关键词:水稻; 杂种不育基因; 基因定位 Identification and Mapping of a Hybrid Sterility Gene between 9311 and Nipponbare ZHANG Hong-Gen**, ZHANG Li-Jia**, SUN Yi-Biao, SI Hua, LIU Qiao-Quan, TANG Shu-Zhu*, GU Ming-Hong Jiangsu Key Laboratory of Crop Genetics and Physiology / Co-Innovation Center for Modern Production Technology of Grain Crops / Key Laboratory of Plant Functional Genomics of the Ministry of Education, Yangzhou University, Yangzhou 225009, China Fund:This study was supported by the Key Project of Chinese National Programs for Fundamental Research and Development (2011CB100101) and the Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions AbstractExploitation of subspecific heterosis is an effective method to improve rice yield by overcoming hybrid sterility between subspecies. In this study, F1 plants of the cross between Nipponbare and T9424, a line from a set of chromosome segment substitution lines with Nipponbare background as recipient and 9311 as donor, showed the decreasing spikelet and pollen fertility compared with the two parents, indicating that there was the incompatibility between the parents. Three substituted chromosome segments on chromosome 1, 4, and 5, respectively, were identified by whole genome re-sequencing of T9424. Analysis of the genotypes and spikelet fertility of plants in Nipponbare/T9424 F2 population indicated that hybrid sterility gene between T9424 and Nipponbare was located on chromosome 5. A total of 790 plants were then used for mapping the hybrid sterility gene, and the target gene was mapped to a candidate region with the physical distance of 110 kb between PSM8 and A14 on chromosome 5. The hybrid sterility gene, named S39(t) temporarily , controlled partial abortion of both pollen grains and embryo-sac of Nipponbare/T9424 F1 plants .These results are useful for deepening understanding of the phenomenon of hybrid sterility, and lay the groundwork for the gene cloning and its use in breeding.
表1 双亲及F1抽穗期、株高、花粉育性与小穗育性 Table 1 Heading stage, plant height, pollen and spikelet fertility of the parents and F1
双亲及F1 Parent and F1
抽穗期 Heading period (d)
株高 Plant height (cm)
花粉育性 Pollen sterility (%)
小穗育性 Spikelet fertility (%)
日本晴Nipponbare
94 a
102.01± 4.61 a
95.00± 2.13 a
91.93± 7.25 a
F1
94 a
89.31± 3.46 b
52.58± 12.98 b
65.36± 15.94 b
T9424
94 a
70.72± 5.60 c
94.00± 3.32 a
94.50± 2.50 a
Values are not significantly different and that followed by different letters are significantly different at the 0.05 probability level. 标明不同字母的值差异达0.05显著水平。
表1 双亲及F1抽穗期、株高、花粉育性与小穗育性 Table 1 Heading stage, plant height, pollen and spikelet fertility of the parents and F1
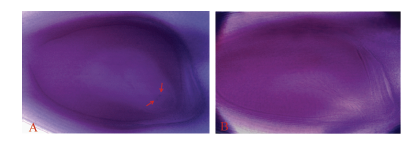
图1 双亲及F1的植株和花粉Fig. 1 Plant and pollen grains phenotype of the parents and F1 A: 日本晴、日本晴/T9424 F1及T9424植株表型; B、C、D: 分别为日本晴、日本晴/T9424 F1和T9424的花粉表型。 A: plant phenotypes of Nipponbare, Nipponbare/T9424 F1, and T9424; B, C, D: pollen grains phenotypes of Nipponbare, Nipponbare/T9424 F1, and T9424, respectively.
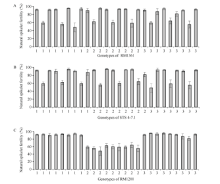
图3 不同基因型与小穗育性分布图Fig. 3 Distribution of different genotypes and spikelet fertility A、B、C分别为标记RM1361、STS 4-7.1、RM1200检测基因型与小穗育性分布图。 A, B, and C represent the genotypes of marker RM1361, STS4-7.1, and RM1200, respectively.
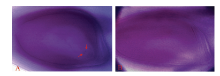
图5 T9424及日本晴/T9424 F1胚囊Fig. 5 Embryo-sac of T9424 and F1 A: T9424的胚囊; B: 日本晴/T9424 F1胚囊。箭头示极核。 A: embryo-sac of T9424; B: embryo-sac of Nipponbare/T9424 F1. The arrows show the polar nucleus.
