关键词:水稻; ospls3; 叶片早衰; 生理分析; 基因定位 Physiological Characteristics and Gene Mapping of a Precocious Leaf Senescence Mutant ospls3 in Rice GONG Pan1,**, LI Kun-Yu1,**, HUANG Fu-Deng2, WEI Li-Quan1, YANG Xi1, CHENG Fang-Min1, PAN Gang1,* 1 College of Agriculture and Biotechnology, Zhejiang University, Hangzhou 310058, China
2 Zhejiang Academy of Agricultural Sciences, Hangzhou 310021, China
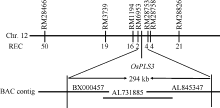
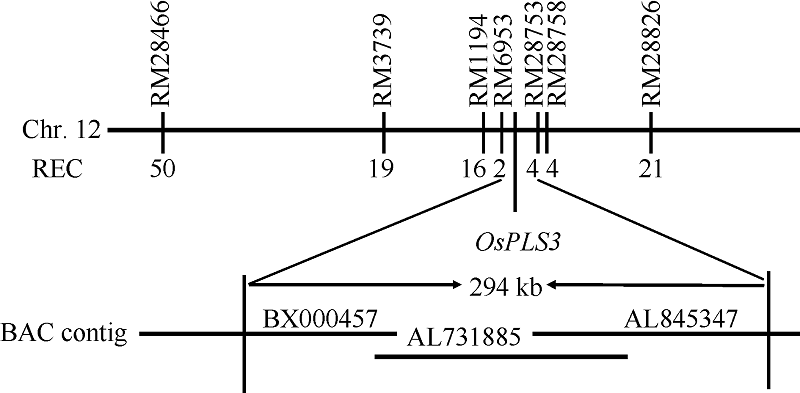
Fund:This study was supported by the National Natural Science Foundation of China (31271691) and the Major Project of China on New Varieties of GMO Cultivation (2013ZX08001-002) AbstractLeaf senescence is the final stage of leaf development. However, premature aging of functional leaves leads to yield reduction and quality decline. Thus, it is very important for developing novel crop germplasms with delayed leaf-senescence characteristics through investigating the molecular mechanism of leaf senescence. In this study, an ospls3 ( Oryza sativa precocious leaf senescence 3) mutant, produced by60Co γ-radiation treatment of indica cultivar N142, was identified. The symptoms of the premature senescence mutant presented firstly at tillering stage showing brown leaf tip and brown spots in top part of leaf blade, then spread rapidly to basal part of leaf blade and led leaf to die. The physiological analysis indicated that, in the ospls3 mutant, the content of chlorophyll was the highest in the flag leaf, the following was in second-top and third-top leaves, but all of them were significantly lower than those in the wild type. The contents of MDA, O2?, and H2O2 and the activities of SOD and POD among the top three leaves in the wild type maintained similar levels, which were significantly lower than those in the mutant. The soluble protein contents and the activity of CAT had no significant difference among top three leaves in the wild type while significantly decreased in the mutant. Genetic analysis verified that the ospls3 is a recessive mutant and was mapped in a 294 kb interval between RM6953 and RM28753 on the long arm of chromosome 12, which establishes a solid foundation for further cloning and functional studies of this gene.
Keyword:Rice; ospls3; Precocious leaf senescence; Physiological analysis; Gene mapping Show Figures Show Figures
图1 不同生育期突变体ospls3及其野生型的表型A: 分蘖期; B: 抽穗期; C: 灌浆结实期; D: 抽穗期叶片(1~4代表剑叶至倒四叶); Bar = 20 cm。Fig. 1 Phenotype of ospls3 and its wild-type (WT) plants at different growth stages A: tillering stage; B: heading stage; C: filling stage; D: leaves at heading stage (1-4 means the flag leaf to 4th leaf from top); Bar = 20 cm.
表1 Table 1 表1(Table 1)
表1 突变体ospls3及其野生型的农艺性状 Table 1 Agronomic traits of ospls3 and its wild-type (WT) plants
性状 Trait
2013
2014
野生型WT
突变体ospls3
野生型WT
突变体ospls3
株高 Plant height (cm)
112.32± 2.83
81.19± 2.56* *
109.78± 2.78
79.31± 3.45* *
有效穗数 Effective panicle number
11.52± 2.43
7.34± 1.25* *
12.74± 2.79
9.15± 0.84* *
每穗粒数 Grain number per panicle
175.79± 24.84
47.71± 9.17* *
167.43± 25.94
58.42± 12.65* *
结实率 Seed-setting rate (%)
88.05± 3.92
28.85± 2.84* *
80.34± 5.35
19.39± 4.11* *
千粒重 1000-grain weight (g)
25.33± 0.79
16.65± 1.13* *
24.68± 0.74
15.34± 1.12* *
单株产量 Yield per plant (g)
47.27± 5.32
2.78± 0.56* *
44.35± 7.72
1.69± 0.42* *
* * 表示在0.01水平上差异显著。* * Significant difference at P< 0.01.
