关键词:杂交水稻; 种子; 成熟; 小热激蛋白; 寡糖; 活力 Soluble Oligosaccharide and Small Heat Shock Protein Correlated with Seed Germination and Vigor during Hybrid Rice Seed Maturation ZHU Li-Wei1,2,**, CAO Dong-Dong3,**, FU Yu-Ying1, HU Qi-Juan1, LI Zhan1, GUAN Ya-Jing1, HU Wei-Min1, HU Jin1,* 1 Seed Science Center, College of Agriculture and Biotechnology, Zhejiang University, Hangzhou 310058, China
2 Research Center of Buckwheat Industry Technology, Guizhou Normal University, Guiyang 550001, China
3 Zhejiang Agriculture Science Limited Company, Hangzhou 310058, China
Fund:This study was supported by the China Special Fund for Agro-Scientific Research in the Public Interest (201203052), the Subject Fund of Zhejiang Province Science and Technology Hall (2013C02005, 2013C32023), the Natural Sciences Fund of Zhejiang Province (LZ14C130002, LY15C130002), and Jiangsu Collaborative Innovation Center for Modern Crop Production AbstractTo study the relationship between seed soluble oligosaccharide, small heat shock protein (sHSPs) and seed vigor, we systematically studied their changes during hybrid rice seed maturity. The mean fuzzy subordinate function values of germination percentage, germination energy, germination index, vigor index and mean germination time, root number, root length, shoot length and seedling weight were analyzed. The results indicated that seeds harvesting at 19 to 34 days after pollination (DAP) had a higher viability and vigor. The seed moisture content had a decline trend, with a decrease from 48.2% at 10 DAP to 23.9% at 22 DAP. The raffinose content increased significantly, the fructose content kept decreasing, the contents of stachyose and glucose significantly increased firstly and then decreased during seed development. The relative expression levels of 64SHsp18.0 and Os03g0267200 genes increased significantly from 10 to 19 DAP, and then decreased. The moisture content, the contents of raffinose, stachyose and sucrose, thousand seed weight of fresh seeds and dry seeds, relative transcriptions of 64SHsp18.0 and Os03g0267200 genes were all significantly correlated with seed germination during seed maturity.
表1 不同成熟期水稻基因表达量的实时荧光定量PCR所用的引物序列 Table 1 Primers used in q-PCR analysis of the expression of sHsp genes in hybrid rice seeds at different development phases
基因名称Gene name
引物序列Primer sequences(5'-3')
β -OsActin
Forward CTTCATAGGAATGGAAGCTGCGGGTA
Reverse CGACCACCTTGATCTTCATGCTGCTA
64S Hsp18.0
Forward CGTCCTCCACCTCCACCTTG
Reverse GGCAGCATCTTCCCGTCCTT
Os03g0267200
Forward TTCCTCCGCAGGTTCCG
Reverse GGGCTTCTTGGGCTCTT
表1 不同成熟期水稻基因表达量的实时荧光定量PCR所用的引物序列 Table 1 Primers used in q-PCR analysis of the expression of sHsp genes in hybrid rice seeds at different development phases
表2 不同成熟期Y两优689杂交水稻种子的发芽势、发芽率、发芽指数、活力指数和平均发芽时间 Table 2 Germination energy (GE), germination percentage (GP), germination index (GI), vigor index (VI), and mean germination time (MGT) of Y Liangyou 689 hybrid rice seeds at different maturation stages