SanoY. Sterility barriers between Oryza sativa and O. glaberrima. In: International Rice Research Institute, eds. Rice Genetics. Manila: International Rice Research Institute, 1986. pp 109-118[本文引用:1]
[2]
Orr HA, Presgraves DC. Speciation by postzygotic isolation: forces, genes and molecules. Bioessays, 2000, 22: 1085-1094[本文引用:1]
[3]
何光华, 郑家奎, 阴国大, 杨正林. 水稻亚种间杂种配子育性的研究. 中国水稻科学, 1994, 8: 177-180He GH, Zheng JK, Yin GD, Yang ZL. Game fertility of F1 between indica and japonica. Chin J Rice Sci, 1994, 8: 177-180 (in Chinese with English abstract)[本文引用:1]
[4]
王才林, 张兆兰, 汤述翥, 施建达. 三系法籼粳亚种间杂种优势利用研究: I. 籼粳交不育与细胞质雄性不育的区别及其检测. 江苏农业学报, 1992, 8(3): 1-7Wang CL, Zhang ZL, Tang SZ, Shi JD. Exploitation of heterosis between indica and japonica by three-line method I differentiation between indica-japonica sterility and cytoplasmic male sterility. Jiangsu J Agric Sci, 1992, 8(3): 1-7 (in Chinese with English abstract)[本文引用:1]
[5]
SanoY, SanoR. Variation of the intergenic spacer region of ribosomal DNA in cultivated and wild rice species. Genome, 1990, 33: 209-218[本文引用:1]
[6]
MaekawaM, InuakiT, ShinbashiN. Spikelet sterility in F1 hybrids between rice varieties Silewah and Hayakogane. Ikushugaku Zasshi, 1991, 41: 359-363[本文引用:1]
[7]
马生健, 刘耀光, 刘金祥. 水稻的杂种不育研究进展. 植物遗传资源学报, 2014, 15: 1080-1088Ma SJ, Liu YG, Liu JX. Research progress of hybrid sterility of rice. J Plant Genet Resour, 2014, 15: 1080-1088 (in Chinese with English abstract)[本文引用:1]
[8]
陆驹飞, 严长杰, 汤述翥, 朱立煌, 顾铭洪. 云南水稻品种花糯广亲和性的遗传分析. 扬州大学学报(自然科学版), 1998, 1(4): 31-35Lu JF, Yan CJ, Tang SZ, Zhu LH, Gu MH. Genetic analysis of the wide compatiblity of rice variety Huanuo from Yunnan province. J Yangzhou Univ (Nat Sci), 1998, 1(4): 31-35 (in Chinese with English abstract)[本文引用:1]
[9]
Yao SY, Henderson MT, Jodon NE. Cryptic structural hybridity as a probable cause of sterility in inter-varietal hybrids of cultivated rice, Oryza sativa L. Cytologia, 1958, 23: 46-55[本文引用:1]
[10]
OkaH. Analysis of genes controlling F1 sterility in rice by the use of isogenic lines. Genetics, 1974, 77: 521-534[本文引用:1]
[11]
Chen JJ, Ding JH, Ou-Yang Y D, Du H Y, Yang J Y, Cheng K. A triallelic system of S5 is a major regulator of the reproductive barrier and compatibility of indica-japonica hybrids in rice. Proc Natl Acad Sci USA, 2008, 105: 11436-11441[本文引用:1]
[12]
田华. 水稻籼粳亚种间杂种胚囊不育基因S7的精细定位及细胞学研究. 南京农业大学硕士学位论文, 江苏南京, 2009TianH. Cytological Studies and Gene Mapping of Inter- subspecies Hybrid Embryo Sac Sterility Gene S7 of Rice (Oryza sativa L. ). MS Thesis of Nanjing Agricultural University, Nanjing, China, 2009 (in Chinese with English abstract)[本文引用:1]
[13]
Singh SP, Sundaram RM, Biradar SK, Ahmed ML, Viraktamath BC, Siddiq EA. Identification of simple sequence repeat markers for utilizing wide-compatibility genes in inter-subspecific hybrids in rice (Oryza sativa L. ). Theor Appl Genet, 2006, 113: 509-517[本文引用:1]
[14]
WanJ, YamaguchiY, KatoH, IkehashiH. Two new loci for hybrid sterility in cultivated rice (Oryza sativa L. ). Theor Appl Genet, 1996, 92: 183-190[本文引用:1]