表1 突变体ospls3及其野生型的农艺性状 Table 1 Agronomic traits of ospls3 and its wild-type (WT) plants
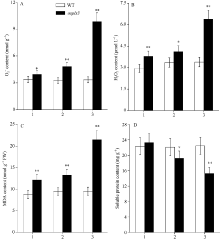
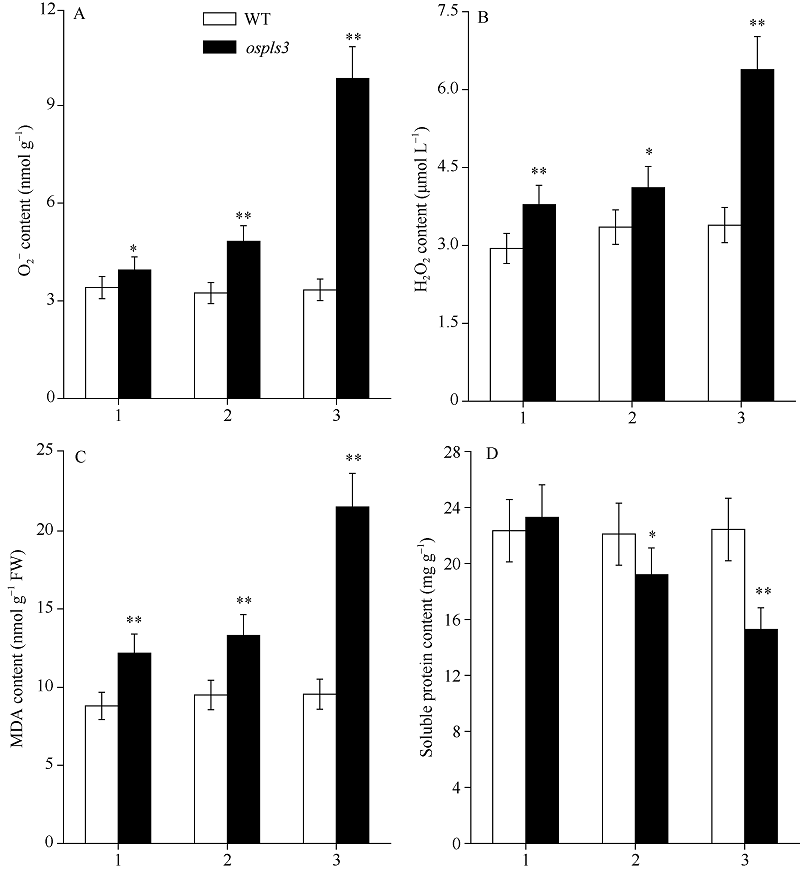
图3 孕穗期突变体ospls3及其野生型叶片的O2?、H2O2、MDA和可溶性蛋白含量1: 剑叶; 2: 倒二叶; 3: 倒三叶。* 表示在0.05水平上差异显著; * * 在0.01水平上差异显著。Fig. 3 O2?, H2O2, MDA, and soluble protein contents of ospls3 and its wild-type (WT) plants at booting stage1: flag leaves; 2: 2nd leaves from top; 3: 3rd leaves from top. * Significant difference at P< 0.05; * * Significant difference at P< 0.01.
Lim PO, Kim HJ, Nam HG. Leaf senescence. Annu Rev Plant Biol, 2007, 58: 115-136[本文引用:3]
[2]
伍泽堂. 超氧自由基与叶片衰老时叶绿素破坏的关系. 植物生理学通讯, 1991, 27: 277-279Wu ZT. Relationship between superoxide radical and destruction of chlorophyll during leaf senescence. Plant Physiol Commun, 1991, 27: 277-279 (in Chinese with English abstract)[本文引用:2]
[3]
段俊, 梁承邺, 黄毓文. 杂交水稻开花结实期间叶片衰老. 植物生理学报, 1997, 23: 139-144DuanJ, Liang CY, Huang YW. Studies on leaf senescence of hybrid rice at flowering and grain formation stage. Acta Phytophysiol Sin, 1997, 23: 139-144 (in Chinese with English abstract)[本文引用:1]
[4]
Zhang CJ, Chu HJ, Chen GX, Shi DW, ZuoM, WangJ, Lu CG, WangP, ChenL. Photosynthetic and biochemical activities in flag leaves of a newly developed superhigh-yield hybrid rice (Oryza sativa) and its parents during the reproductive stage. J Plant Res, 2007, 120: 209-217[本文引用:1]
[5]
InadaN, SakaiA, KuroiwaH, KuroiwaT. Senescence program in rice (Oryza sativa L. ) leaves: analysis of the blade of the second leaf at the tissue and cellular levels. Protoplasma, 1999, 207: 222-232[本文引用:1]
ThomasH, Smart CM. Crops that stay green. Ann Appl Biol, 1993, 123: 193-219[本文引用:1]
[8]
魏道智, 戴新宾, 许晓明, 张荣铣. 植物叶片衰老机理的几种假说. 广西植物, 1998, 18(1): 90-97Wei DZ, Dai XB, Xu XM, Zhang RX. Several hypotheses on the mechanism of the plant leaf senescence. Guihaia, 1998, 18(1): 90-97 (in Chinese with English abstract)[本文引用:1]
[9]
WangZ, WangY, HongX, HuD, LiuC, YangJ, LiY, HuangY, FengY, GongH, LiY, FangG, TangH, LiY. Functional inactivation of UDP-N-acetylglucosamine pyrophosphorylase 1 (UAP1) induces early leaf senescence and defence responses in rice. J Exp Bot, 2015, 66: 973-987[本文引用:1]
[10]
LiangC, WangY, ZhuY, TangJ, HuB, LiuL, OuS, WuH, SunX, ChuJ, ChuC. OsNAP connects abscisic acid and leaf senescence by fine-tuning abscisic acid biosynthesis and directly targeting senescence-associated genes in rice. Proc Natl Acad Sci USA, 2014, 111: 10013-10018[本文引用:2]
