关键词:甘蔗; SOS1基因; 电子克隆; 生物信息学; 实时荧光定量PCR Isolation and Characterization of a Na+/H+ Antiporter Gene from Sugarcane LIU Feng**, SU Wei-Hua**, HUANG Long, XIAO Xin-Huan, HUANG Ning, LING Hui, SU Ya-Chun, ZHANG Hua, QUE You-Xiong* Key Laboratory of Sugarcane Biology and Genetic Breeding (Fujian), Ministry of Agriculture, Fujian Agriculture and Forestry University / Sugarcane Research & Development Center, China Agricultural Technology System, Fuzhou 350002, China Fund:This study was supported by the China Agriculture Research System (CARS-20), the Special Fund for Agro-Scientific Research in the Public Interest (201503119), and the Program for New Century Excellent Talents in Fujian Province University (JA14095) AbstractSalt overly sensitive 1 ( SOS1) gene, encoding a Na+/H+ antiport protein, plays an important role in biological processes of plants against salt stress. Using a SOS1mRNA sequence from Triticum aestivum (KJ563230) as the probe, the homologous ESTs of sugarcane were obtained from NCBI database. A sugarcane cDNA sequence of SOS1 gene was cloned by in silico cloning combined with RT-PCR, and named as ScSOS1 (GenBank accession number: KT003285). The bioinformatics analysis showed that ScSOS1has a length of 1403 bp with a complete open reading frame (ORF, 107 to 1423 bp), encoding a 423 amino acid residues of sugarcane SOS1 protein with an estimated molecular weight of 47.6 kD and a calculated isoelectric point (pI) of 9.12. The protein of ScSOS1 belongs to a conserved CAP-ED superfamily. Yet the ScSOS1 protein has no signal peptide and belongs to hydrophilic protein with the main function for intermediary metabolism. The mainly secondary structure element of ScSOS1 protein is random coil. Real-time quantitative PCR (RT-qPCR) analysis revealed that ScSOS1 was tissue-specifically expressed in leaf sheath, bark, pulp, bud and root of sugarcane, with the highest expression in leaf sheath and the lowest in root. Besides, the expression of ScSOS1gene could be regulated by the treatments of NaCl, PEG, ABA, SA, and MeJA, and up-regulated by the stresses of NaCl and PEG, with the highest inducible expression levels of 1.5 times and 4.0 times as high as those of control at 24 hours, respectively. This paper suggested that ScSOS1 involves in sugarcane tolerance salt and osmotic stresses. It can set up a basis for the elucidation of sugarcane salt resistance mechanism.
Keyword:Sugarcane; SOS1gene; in silico cloning; Bioinformatics; Real-time quantitative PCR Show Figures Show Figures
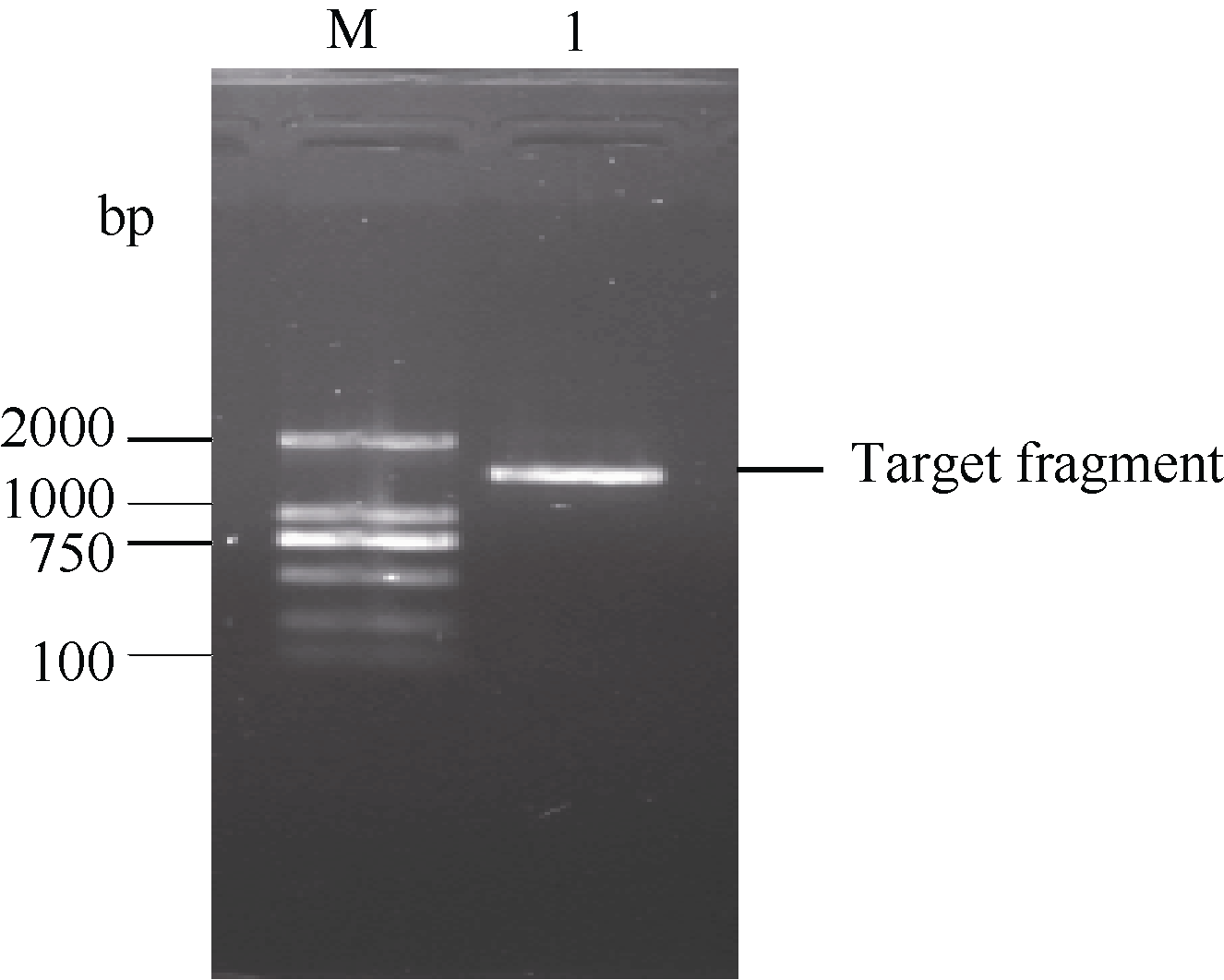
图2 甘蔗ScSOS1基因的RT-PCR扩增验证所获得的序列及其推导的氨基酸序列(* : 终止密码子) 黑色框部分为特异性引物在基因序列中的位置。Fig. 2 Nucleotide acid sequence and deduced amino acid sequence of sugarcane ScSOS1 gene obtained by cloning (* stop codon) The sequence fragment complementary to primer is highlighted in black box.
表3 Table 3 表3(Table 3)