授粉后天数DAP
发芽势GE (%)
发芽率GP (%)
发芽指数GI
活力指数VI
平均发芽时间MGT (d)
10 d
19.3 c
30.7 c
3.402 f
0.4 f
5.0 a
13 d
70.7 b
83.3 b
10.541 e
2.0 e
4.2 b
16 d
94.7 a
96.0 a
14.973 d
3.2 d
3.3 c
19 d
99.3 a
99.3 a
19.483 a
4.6 a
2.7 e
22 d
99.3 a
99.3 a
17.900 b
4.3 b
2.9 de
25 d
97.3 a
97.3 a
16.794 c
4.0 c
3.1 cd
28 d
100.0 a
100.0 a
19.467 a
4.4 b
2.7 e
31 d
99.3 a
99.3 a
18.217 b
4.1 c
2.8 de
34 d
100.0 a
100.0 a
18.261 b
4.0 c
2.8 de
Values within a column followed by a different small letter are significantly different (α = 0.05, LSD). DAP: days after pollination. 数值后不同字母表示同一列数值间差异显著(α = 0.05, LSD)。
表2 不同成熟期Y两优689杂交水稻种子的发芽势、发芽率、发芽指数、活力指数和平均发芽时间 Table 2 Germination energy (GE), germination percentage (GP), germination index (GI), vigor index (VI), and mean germination time (MGT) of Y Liangyou 689 hybrid rice seeds at different maturation stages
表3 不同成熟期Y两优689杂交水稻种子幼苗的根数、根长、苗高和幼苗鲜重、幼苗干重 Table 3 Root numbers and length, shoot height, seedling fresh, and dry weights of Y Liangyou 689 hybrid rice seedlings at different maturation stages
授粉后天数 DAP
根数 RN
根长 RL (cm)
苗高 SH (cm)
幼苗鲜重 SFW (g 20 seedlings-1)
幼苗干重 SDW (g 20 seedlings-1)
10 d
6.0 d
5.5 b
7.3 f
0.714 e
0.133 e
13 d
7.5 c
5.5 b
6.4 g
1.134 d
0.191 d
16 d
9.1 b
4.6 d
8.0 e
1.272 cd
0.210 cd
19 d
10.2 a
5.3 bc
9.8 bc
1.511 ab
0.235 ab
22 d
10.5 a
5.2 bc
10.7 a
1.372 abc
0.241 a
25 d
10.0 a
5.1 c
8.7 d
1.548 a
0.241 a
28 d
10.1 a
4.8 d
10.0 b
1.352 bc
0.223 abc
31 d
10.5 a
6.2 a
10.5 a
1.423 abc
0.225 abc
34 d
9.3 b
6.3 a
9.4 c
1.428 abc
0.217 bc
Values within a column followed by a different small letter are significantly different (α = 0.05, LSD). RN: root numbers; RL: root length; SH: shoot height; SFW: seedling fresh weights; SDW: seedling dry weights; DAP: days after pollination. 数值后不同字母表示同一列数值间差异显著性(α = 0.05, LSD)。
表3 不同成熟期Y两优689杂交水稻种子幼苗的根数、根长、苗高和幼苗鲜重、幼苗干重 Table 3 Root numbers and length, shoot height, seedling fresh, and dry weights of Y Liangyou 689 hybrid rice seedlings at different maturation stages
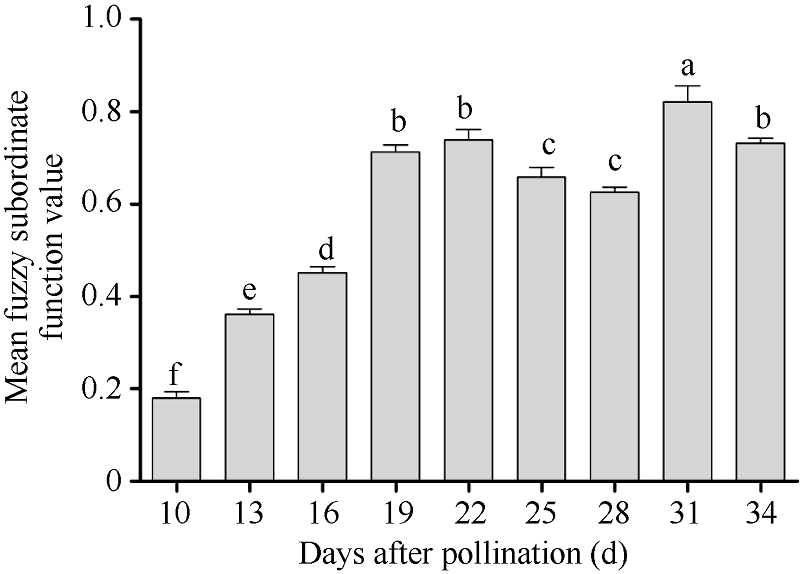
表4 不同成熟期Y两优689杂交水稻种子的发芽势、发芽率、发芽指数、活力指数和平均发芽时间的模糊隶属函数值 Table 4 Mean fuzzy subordinate function value of germination energy, germination percentage, germination index, vigor index, and mean germination time of Y Liangyou 689 rice seeds at different maturation stages
授粉后天数DAP
FGE
FGP
FGI
FVI
FMGT
平均函数A-value
10 d
0.000
0.000
0.000
0.000
0.000
0.000 e
13 d
0.637
0.759
0.444
0.381
0.348
0.514 d
16 d
0.934
0.942
0.720
0.667
0.739
0.800 c
19 d
0.991
0.990
1.000
1.000
1.000
0.996 a
22 d
0.991
0.990
0.902
0.929
0.913
0.945 ab
25 d
0.967
0.961
0.833
0.857
0.826
0.889 b
28 d
1.000
1.000
0.999
0.952
1.000
0.990 ab
31 d
0.991
0.990
0.921
0.881
0.957
0.948 ab
34 d
1.000
1.000
0.924
0.857
0.957
0.948 ab