[15]
WanJ, IkehashiH. Identification of anew locus S16 causing hybrid sterility in native rice varieties (Oryza sativa L. ) from Taihu Lake region and Yunnan province, China. Breed Sci, 1995, 45: 461-470[本文引用:1]
[16]
Wan JM, IkehashiH, SakaiM, HorisueH, ImbeT. Mapping of hybrid sterility gene S17 of rice (Oryza sativa L. ) by isozyme and RFLP markers. Rice Genet Newsl, 1998, 15: 151-154[本文引用:1]
[17]
ZhuS, WangC, ZhengT, ZhaoZ, IkehashiH, WanJ. A new gene located on chromosome 2 causing hybrid sterility in a remote cross of rice. Plant Breed, 2005, 124: 440-445[本文引用:1]
[18]
Zhu SS, JiangL, Wang CM, Zhai HQ, Li DT, Wan JM. The origin of weedy rice Ludao in China deduced by genome wide analysis of its hybrid sterility genes. Breed Sci, 2005, 55: 409-414[本文引用:1]
[19]
Zhao ZG, JiangL, Zhang WW, Yu CY, Zhu SS, XieK, TianH, Liu LL, IkehashiH, Wan JM. Fine mapping of S31, a gene responsible for hybrid embryo-sac abortion in rice (Oryza sativa L. ). Planta, 2007, 226: 1087-1096[本文引用:3]
[20]
Li DT, Chen LM, LingJ, Zhu SS, Zhao ZG, Liu SJ, SuN, Zhai HQ, IkehashiH, Wan JM. Fine mapping of S32(t), a new gene causing hybrid embryo sac sterility in a Chinese land race rice (Oryza sativa L. ). Theor Appl Genet, 2007, 114: 515-524[本文引用:1]
[21]
ChenM, ZhaoZ, JiangL, WanJ. A new gene controlling hybrid sterility in rice (Oryza sativa L. ). Euphytica, 2012, 184: 15-22[本文引用:1]
[22]
张桂权, 卢永根. 栽培稻( Oryza sativa)杂种不育性的遗传研究: I. 等基因F1不育系杂种不育性的双列分析. 中国水稻科学, 1989, 3: 97-101Zhang GQ, Lu YG. Genetic studies on the hybrid sterility in cultivated rice (Oryza sativa): I. Diallel analysis of the hybrid sterility among isogenic F1 sterile lines. Chin J Rice Sci, 1989, 3: 97-101 (in Chinese with English abstract)[本文引用:1]
[23]
张桂权, 卢永根. 栽培稻杂种不育性的遗传研究: II. F1花粉不育性的基因模式. 遗传学报, 1993, 20: 541-551Zhang GQ, Lu YG. Genetic studies on the hybrid sterility in cultivated rice (Oryza sativa): II. A genic model for F1 pollen sterility. Acta Genet Sin, 1993, 20: 222-228 (in Chinese with English abstract)[本文引用:1]
[24]
张桂权, 卢永根, 张华, 杨进昌, 刘桂富. 栽培稻( Oryza sativa)杂种不育性的遗传研究: IV. F1花粉不育性的基因型遗传. 遗传学报, 1994, 21: 34-41Zhang GQ, Lu YG, ZhangH, Yang JC, Liu GF. Genetic studies on the hybrid sterility in cultivated rice (Oryza sativa): IV. Genotypes for F1 pollen sterility. Acta Genet Sin, 1994, 21: 34-41 (in Chinese with English abstract)[本文引用:1]
[25]
夏继星. 水稻矮化突变体的分子遗传学分析和杂种不育基因S20的图位克隆. 华南农业大学博士学位论文, 广东广州, 2008Xia JX. Genetic Analysis of a Rice Dwarf Mutant Z110-6 and Molecular Cloning of S20 for Hybrid Male Sterility in Rice. PhD Dissertation of South China Agricultural University, Guangzhou, China, 2008 (in Chinese with English abstract)[本文引用:1]
[26]
TakahikoK, AtsushiY, NoriK. Hybrid male sterility in rice is due to epistatic interactions with a pollen killer locus. Genetics, 2011, 189: 1083-1092[本文引用:1]
[27]
Win KT, KuboT, MiyazakiY, DoiK, YamagataY, YoshimuraA. Identification of two loci causing F1 pollen sterility in inter- and intraspecific crosses of rice. Breed Sci, 2009, 59: 411-418[本文引用:1]
[28]