表3 甘蔗ScSOS1蛋白二级结构预测分析 Table 3 Secondary structure prediction of sugarcane ScSOS1 protein
二级结构类型 Secondary structure type
氨基酸残基数目(个) Amino acid residue number (a)
百分比 Percentage (%)
α -螺旋 Alpha-helix
139
32.86
延伸链 Extended strand
67
15.84
无规则卷曲 Random coil
217
51.30
β -螺旋 Beta-helix
0
0
表3 甘蔗ScSOS1蛋白二级结构预测分析 Table 3 Secondary structure prediction of sugarcane ScSOS1 protein
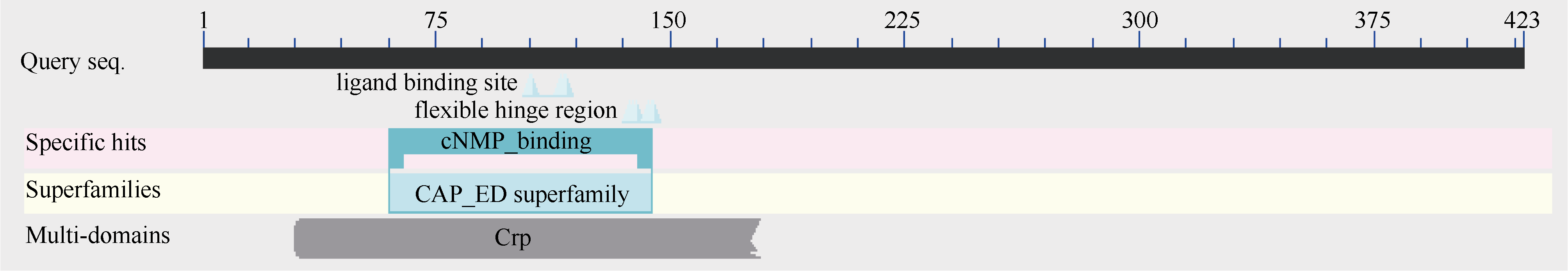
图3 甘蔗、高粱、水稻、小麦和玉米SOS1蛋白三级结构预测Fig. 3 Predicted third structure of SOS1 protein in sugarcane, sorghum, nice, wheat, and maize
表4 Table 4 表4(Table 4)
表4 甘蔗ScSOS1蛋白信号肽预测 Table 4 Signal P-NN prediction for sugarcane ScSOS1 protein
指标 Index
位点 Site
分值 Score
有无信号肽 Signal peptide
Max. C
40
0.130
Max. Y
40
0.125
Max. S
39
0.267
Mean S
1-39
0.112
D
1-39
0.118
Absence
C score: scores of Putative cleavage site; S score: scores of Signal peptide; Y score: scores of Synthesis cleavage site S-mean: the average of the S-score; D score: a weighted average of the S-mean and the Y-max scores. C score: 原始剪切位点的分值; S score: 信号肽的分值; Y score: 综合剪切位点的分值; S-mean: 信号肽分值的平均值; D score: S-mean和Y-max的加权平均值。
表4 甘蔗ScSOS1蛋白信号肽预测 Table 4 Signal P-NN prediction for sugarcane ScSOS1 protein
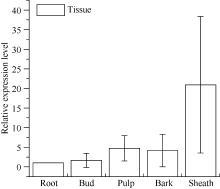
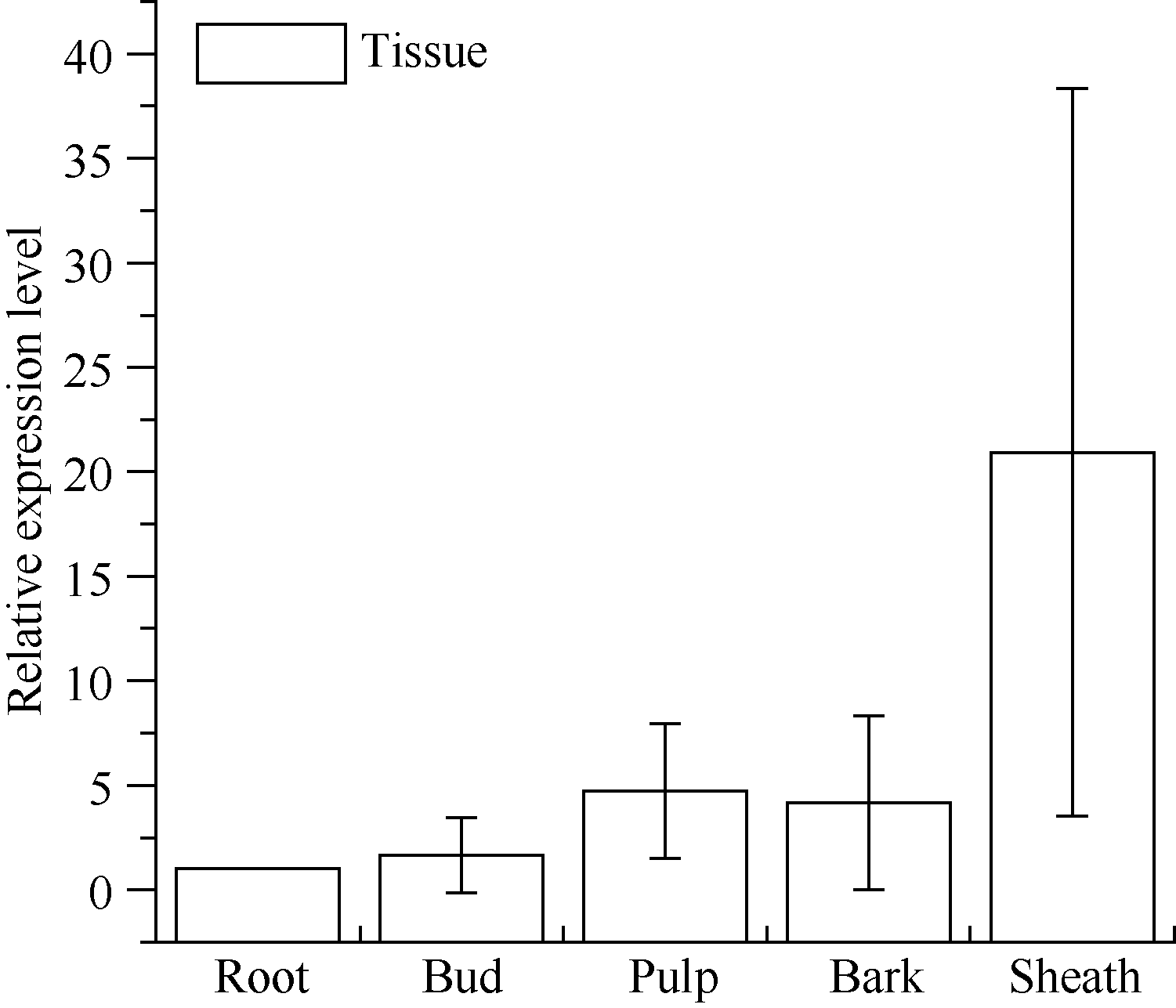
图7 甘蔗ScSOS1基因在不同组织中的表达 误差线为每组处理的标准误差(n = 3)。Fig. 7 Relative expression of ScSOS1 gene in different sugarcane tissues The error bars represent the standard error of each treating group (n= 3).