Values within a column followed by a different small letter are significantly different (α = 0.05, LSD). FGE: fuzzy subordinate function value of germination energy; FGP: fuzzy subordinate function value of germination percentage; FGI: fuzzy subordinate function value of germination index; FVI: fuzzy subordinate function value of vigor index; FMGT: fuzzy subordinate function value of mean germination time; A-value: mean fuzzy subordinate function value of evaluation index. DAP: days after pollination. 数值后不同字母表示同一列数值间差异显著(α = 0.05, LSD)。FGE: 发芽势的模糊隶属函数; FGP: 发芽率的模糊隶属函数; FGI: 发芽指数的模糊隶属函数; FVI: 活力指数的模糊隶属函数; FMGT: 平均发芽时间的模糊隶属函数; A-value: 所有指标的平均模糊隶属函数。
表4 不同成熟期Y两优689杂交水稻种子的发芽势、发芽率、发芽指数、活力指数和平均发芽时间的模糊隶属函数值 Table 4 Mean fuzzy subordinate function value of germination energy, germination percentage, germination index, vigor index, and mean germination time of Y Liangyou 689 rice seeds at different maturation stages
表5 不同成熟期Y两优689杂交水稻种子幼苗的根数、根长、苗高和苗鲜重、苗干重的模糊隶属函数值 Table 5 Mean fuzzy subordinate function values of root number and length, shoot height, fresh and dry weights of seedling in Y Liangyou 689 hybrid rice at different maturation stages
授粉后天数DAP
FRN
FRL
FSH
FSFW
FSDW
平均函数A-value
10 d
0.000
0.529
0.209
0.000
0.000
0.148 f
13 d
0.333
0.529
0.000
0.504
0.537
0.381 e
16 d
0.689
0.000
0.372
0.669
0.713
0.489 d
19 d
0.933
0.412
0.791
0.956
0.944
0.807 b
22 d
1.000
0.353
1.000
0.789
1.000
0.828 b
25 d
0.889
0.294
0.535
1.000
1.000
0.744 c
28 d
0.911
0.118
0.837
0.765
0.833
0.693 c
31 d
1.000
0.941
0.953
0.850
0.852
0.919 a
34 d
0.733
1.000
0.698
0.856
0.778
0.813 b
Values within a column followed by a different small letter are significantly different (α = 0.05, LSD). FRN: fuzzy subordinate function value of root number; FRL: fuzzy subordinate function value of root length; FSH: fuzzy subordinate function value of shoot height; FSFW: fuzzy subordinate function value of fresh weights of seedling; FSDW: fuzzy subordinate function value of dry weights of seedling. A-value: mean fuzzy subordinate function value of evaluation index. DAP: days after pollination. 数值后不同字母表示同一列数值间差异显著性(α = 0.05, LSD)。FRN: 根数的模糊隶属函数; FRL: 根长的模糊隶属函数; FSH: 苗高的模糊隶属函数; FSFW: 苗鲜重的模糊隶属函数; FSDW: 苗干重的模糊隶属函数; A-value: 所有指标的平均模糊隶属函数。
表5 不同成熟期Y两优689杂交水稻种子幼苗的根数、根长、苗高和苗鲜重、苗干重的模糊隶属函数值 Table 5 Mean fuzzy subordinate function values of root number and length, shoot height, fresh and dry weights of seedling in Y Liangyou 689 hybrid rice at different maturation stages
图1 不同成熟期Y两优689杂交水稻种子发芽与幼苗质量的平均模糊隶属函数值柱状图上方不同的字母表示不同成熟期种子间的差异显著(α = 0.05, LSD)。Fig. 1 Mean fuzzy subordinate function value in seed germination and seedling quality at different maturation stages of Y Liangyou 689 hybrid riceDifferent small letters on top of bars indicate the significant differences of mean fuzzy subordinate function values of seeds at different maturation stages (α = 0.05, LSD).
