB. Expression of Umbelopsis ramanniana DGAT2A in seed increases oil in soybean. Plant Physiol, 2008, 148: 89-96[本文引用:1]
[22]
BurgalJ, ShockeyJ, LuC, DyerJ, LarsonT, GrahamI, BrowseJ. Metabolic engineering of hydroxy fatty acid production in plants: RcDGAT2 drives dramatic increases in ricinoleate levels in seed oil. Plant Biotechnol J, 2008, 46: 1502-1511[本文引用:1]
[23]
万书波主编. 花生品质学. 北京: 中国农业科学技术出版社, 2005. pp 2-10Wan S B. Peanut Quality. Beijing: China Agricultural Science and Technology Publishers, 2005. pp 2-5(in Chinese)[本文引用:1]
[24]
廖伯寿. 中国花生油脂产业竞争力浅析. 花生学报, 2003, 32: 11-15Liao BS. Analysis the competitiveness of Chinese peanut oil industry. J Peanut Sci, 2003, 32: 11-15 (in Chinese)[本文引用:1]
[25]
王龙龙. 花生二酰甘油酰基转移酶(DGAT)基因的克隆与分析. 山东师范大学硕士学位论文, 山东济南, 2010Wang LL. Cloning and Characterization of Diacylglycerol Acyltransferase (DGAT) Gene in Peanut (Arachis hypogaea L. ). MS Thesis of Shand ong Normal University, Jinan, China, 2010 (in Chinese with English abstract)[本文引用:6]
[26]
Peng ZY, LiL, Yang LQ, ZhangB, ChenG, Bi YP. Overexpression of peanut diacylglycerol acyltransferase2 in Escherichia coli. PLoS One, 2013, 8: e61363[本文引用:2]
[27]
ChiX, HuR, ZhangX, ChenM, ChenN, PanL, WangT, WangM, YangZ, Wang QF, Yu SL. Cloning and functional analysis of three diacylglycerol acyltransferase genes from peanut (Arachis hypogaea L. ). PLoS One, 2014, 9: e105834[本文引用:1]
[28]
Shockey JM, Gidda SK, Chapital DC, Kuan JC, Dhanoa PK, Bland JM, Rothstein SJ, Mullen RT, Dyera JM. Tung tree DGAT1 and DGAT2 have nonredundant functions in triacylglycerol biosynthesis and are localized to different subdomains of the endoplasmic reticulum. Plant Cell, 2006, 18: 2294-2313[本文引用:2]
[29]
Perry HJ, Harwood JL. Changes in the lipid content of developing seeds of Brassica napus. Phytochemistry, 1993, 32: 1411-1415[本文引用:1]
[30]
Lock YY, Snyder CL, ZhuW, Siloto RM, Weselake RJ, ShahS. Antisense suppression of type 1 diacylglycerol acyltransferase adversely affects plant development in Brassica napus. Physiol Plant, 2009, 137: 67-74[本文引用:1]
[31]
Donald RG, Cashmore AR. Mutation of either G box or I box sequences profoundly affects expression from the Arabidopsis rbcS-1A promoter. EMBO J, 1990, 9: 1717-1726[本文引用:2]
[32]
PuenteP, WeiN, Deng XW. Combinatorial interplay of promoter elements constitutes the minimal determinants for light and developmental control of gene expression in Arabidopsis. EMBO J, 1996, 15: 3732-3743[本文引用:1]
[33]
LamE, Chua NH. Gt1 binding site confers light-responsive expression in transgenic tobacco. Science, 1990, 248: 471-474[本文引用:1]
[34]
Chen PW, Lu CA, Yu TS, Teng TH, Wang CS, Yu SM. Rice alpha-amylase transcriptional enhancers direct multiple mode regulation of promoters in transgenic rice. J Biol Chem, 2002, 277: 13641-13649[本文引用:1]
[35]
杨予涛. 一个光合组织特异表达启动子的克隆、功能分析及其转录因子的鉴定. 山东农业大学博士学位论文, 山东泰安, 2005Yang YT. Isolation and Characterization of A Strong Specific Promoter in Photosynthetic Tissues and Identification of a bZIP Transcription Factor. PhD Dissertation of Shand ong Agricultural University, Tai’an, China, 2005 (in Chinese with English abstract)[本文引用:1]
[36]
HoffmannM, BinderS. Functional importance of nucleotide identities within the peat p9 promoter sequence. Mol Biol, 2002, 320: 943-950[本文引用:1]
[37]
YadavV, KunduS, ChattopadhyayD, NegiP, WeiN, Deng XW, ChattopadhyayS. Light regulated modulation of Z-box containing promoters by photoreceptors and downstream regulatory components, COP1 and HY5, in Arabidopsis. Plant J, 2002, 31: 741-753[本文引用:1]
[38]
Tao YB, LuoL, He LL, NiJ, Zeng FX. A promoter analysis of MOTHER OF FT AND TFL1 (JcMFT1), a seed-preferential gene from the biofuel plant Jatropha curcas. Plant Res, 2014, 127: 513-524[本文引用:1]
[39]
XuW, YuY, DingJ, HuaZ, WangL. Characterization of a novel stilbene synthase promoter involved in pathogen- and stress- inducible expression from Chinese wild Vitis pseudoreticulata. Planta, 2010, 231: 475-487[本文引用:1]
[40]
WangH, ZhangJ, GaiJ, ChenS. Cloning and comparative analysis of the gene encoding diacylglycerol acyltransferase from wild type and cultivated soybean. Theor Appl Genet, 2006, 112: 1086-1097[本文引用:1]
[41]
XuJ, FrancisT, MietkiewskaE, Giblin EM, Barton DL, ZhangY, ZhangM, Taylor DC. Cloning and characterization of an acyl-CoA-dependent diacylglycerol acyltransferasel (DGATl) gene from Tropaeolum majus, and a study of the functional motifs of the DGAT protein using site-directed mutagenesis to modify enzyme activity and oil content. Plant Biotechnol J, 2008, 6: 799-818[本文引用:1]
[42]
Li RZ, Yu KS, Hildebrand DF. DGAT1, DGAT2 and PDAT expression in seeds and other tissues of epoxy and hydroxy fatty acid accumulating plants. Lipids, 2010, 45: 145-157[本文引用:1]
 , 彭振英
, 彭振英





{kind=link}
{kind=link}
{kind=link}