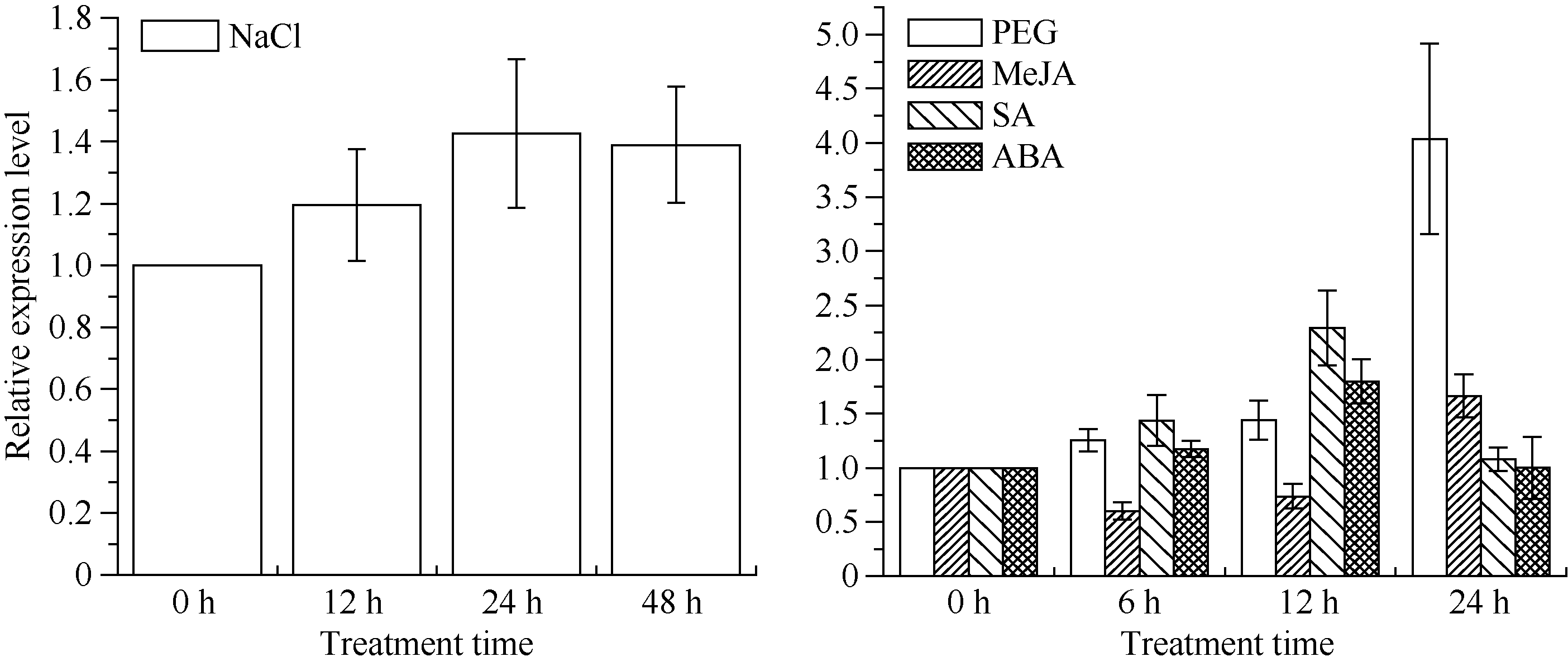
图8 甘蔗ScSOS1基因在不同外源胁迫下的表达 误差线为每组处理的标准误差(n=3)。Fig. 8ScSOS1 gene expression of sugarcane under different exogenous stresses The error bars represent the standard error of each treating group (n=3).