表6 不同成熟期Y两优689杂交水稻种子含水量及其鲜重和干重 Table 6 Differences of moisture content, thousand-seed weight of fresh seeds and dry seeds at different maturation stages of Y Liangyou 689 hybrid rice seeds
授粉后天数 DAP
天气 Weather
相对湿度 RH (%)
含水量 MC (%)
鲜种子千粒重 TFSW (g)
干种子千粒重 TDSW (g)
10 d
阴 Cloudy
92
48.2 a
20.568 c
11.640 d
13 d
阴 Cloudy
93
34.1 b
22.828 b
15.960 c
16 d
阴 Cloudy
91
26.0 cd
24.170 a
19.047 b
19 d
雨 Rain
97
27.9 c
24.091 a
19.047 b
22 d
阴 Cloudy
90
23.9 d
23.951 ab
19.386 ab
25 d
阴 Cloudy
94
23.9 d
24.166 a
19.533 a
28 d
雨 Rain
96
26.4 cd
24.170 a
19.540 a
Different small letters after the number indicate the significant difference (α = 0.05, LSD) within the same column. RH: relative humidity; MC: moisture content; TFSW: thousand-seed weight of fresh seeds; TDSW: thousand seed weight of dry weight. DAP: days after pollination. 数值后不同字母表示同一列数值间差异显著(α = 0.05, LSD)。
表6 不同成熟期Y两优689杂交水稻种子含水量及其鲜重和干重 Table 6 Differences of moisture content, thousand-seed weight of fresh seeds and dry seeds at different maturation stages of Y Liangyou 689 hybrid rice seeds
表7 不同成熟期Y两优689杂交水稻种子中水苏糖、棉子糖、蔗糖、葡萄糖和果糖的含量 Table 7 Soluble stachyose, raffinose, sucrose, glucose, and fructose contents at different maturation stages of Y Liangyou 689 hybrid rice (µ g 10-seed-1)
授粉后天数 DAP
水苏糖 Stachyose
棉子糖 Raffinose
蔗糖 Sucrose
葡萄糖 Glucose
果糖 Fructose
10 d
52.969 c
5.110 f
42.045 f
4.814 c
1.103 b
13 d
52.565 c
15.495 e
69.859 e
7.868 a
1.200 b
16 d
101.208 b
16.136 e
117.949 d
5.124 c
0.871 c
19 d
184.617 a
21.252 d
191.934 a
6.022 b
1.693 a
22 d
187.552 a
28.031 c
148.509 b
3.106 d
1.732 a
25 d
27.187 d
35.816 b
142.333 c
2.603 e
1.067 c
28 d
31.117 d
43.004 a
180.374 a
1.549 f
0.869 c
Values within the same column followed by different letters are significantly different (α = 0.05, LSD). DAP: days after pollination. 数值后不同字母表示同一列数值间差异显著(α = 0.05, LSD)。
表7 不同成熟期Y两优689杂交水稻种子中水苏糖、棉子糖、蔗糖、葡萄糖和果糖的含量 Table 7 Soluble stachyose, raffinose, sucrose, glucose, and fructose contents at different maturation stages of Y Liangyou 689 hybrid rice (µ g 10-seed-1)
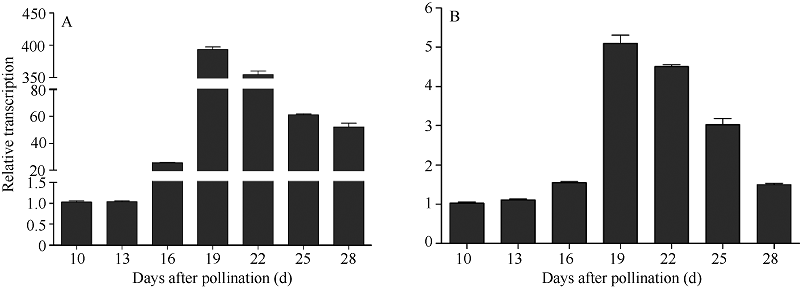
图2 不同成熟期Y两优689水稻种子64S Hsp18.0 (A)和Os03g0267200 (B)基因相对表达量Fig. 2 Relative expression levels of 64S Hsp18.0 (A) and Os03g0267200 (B) genes at different maturation stages of Y Liangyou 689 hybrid rice seed
表8 Y两优689杂交水稻种子成熟过程生理生化的变化与种子发芽率和发芽势之间的直线回归关系(授粉后10~22 d) Table 8 Linear regression relationships of the physiological and biochemical changes with germination percentage and germination energy at different maturation stages in Y Liangyou 689 hybrid rice seeds (10 to 22 DAP)
生理指标 Physiological parameter
发芽率GP
发芽势GE
线性回归方程 Linear regression equation
皮尔逊相关系数 Pearson correlation coefficient
线性回归方程 Linear regression equation
皮尔逊相关系数 Pearson correlation coefficient
含水量MC
y = -2.322x + 147.2
-0.947*
y = -3.029x + 147.7
-0.834
鲜种子千粒重TFSW
y = 14.510x - 261.3
0.894*
y = 17.860x - 360.9
0.743
干种子千粒重TDSW
y = 7.060x - 46.47
0.947*
y = 9.074x - 102.6
0.821
棉子糖含量Raffinose content
y = 0.321x + 35.72
0.897*
y = 0.499x - 7.057
0.941*
水苏糖含量Stachyose content
y = 2.738x + 25.81
0.957* *
y = 4.099x - 19.72
0.968* *
蔗糖含量Sucrose content
y = 0.369x + 30.74
0.923*
y = 0.534x - 10.15
0.889*