Long YM, Zhao LF, Niu BX, SuJ, WuH, Chen YL, Zhang QY, Guo JX, Zhuang CX, Mei MT, Xia JX, WangL, Wu HB, Liu YG. Hybrid male sterility in rice controlled by interaction between divergent alleles of two adjacent genes. Proc Natl Acad Sci USA, 2008, 105: 18871-18876[本文引用:1]
[29]
YamagataY, YamamotoE, AyaK, Win KT, DoiK, Sobrizal, ItoT, Kanamori H, Wu J Z, Matsumoto T, Matsuoka M, Ashikari M, Yoshimura A. Mitochondrial gene in the nuclear genome induces reproductive barrier in rice. Proc Natl Acad Sci USA, 2010, 107: 1494-1499[本文引用:2]
[30]
MizutaY, HarushimaY, KurataN. Rice pollen hybrid incompatibility caused by reciprocal gene loss of duplicated genes. Proc Natl Acad Sci USA, 2010, 107: 20417-20422[本文引用:2]
[31]
SanoY, SanoR. Variation of the intergenic spacer region of ribosomal DNA in cultivated and wild rice species. Genome, 1990, 33: 209-218[本文引用:1]
[32]
KoideY, OnishiK, NishimotoD, Baruah AR, KanazawaA, SanoY. Sex-independent transmission ratio distortion system responsible for reproductive barriers between Asian and African rice species. New Phytol, 2008, 179: 888-900[本文引用:1]
[33]
SanoY, SanoR, EiguchiM, Hirano HY. Gamete eliminator adjacent to the wx locus as recealed by pollen analysis in rice. J Hered, 1994, 85: 310-312[本文引用:1]
[34]
SanoY. Genetic comparisons of chromosome 6 between wild and cultivated rice. Jpn J Breed, 1992, 42: 561-572[本文引用:1]
赵世绪, 杜中, 凌祖铭. 用子房整体透明法和微分干涉差显微镜研究水稻的胚胎发育. 遗传, 1993, 15: 33-33Zhao SX, DuZ, Ling ZM. The use of a whole clearing technique and differential interference contrast microscope for study on embryology of rice. Hereditas (Beijing), 1993, 15: 33-33 (in Chinese)[本文引用:1]
[37]
Murray MG, Thompson WF. Rapid isolation of high molecular weight plant DNA. Nucl Acid Res, 1980, 8: 4321-4325[本文引用:1]
[38]
李文涛, 曾瑞珍, 张泽民, 丁效华, 张桂权. 水稻F1花粉不育基因座S-b的精细定位. 科学通报, 2006, 51: 404-408Li WT, Zeng RZ, Zhang ZM, Ding XH, Zhang GQ. Fine mapping of F1 pollen sterility loci S-b in rice. Chin Sci Bull, 2006, 51: 404-408 (in Chinese)[本文引用:3]
[39]
KuboT, YoshimuraA, KurataN. Hybrid male sterility in rice is due to epistatic interactions with a pollen killer locus. Genetics, 2011, 18: 1083-1092[本文引用:2]
[40]
Wang GW, He YQ, Xu CG, ZhangQ. Fine mapping of f5-Du, a gene conferring wide-compatibility for pollen fertility in inter- subspecific hybrids of rice (Oryza sativa L. ). Theor Appl Genet, 2006, 112: 382-387[本文引用:2]
[41]
IkehashiH, ArakiH. Varietal screening of compatibility types revealed in F1 fertility of distant crosses in rice. Ikushugaku Zasshi, 1984, 34: 304-313[本文引用:1]
[42]
ZhangH, ZhaoQ, Sun ZZ, Zhang CQ, FengQ, Tang SZ, Liang GH, Gu MH, HanB, Liu Q Q. Development and high- throughput genotyping of substitution lines carrying the chromosome segments of indica9311 in the background of japonica Nipponbare. J Genet Genomics, 2011, 38: 603-611[本文引用:1]
[43]
KitamuraE. Studies on cytoplasmic sterility of hybrids in distantly related: varieties of rice (Oryza sativa L. ). Jpn J Breed, 1962, 12: 81-84[本文引用:1]
 , 顾铭洪
, 顾铭洪







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}