赵可夫, 李法曾, 樊守金, 冯立田. 中国的盐生植物. 植物学通报, 1999, 16: 201-207Zhao KF, Li FZ, Fan SJ, Feng LT. Halophytes in china. Chin Bull Bot, 1999, 16: 201-207 (in Chinese with English abstract)[本文引用:1]
[2]
Niu XM, Bressan R A. Hasegawa P M, Pardo JM. Ion homeostasis in NaCl stress environments. Plant Physiol, 1995, 109: 735-742[本文引用:1]
[3]
GreenwayH, MunnsR. Mechanisms of salt tolerance in nonhalophytes. Annu Rev Plant Physiol, 1980, 31: 149-190[本文引用:1]
[4]
Meloni DA, Oliva MA, Martinez CA, CambraiaJ. Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ Exp Bot, 2003, 49: 69-76[本文引用:1]
[5]
YeoA. Molecular biology of salt tolerance in the context of whole-plant physiology. J Exp Bot, 1998, 49: 915-929[本文引用:1]
[6]
Alberte RS, Thornber JP. Water stress effects on the content and organization of chlorophyll in mesophyll and bundle sheath chloroplasts of maize. Plant Physiol, 1977, 59: 351-353[本文引用:1]
[7]
Santos CV. Regulation of chlorophyll biosynthesis and degradation by salt stress in sunflower leaves. Sci Hort, 2004, 103: 93-99[本文引用:1]
[8]
刁丰秋, 章文华. 盐胁迫对大麦叶片类囊体膜组成和功能的影响. 植物生理学报, 1997, 23: 105-110Diao FQ, Zhang WH. Salt stress of barley leaf thylakoid membrane composition function. Acta Photophysiol Sin, 1997, 23: 105-110 (in Chinese with English abstract)[本文引用:1]
[9]
蒋明义, 杨文英, 徐江陈巧云. 渗透胁迫下水稻幼苗中叶绿体降解的活性氧损伤作用. 植物学报, 1994, 36: 289-295Jiang MY, Yang WY, XuJ, Chen QY. Active oxygen damage effect of chlorophyll degradation in rice seedlings under osmotic stress. Acta Bot Sin, 1994, 36: 289-295 (in Chinese with English abstract)[本文引用:1]
[10]
吴雪霞, 张永平, 查丁石. NaCl胁迫对茄子幼苗生长和光合特性的影响. 浙江农业学报, 2010, 22: 193-197Wu XX, Zhang YP, Zha DS. Effect of NaCl stress on growth and photosynthetic characteristics of eggplant seedlings. Acta Agric Zhejiangensis, 2010, 22: 193-197 (in Chinese with English abstract)[本文引用:1]
[11]
郑世英, 商学芳, 王丽燕, 张秀玲. 盐胁迫对不同基因型玉米生理特性和产量的影响. 干旱地区农业研究, 2010, 28: 109-112Zheng SY, Shang XF, Wang LY, Zhang XL. Changes of physiological characteristics and yield different salt-sensitive maize under salt stress. Agric Res Arid Areas, 2010, 28: 109-112 (in Chinese with English abstract)[本文引用:1]
[12]
武俊英, 刘景辉, 李倩. 盐胁迫对燕麦幼苗生长, K+、Na+吸收和光合性能的影响. 西北农业学报, 2010, 19: 100-105Wu JY, Liu JH, LiQ. Effects of salt stress on oat seedling growth and selective absorption of K+ and Na+ and photosynthetic characters. Acta Agric Boreali-Occident Sin, 2010, 19: 100-105 (in Chinese with English abstract)[本文引用:1]
[13]
赵自国, 陆静梅. 植物耐盐性研究及进展. 长春师范学院学报, 2002, 21(2): 51-53Zhao ZG, Lu JM. Progress or research in plant salt tolerance. J Chang Chun Teach Coll, 2002, 21(2): 51-53 (in Chinese with English abstract)[本文引用:1]
[14]
韩梦娴. Na+、K+、Ca2+对植物耐盐性影响的研究进展. 广东农业科学, 2009, (10): 81-83Han MX. Research progress in plant salt tolerance on Na+, K+, Ca2+. Guangdong Agric Sci, 2009, (10): 81-83 (in Chinese)[本文引用:1]
[15]
赵祥强. 植物耐盐性分子机理研究进展. 安徽农业科学, 2009, 37: 7844-7849Zhao XQ. Advances in studies on the molecular mechanism of plant’s salt tolerance. J Anhui Agric Sci, 2009, 37: 7844-7849 (in Chinese with English abstract)[本文引用:1]
[16]
杨少辉, 季静, 王罡. 盐胁迫对植物的影响及植物的抗盐机理. 世界科技研究与发展, 2006, 28(4): 70-76Yang SH, JiJ, WangG. Effects of salt stress on plant and the mechanism of salt tolerance. World Sci-tech R&D, 2006, 28(4): 70-76 (in Chinese with English abstract)[本文引用:2]