葡萄糖含量Glucose content
y = -3.670x + 92.70
-0.266
y = -6.595x + 86.33
-0.322
果糖含量Fructose content
y = 41.880x + 17.65
0.658
y = 79.050x - 53.54
0.838
64S Hsp18. relative express level
y = 0.096x + 57.77
0.816
y = 0.161x + 25.41
0.922*
Os03g0267200 relative express level
y = 9.897x + 46.90
0.826
y = 16.270x + 7.993
0.917*
* and * * indicate significant correlation at the 0.05 and 0.01 probability levels, respectively. Abbreviations correspond with those given in Table 2 and Table 6. 数值后的* 和* * 分别表示α =0.05和α =0.01水平上显著相关。缩写同表2和表6。
表8 Y两优689杂交水稻种子成熟过程生理生化的变化与种子发芽率和发芽势之间的直线回归关系(授粉后10~22 d) Table 8 Linear regression relationships of the physiological and biochemical changes with germination percentage and germination energy at different maturation stages in Y Liangyou 689 hybrid rice seeds (10 to 22 DAP)
表9 Y两优689杂交水稻种子成熟过程中生理生化变化与种子发芽指数和活力指数间的直线回归关系(授粉后10~22 d) Table 9 Linear regression relationships of the physiological and biochemical changes with germination index and vigor index at different maturation stages in Y Lianyou 689 hybrid rice seeds (10 to 22 DAP)
生理指标 Physiological parameter
发芽指数GI
活力指数VI
线性回归方程 Linear regression equation
皮尔逊相关系数 Pearson correlation coefficient
线性回归方程 Linear regression equation
皮尔逊相关系数 Pearson correlation coefficient
含水量MC
y = -0.443x + 21.88
0.801
y = -0.085x + 4.030
-0.825
鲜种子千粒重TFSW
y = 2.628x - 52.89
0.717
y = 0.476x - 9.689
0.697
干种子千粒重TDSW
y = 1.339x - 14.98
0.795
y = 0.250x - 2.929
0.794
棉子糖含量Raffinose content
y = 0.076x - 1.229
0.950*
y = 0.014x - 0.362
0.948*
水苏糖含量Stachyose content
y = 0.613x - 2.880
0.951*
y = 0.116x - 0.711
0.969* *
蔗糖含量Sucrose content
y = 0.082x - 1.700
0.907*
y = 0.014x - 0.359
0.859
葡萄糖含量Glucose content
y = -0.984x + 12.97
-0.316
y = -0.277x + 2.790
-0.476
果糖含量Fructose content
y = 12.440x - 8.747
0.865
y = 2.183x - 1.583
0.813
64S Hsp18.0 relative express level
y = 0.025x + 3.713
0.943*
y = 0.004x + 0.584
0.911*
Os03g0267200 relative express level
y = 2.541x + 0.987
0.939*
y = 0.451x + 0.110
0.894*
* and * * indicate significant correlation at the 0.05 and 0.01 probability levels, respectively. Abbreviations correspond with those given in Table 2 and Table 6. 数值后的* 和* * 分别表示α =0.05和α =0.01 水平上显著相关。缩写同表2和表6。
表9 Y两优689杂交水稻种子成熟过程中生理生化变化与种子发芽指数和活力指数间的直线回归关系(授粉后10~22 d) Table 9 Linear regression relationships of the physiological and biochemical changes with germination index and vigor index at different maturation stages in Y Lianyou 689 hybrid rice seeds (10 to 22 DAP)
表10 Y两优689杂交水稻种子成熟过程中生理生化变化与种子平均发芽时间(MGT)的直线回归关系(授粉后10~22 d) Table 10 Linear regression relationships of the physiological and biochemical changes with mean germination time at different maturation stages in Y Liangyou 689 hybrid rice seeds (10 to 22 DAP)
生理指标 Physiological parameters
线性回归方程 Linear regression equation
皮尔逊相关系数 Pearson correlation coefficient
含水量MC
y = 0.203x - 1.947
0.984* *
鲜种子千粒重TFSW
y = -1.362x + 35.97
-0.993* *
干种子千粒重TDSW
y = -0.627x + 15.19
-0.995* *
棉子糖含量Raffinose content
y = -0.006x + 4.648
-0.681
水苏糖含量Stachyose content
y = -0.200x + 8.021
-0.829
蔗糖含量Sucrose content
y = -0.027x + 7.709
-0.810
葡萄糖含量Glucose content
y = 0.077x + 4.164
0.066
果糖含量Fructose content
y = -1.750x + 6.890
-0.325
64S Hsp18.0 relative express level
y = -0.005x + 5.424
-0.538
Os03g0267200 relative express level
y = -0.574x + 6.091
-0.568
* and * * indicate significant correlation at the 0.05 and 0.01 probability levels, respectively. Abbreviations correspond with those given in Table 2 and Table 6. 数值后的* 和* * 分别表示α = 0.05和α = 0.01水平上显著相关。缩写同表2和表6。
表10 Y两优689杂交水稻种子成熟过程中生理生化变化与种子平均发芽时间(MGT)的直线回归关系(授粉后10~22 d) Table 10 Linear regression relationships of the physiological and biochemical changes with mean germination time at different maturation stages in Y Liangyou 689 hybrid rice seeds (10 to 22 DAP)
王仁祥, 曹文亮, 肖层林, 谢建华. 株1 S杂交组合制种不同收获期种子贮藏特性研究. 种子, 2008, 27(12): 101-104Wang RX, Cao WL, Xiao CL, Xie JH. Study on seed storage in different harvesting dates of Zhu 1 S hybrid rice combinations. Seed, 2008, 27(12): 101-104 (in Chinese)[本文引用:1]
[2]
周建明, 林一波, 何建华, 沈雪林, 戴华军, 吴锡清. 不同收获期和贮藏时间对杂交粳稻种子活力的影响. 种子, 2010, 29(10): 98-101Zhou JM, Lin YB, He JH, Shen XL, Dai HJ, Wu XQ. Effects of different harvest date and storage time on seed vigor of hybrid japonica. Seed, 2010, 29(10): 98-101 (in Chinese)[本文引用:2]
[3]
Fussell LK, Pearson CJ. Effects of grain development and thermal history on grain maturation and seed vigour of Pennisetuma mericanum. J Exp Bot, 1980, 31: 635-643[本文引用:1]
[4]
Cao DD, HuJ, Huang XX, Wang XJ, Guan YJ, Wang ZF. Relationships between changes of kernel nutritive components and seed vigor during development stages of F1 seeds of sh2 sweet corn. J Zhejiang Univ-Sci B, 2008, 9: 964-968[本文引用:2]
[5]
孟庆立, 关周博, 冯佰利, 柴岩, 胡银岗. 谷子抗旱相关性状的主成分与模糊聚类分析. 中国农业科学, 2009, 42: 2667-2675Meng QL, Guan ZB, Feng BL, ChaiY, Hu YG. Principal component analysis and fuzzy clustering on drought-tolerance related traits of foxtail millet (Setaria italica). Sci Agric Sin, 2009, 42: 2667-2675 (in Chinese with English abstract)[本文引用:2]
[6]
黄先晖, 杨远柱, 姜孝成. 水稻种子脱水耐性的形成及其与贮藏特性的关系. 种子, 2010, 29(7): 25-29Huang XH, Yang YZ, Jiang XC. Studies on the formation of desiccation tolerance and the correlation with storage of Oryza sativa L. seeds. Seed, 2010, 29(7): 25-29 (in Chinese with English abstract)[本文引用:2]
[7]
Harrington JF. Seed storage and longevity. In: Kozlowski T T, ed. Seed Biology. New York: Academic Press, 1972. pp 145-245[本文引用:1]
[8]
Eskand ariH. Seed quality changes in cowpea (Vigna sinensis) during seed development and maturation. Seed Sci Technol, 2012, 40: 108-112[本文引用:1]
[9]
DemirI, SamitY. Seed quality in relation to fruit maturation and seed dry weight during development in tomato. Seed Sci Technol, 2001, 29: 453-462[本文引用:1]
[10]
DemirI, MaviK, OztokatC. Changes in germination and potential longevity of watermelon (Citrullus lanatus) seeds during development. New Zeal J Crop Hortic, 2004, 32: 139-145[本文引用:1]
[11]
Blackman SA, Obendorf RL, Leopold AC. Maturation proteins and sugars in desiccation tolerance of developing soybean seeds. Plant Physiol, 1992, 100: 225-230[本文引用:1]
[12]
Gorecki RJ, Lahuta LB, Jones AD, Hedley CL. Soluble sugar in maturing pea seeds of different lines in relation to desiccation tolerance. In: Black M, Bradford K J, Vazquezramos J, eds. Seed Biology: Advances and Applications. Proceedings of the Sixth International Workshop on Seeds. Merida, Mexico, 1999. CABI, 2000. pp 67-74[本文引用:1]