[17]
伍林涛, 杜才富, 邵明波. 植物盐胁迫耐受性研究进展. 吉林农业, 2010, (9): 51-52Wu LT, Du CF, Shao MB. Research progress in plant salt tolerance. Jilin Agric, 2010, (9): 51-52 (in Chinese with English abstract)[本文引用:1]
[18]
ChinnusamyV, SchumakerK, Zhu JK. Molecular genetic perspectives on cross-talk and specificity in abiotic stress signaling in plants. J Exp Bot, 2004, 55: 225-236[本文引用:1]
[19]
陈义强, 郭莺, 郭春芳, 张木清. 甘蔗斑茅属间远缘杂种后代对NaCl胁迫的响应. 热带作物学报, 2005, 26: 46-51Chen YQ, GuoY, Guo CF, Zhang MQ. Analysis of the hardiness of the intergeneric hybrids between Saccharum L. and Erianthus michx subjected to NaCl stress. Chin J Trop Crops, 2005, 26: 46-51 (in Chinese with English abstract)[本文引用:1]
[20]
Apes MP, Aharon GS, Snedden WA, BlumwaldE. Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science, 1999, 285: 1256-1258[本文引用:1]
[21]
WaditeeR, HibinoT, NakamuraT, IncharoensakdiA, TakabeT. Over expression of a Na+/H+ antiporter confers salt tolerance on a freshwater cyanobacterium, making it capable of growth in sea water. Proc Natl Acad Sci USA, 2002, 99: 4109-4114[本文引用:1]
[22]
Zhu JK. Genetic analysis of plant salt tolerance using Arabidopsis. Plant Physiol, 2000, 124: 941-948[本文引用:1]
[23]
Shi HZ, Xiong LM, StevensonB, Lu TG, Zhu JK. The Arabidopsis salt overly sensitive 4 mutants uncover a critical role for vitamin B6 in plant salt tolerance. Plant Cell, 2002, 14: 575-588[本文引用:1]
[24]
ShiH, Zhu JK. SOS4, a Pyridoxal kinase gene, is required for root hair development in Arabidopsis. Plant Physiol, 2002, 129: 585-593[本文引用:1]
[25]
Shi HZ, KinY, GuoY, StevensonB, Zhu JK. The Arabidopsis SOS5 locus encodes a putative cell surface adhesion protein and is required for normal cell expansion. Plant Cell, 2003, 15: 19-32[本文引用:3]
[26]
Zhu JH, Lee BH, DellingerM, Cui XP, Zhang CQ, WuS, Nothnagel EA, Zhu JK. A cellulose synthase-like protein is required for osmotic stress tolerance in Arabidopsis. Plant J, 2010, 63: 128-140[本文引用:1]
[27]
YamaguchiT, Apse MP, Shi HZ, BlumwaldE. Topological analysis of a plant vacuolar Na+/H+ antiporter reveals a luminal C terminus that regulates antiporter cation selectivity. Proc Natl Acad Sci USA, 2003, 100: 12510-12515[本文引用:2]
[28]
Zhu JK. Regulation of ion homeostasis under salt stress. Curr Opin Plant Biol, 2003, 6: 441-445[本文引用:2]
[29]
Zhang JZ, Creelman RA, Zhu JK. From laboratory to field. Using information from Arabidopsis to engineer salt, cold, and drought tolerance in crops. Plant Physiol, 2004, 135: 615-621[本文引用:2]
[30]
张莹. 互花米草SOS1基因和HKT1基因的克隆及耐盐转基因水稻研究. 烟台大学硕士学位论文, 山东烟台, 2009ZhangY. Clone SOS1, HKT1 of Spartina alterniflora and the Research of Salt-Tolerant Transgenic Rice. MS Thesis of Yantai University, Yantai, China, 2009 (in Chinese with English abstract)[本文引用:2]
[31]
权庚, 张侠, 尹海波, 郭善利. 过量表达SaSOS1水稻的幼苗鉴定及生理特性分析. 河南农业科学, 2015, 44(3): 14-18QuanG, ZhangX, Yin HB, Guo SL. Identification and physiological characterization of rice seedling overexpressing SaSOS1. J Henan Agric Sci, 2015, 44(3): 14-18 (in Chinese with English abstract)[本文引用:2]
[32]