[13]
BaudS, Boutin JP, MiquelM, LepiniecL, RochatC. An integrated overview of seed development in Arabidopsis thaliana ecotype WS. Plant Physiol Biochem, 2002, 40: 151-160[本文引用:1]
[14]
WehmeyerN, VierlingE. The expression of small heat shock proteins in seeds responds to discrete developmental signals and suggests a general protective role in desiccation tolerance. Plant Physiol, 2000, 122: 1099-1108[本文引用:1]
[15]
Neta-sharirI, IsaacsonT, LurieS, WeissD. Dual role for tomato heat shock protein 21: protecting photosystem II from oxidative stress and promoting color changes during fruit maturation. Plant Cell, 2005, 17: 1829-1838[本文引用:1]
[16]
Sarkar NK, Kim YK, GroverA. Rice sHsp genes: genomic organization and expression profiling under stress and development. BMC Genomics, 2009, 10: 393[本文引用:3]
[17]
ZouJ, Liu CF, Liu AL, ZouD, Chen XB. Overexpression of OsHsp17. 0 and OsHsp23. 7 enhances drought and salt tolerance in rice. J Plant Physiol, 2012, 169: 628-635[本文引用:1]
[18]
ChauhanH, KhuranaN, AgarwalP, KhuranaP. Heat shock factors in rice (Oryza sativa L. ): genome-wide expression analysis during reproductive development and abiotic stress. Mol Genet Genomics, 2011, 286: 171-187[本文引用:1]
[19]
Tian YX, LiZ, HeF, Guan YJ, Zhu SJ, HuJ. A novel anti-counterfeiting methods: application and decomposition of RB for broad bean seeds (Vicia faba L. ). Ind Crop Prod, 2014, 61: 278-283[本文引用:2]
[20]
BaillyC, AudigierC, LadonneF, LadonneF, Wagner MH, CosteF, CorbineauF, CômeD. Changes in oligosaccharide content and antioxidant enzyme activities in developing bean seeds as related to acquisition of drying tolerance and seed quality. J Exp Bot, 2001, 52: 701-708[本文引用:1]
[21]
Zheng Y Y. Hu J, Xu SC, Guan YJ, Wang XJ. Relationship between polyamine changes in embryos of maize and seed imbibitional chilling tolerance. Seed Sci Technol, 2009, 37: 59-69[本文引用:1]
肖层林, 刘爱民, 张海清, 肖晓, 余雪晴. 中国杂交水稻制种技术的进步与发展方向. 杂交水稻, 2010, (增刊1): 46-50Xiao CL, Liu AM, Zhang HQ, XiaoX, Yu XQ. Progress and developing direction of hybrid rice seed production technology in china. Hybrid Rice, 2010, (suppl-1): 46-50 (in Chinese)[本文引用:1]
[24]
王业文, 张莹, 靳西彪, 魏毅, 朱建清. 抗穗萌药剂组合的筛选及其抑制种子发芽的同功酶电泳研究. 种子, 2008, 27(8): 18-21Wang YW, ZhangY, Jin XB, WeiY, Zhu JQ. The selection of inhibitor combination to inhibit pre-harvest sprouting in rice and study on eletrophoretic analysis. Seed, 2008, 27(8): 18-21 (in Chinese with English abstract)[本文引用:1]
[25]
朱丽伟. 杂交水稻种子成熟过程活力、生理生化和耐藏力的变化及脱水剂应用效果的研究. 浙江大学博士学位论文, 2015Zhu LW. Changes of Vigor, Physiology, Biochemistry and Storability during Hybrid Rice Seed Maturity and the Effect of Dehydrating Agent Application. PhD Dissertation of Zhejiang University, Hangzhou, China, 2015 (in Chinese with English abstract)[本文引用:1]
[26]
樊廷录, 王淑英, 王建华, 杨珍. 河西制种基地玉米杂交种种子成熟期与种子活力的关系. 中国农业科学, 2014, 47: 2960-2970Fan TL, Wang SY, Wang JH, YangZ. Relationship of days after pollination and vigor traits on maize seed maturity in Hexi seed production area in China. Sci Agric Sin, 2014, 47: 2960-2970 (in Chinese with English abstract)[本文引用:1]
[27]