邱生平. 水稻耐盐性的遗传分析及耐盐相关基因的克隆. 南京农业大学博士学位论文, 江苏南京, 2005Qiu SP. Genetic Analysis and Relative Gene Cloning for Salt Tolerance in Rice. PhD Dissertation of Nanjing Agricultural University, Nanjing, China, 2005 (in Chinese with English abstract)[本文引用:1]
[33]
王化波. 小麦盐胁迫应答基因的克隆及其功能研究. 中国科学院遗传与发育生物学研究所博士学位论文, 北京, 2004Wang HB. Isolation and Characterization of Salt-Induced Gene from Wheat. PhD Dissertation of Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing, China, 2004 (in Chinese with English abstract)[本文引用:1]
[34]
周玲玲, 祝建波, 曹连莆. 大叶补血草Na+/H+逆向转运蛋白基因(SOS1)的克隆与序列分析. 园艺学报, 2009, 36: 1353-1358Zhou LL, Zhu JB, Cao LP. Cloning and sequence analysis of a Na+/H+ antiporter gene in halophyte Limonium gmelinii. Acta Hortic Sin, 2009, 36, 1353-1358 (in Chinese with English abstract)[本文引用:1]
[35]
黄珑, 苏炜华, 张玉叶, 黄宁, 凌辉, 肖新换, 阙友雄, 陈如凯. 甘蔗CIPK基因的同源克隆与表达. 作物学报, 2015, 41: 499-506HuangL, Su WH, Zhang YY, HuangN, LingH, Xiao XH, Que YX, Chen RK. Cloning and expression analysis of CIPK gene in sugarcane. Acta Agron Sin, 2015, 41: 499-506 (in Chinese with English abstract)[本文引用:2]
[36]
Gand onouB, AgbanglaC, AhanhanzoC, ErrabiiT, IdaomarM, AbriniJ, Skali-SenhajiN. In vitro culture techniques as a tool of sugarcane bud germination study under salt stress. Afr J Biotechnol, 2010, 7: 3680-3682[本文引用:1]
[37]
AshrafM, Rahmatullah, AhmadR, Afzal M, Tahir M A, Kanwal S, Maqsood M A. Potassium and silicon improve yield and juice quality in sugarcane (Saccharum officinarum L. ) under salt stress. J Agron Crop Sci, 2009, 195: 284-291[本文引用:1]
[38]
张玉叶, 黄宁, 苏炜华, 肖新换, 罗俊, 阙友雄. 甘蔗苏氨酸脱氨酶基因的克隆与表达分析. 热带作物学报, 2014, 35: 59-67Zhang YY, HuangN, Su WH, Xiao XH, LuoJ, Que YX. Cloning and expression analysis of threonine deaminase gene in sugarcane. Chin J Trop Crops, 2014, 35: 59-67 (in Chinese with English abstract)[本文引用:1]
[39]
Guo JL, LingH, Wu QB, Xu LP, Que YX. The choice of reference genes for assessing gene expression in sugarcane under salinity and drought stresses. Sci Rep-UK, 2014, 4: 7042, DOI: DOI:10.1038/srep07042[本文引用:1]
[40]
黄宁, 张玉叶, 凌辉, 罗俊, 吴期滨, 阙友雄. 甘蔗二氨基庚二酸异构酶基因的克隆与表达分析. 热带作物学报, 2013, 34: 2200-2208HuangN, Zhang YY, LingH, LuoJ, Wu QB, Que YX. Cloning and expression analysis of a diaminopimelate epimerase gene in sugarcane. Chin J Trop Crops, 2013, 34: 2200-2208 (in Chinese with English abstract)[本文引用:1]
[41]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using Real-time quantitative PCR and the 2-∆∆CT method. Methods, 2001, 25: 402-408[本文引用:1]
[42]
Walker JM. The Proteomics Protocols Hand book. Totowa: Humana Press, 2005. pp 571-607[本文引用:1]
[43]
KyteJ, Doolittle RF. A simple method for displaying the hydropathic character of a protein. J Mol Biol, 1982, 57: 105-132[本文引用:1]
[44]
杨献光, 张榜军, 刘青兰, 史文超, 梁卫红. 盐胁迫应答基因OsSOS5的生物信息学分析. 河南师范大学学报(自然科学版), 2009, 37(6): 123-125Yang XG, Zhang BJ, Liu QL, Shi WC, Liang WH. Bioinformatics analysis of salt overly sensitive 5 gene in rice (Oryza sativa). Henan Norm Univ Life Sci (Nat Sci), 2009, 37(6): 123-125 (in Chinese with English abstract)[本文引用:1]
[45]
赵祥强. 玉米Na+/H+逆向转运蛋白基因ZmSOS1的克隆与鉴定. 安徽农业科学, 2009, 37: 17843-17848Zhao XQ. Cloning and identification of a new Na+/H+ antiporter gene ZmSOS1 in maize (Zea mays L. ). J Anhui Agric Sci, 2009, 37: 17843-17848 (in Chinese with English abstract)[本文引用:2]
[46]
Tang RJ, LiuH, BaoY, Lv QD, YangL, Zhang HX. The woody plant poplar has a functionally conserved salt overly sensitive pathway in response to salinity stress. Plant Mol Biol, 2010, 74: 367-380[本文引用:1]
[47]
徐立新, 于伟, 袁潜华. 木榄质膜型Na+/H+逆向转运蛋白的基因克隆与序列分析. 热带作物学报, 2012, 33: 1800-1807Xu LX, YuW, Yuan QH. Cloning of Na+/H+ an antiporter gene from Bruguiera gymnorrhiza (L. ) LAM. Chin J Trop Crops, 2012, 33: 1800-1807 (in Chinese with English abstract)[本文引用:1]
[48]
周玲玲, 祝建波, 王爱英. 过量表达大叶补草LgSOS1基因对拟南芥耐盐性的影响. 石河子大学学报(自然科学版), 2011, 29: 731-736Zhou LL, Zhu JB, Wang AY. Influence on salt tolerance of Arabidopsis thaliana by overexpressing LgSOS1. J Shihezi Univ (Nat Sci), 2011, 29: 731-736 (in Chinese with English abstract)[本文引用:2]
[49]
王姝杰, 王法龙, 李世访, 闫淑珍. 转Na+/H+ antiporter (Nhap)基因烟草植株的获得及耐盐性鉴定. 农业生物技术学报, 2006, 14: 74-78Wang SJ, Wang FL, Li SF, Yan SZ. Overexpression of Na+/H+ antiporter (Nhap) gene improves salt tolerance in tobacco. J Agric Biotechnol, 2006, 14: 74-78 (in Chinese with English abstract)[本文引用:2]
[50]
Shi HZ, Lee BH, Wu SJ, Zhu JK. Overexpression of a plasma membrane Na+/H+ antiporter gene improves salt tolerance in Arabidopsis thaliana. Nat Biotechnol, 2003, 21: 81-85[本文引用:2]
[51]
BenitoB, Rodríguez-NavarroA. Molecular cloning and characterization of a sodium-pump ATPase of the moss Physcomitrella patens. Plant J, 2003, 36: 382-389[本文引用:2]
[52]
Wu YX, DingN, ZhaoX, Zhao MG, Chang ZQ, Liu JQ, Zhang LX. Molecular characterization of PeSOS1: the putative Na+/H+ antiporter of Populus euphratica. Plant Mol Biol, 2007, 65: 1-11[本文引用:2]
[53]
GarciadeblásB, HaroR, BenitoB. Cloning of two SOS1 transportets from the seagrass Cymodocea nodosa. SOS1 transporters from Cymodocea and Arabidopsis potassium up take in bacteria. Plant Mol Biol, 2007, 63: 479-490[本文引用:2]
[54]
MunnsR, Sharp RE. Involvement of abscisic acid in controlling plant growth in soil of low water potential. Funct Plant Biol, 1993, 20: 425-437[本文引用:1]
[55]
张丽, 张华新, 杨升, 冯永巍. 植物耐盐机理的研究进展. 西南林学院学报, 2010, 30(3): 82-86ZhangL, Zhang HX, YangS, Feng YW. Research advances in plant salt-tolerance mechanism. J Southwest Univ, 2010, 30(3): 82-86 (in Chinese with English abstract)[本文引用:1]
[56]
Jia GL, Dai HP, Feng BL, . Zhang SQ, Zhang SW. Biochemical characteristics in broomcorn millet (Panicum miliaceum L. ) seedlings under PEG simulated drought stress. Acta Bot Boreali- Occident Sin, 2008, 28: 2073-2079[本文引用:1]
[57]
Hu XL, Jiang MY, Zhang JH, Zhang AY, LinF, Tan MP. Calcium-calmodulin is required for abscisic acid-induced antioxidant defense and functions both upstream and downstream of H2O2 production in leaves of maize (Zea mays) plants. New Phytol, 2007, 173: 27-38[本文引用:1]
[58]
谢崇波, 金谷雷, 徐海明, 朱军. 拟南芥在盐胁迫环境下SOS转录调控网络的构建及分析. 遗传, 2010, 32: 639-646Xie CB, Jin GL, Xu HM, ZhuJ. Construction and analysis of SOS pathway-related transcriptional regulatory network underlying salt stress response in Arabidopsis. Hereditas (Beijing), 2010, 32: 639-646 (in Chinese with English abstract)[本文引用:1]
 , 苏炜华
, 苏炜华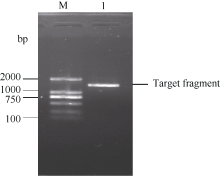











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}