Steadman KJ, Pritchard HW, Dey PM. Tissue-specific soluble sugars in seeds as indicators of storage category. Ann Bot-London, 1996, 77: 667-674[本文引用:1]
[28]
Obendorf RL. Oligosaccharides and galactosyls in seed desiccation tolerance. Seed Sci Res, 1997, 7: 63-74[本文引用:1]
[29]
HeimU, WeberH, BaumleinH, WobusU. A sucrose synthase gene of Vicia faba L. expression pattern in developing seeds in relation to starch synthesis and metabolic regulation. Planta, 1993, 191: 394-401[本文引用:1]
[30]
AbidG, MuhovskiY, Jacquemin JM, MingeotD, SassiK, ToussaintA, BaudoinJ. Characterization and expression profile analysis of a sucrose synthase gene from common bean (Phaseolus vulgaris L. ) during seed development. Mol Biol Rep, 2012, 39: 1133-1143[本文引用:1]
[31]
Ruan YL, Liewellyn DJ, LiuQ, Xu SM, Wu LM, WangL, Furbank RT. Expression of sucrose synthase in the developing endosperm is essential for early seed development in cotton. Funct Plant Biol, 2008, 35: 382-393[本文引用:1]
[32]
Angeles-NúñezJ G, TiessenA. Mutation of the transcription factor LEAFY COTYLEDON 2 alters the chemical composition of Arabidopsis seeds, decreasing oil and protein content, while maintaining high levels of starch and sucrose in mature seeds. J Plant Physiol, 2011, 168: 1891-1900[本文引用:1]
[33]
陶月良, 朱诚. 顽拗性板栗种子成熟前后褐变与可溶性糖的关系. 农业工程学报, 2003, 19(4): 201-204Tao YL, ZhuC. Relationship between browning and soluble sugar of recalcitrant seeds of chestnut in front of and behind maturation. Trans CSAE, 2003, 19(4): 201-204 (in Chinese with English abstract)[本文引用:1]
[34]
Obendorf RL, Zimmerman AD, Zhang QY, CastilloA, Kosina SM, Bryant EG, Sensenig EM, WuJ, Schnebly SR. Accumulation of soluble carbohydrates during seed development and maturation of low-raffinose, low-stachyose soybean. Crop Sci, 2009, 49: 329-341[本文引用:1]
[35]
李永红, 马颖敏, 许柏球, 赵梁军. 巴拿马栗种子发育中可溶性糖变化与种子脱水耐性的关系. 中国农业科学, 2009, 42: 2882-2891Li YH, Ma YM, Xu BQ, Zhao LJ. Relationship between dehydration tolerance and soluble sugars during seed development of Pachira macrocarpa. Sci Agric Sin, 2009, 42: 2882-2891 (in Chinese with English abstract)[本文引用:1]
[36]
Sheila AB, Ralph LO, Leopold AC. Maturation proteins and sugars in desiccation tolerance of developing soybean seeds. Plant Physiol, 1992, 100: 225-230[本文引用:1]
[37]
LehnerA, BaillyC, FlechelB, PoelsP, CômeD, CorbineauF. Changes in wheat seed germination ability, soluble carbohydrate and antioxidant enzymes activities in the embryo during the desiccation phase of maturation. J Cereal Sci, 2006, 43: 175-182[本文引用:1]
[38]
Koster KL, Leopold AC. Sugars and desiccation tolerance in seeds. Plant Physiol, 1988, 88: 829-832[本文引用:1]
Lin SK, Chang MC, Tsai YG, Lur HS. Proteomic analysis of the expression of proteins related to rice quality during caryopsis development and the effect of high temperature on expression. Proteomics, 2005, 5: 2140-2156[本文引用:1]
[41]
郭鹏, 隋娜, 于超, 郭尚敬, 董新纯, 孟庆伟. 转入甜椒热激蛋白基因CaHSP18提高番茄的耐冷性. 植物生理学通讯, 2008, 44: 409-412GuoP, SuiN, YuC, Guo SJ, Dong XC, Meng QW. CaHSP18. 0 of sweet pepper enhanced chilling tolerance of transgenic tomato plants. Plant Physiol Commun, 2008, 44: 409-412 (in Chinese with English abstract)[本文引用:1]
 , 曹栋栋
, 曹栋栋


{kind=link}
{kind=link}