HanM, KimC, LeeJ, LeeS, JeonJ. OsWRKY42 represses OsMT1d and induces reactive oxygen species and leaf senescence in rice. Mol Cells, 2014, 37: 532-539[本文引用:2]
[13]
GaoQ, YangZ, ZhouY, YinZ, QiuJ, LiangG, XuC. Characterization of an Abc1 kinase family gene OsABC1-2 conferring enhanced tolerance to dark-induced stress in rice. Gene, 2012, 498: 155-163[本文引用:1]
[14]
ParkS, YuJ, ParkJ, LiJ, YooS, LeeN, LeeS, JeongS, Seo HS, KohH, JeonJ, ParkY, PaekN. The senescence-induced stay green protein regulates chlorophyll degradation. Plant Cell, 2007, 19: 1649-1664[本文引用:1]
[15]
JiangH, LiM, LiangN, YanH, WeiY, XuX, LiuJ, XuZ, ChenF, WuG. Molecular cloning and function analysis of the stay green gene in rice. Plant J, 2007, 52: 197-209[本文引用:1]
[16]
LeeR, HsuJ, HuangH, LoS, Chen SG. Alkaline alpha- galactosidase degrades thylakoid membranes in the chloroplast during leaf senescence in rice. New Phytol, 2009, 184: 596-606[本文引用:1]
[17]
JiaoB, WangJ, ZhuX, ZengL, LiQ, HeZ. A novel protein RLS1 with NB-ARM domains is involved in chloroplast degradation during leaf senescence in rice. Mol Plant, 2012, 5: 205-217[本文引用:1]
[18]
SatoY, MoritaR, KatsumaS, NishimuraM, TanakaA, KusabaM. Two short-chain dehydrogenase/reductases, NON-YELLOW COLORING 1 and NYC1-LIKE, are required for chlorophyll b and light-harvesting complex II degradation during senescence in rice. Plant J, 2009, 57: 120-131[本文引用:1]
[19]
KusabaM, ItoH, MoritaR, IidaS, SatoY, FujimotoM, KawasakiS, TanakaR, HirochikaH, NishimuraM, TanakaA. Rice NON-YELLOW COLORING1 is involved in light-harvesting complex II and grana degradation during leaf senescence. Plant Cell, 2007, 19: 1362-1375[本文引用:1]
[20]
MoritaR, SatoY, MasudaY, NishimuraM, KusabaM. Defect in non-yellow coloring 3, an alpha/beta hydrolase-fold family protein, causes a stay-green phenotype during leaf senescence in rice. Plant J, 2009, 60: 1110-1110[本文引用:1]
[21]
YamataniH, SatoY, MasudaY, KatoY, MoritaR, FukunagaK, NagamuraY, NishimuraM, SakamotoW, TanakaA, KusabaM. NYC4, the rice ortholog of Arabidopsis THF1, is involved in the degradation of chlorophyll protein complexes during leaf senescence. Plant J, 2013, 74: 652-662[本文引用:1]
[22]
QiaoY, JiangW, LeeJ, ParkB, ChoiM, PiaoR, WooM, RohJ, HanL, PaekN, Seo HS, KohH. SPL28 encodes a clathrin-associated adaptor protein complex 1, medium subunit μ1 (AP1M1) and is responsible for spotted leaf and early senescence in rice (Oryza sativa). New Phytol, 2010, 185: 258-274[本文引用:1]
[23]
Undan JR, TamiruM, AbeA, YoshidaK, KosugiS, TakagiH, YoshidaK, KanzakiH, SaitohH, FekihR, SharmaS, UndanJ, YanoM, TerauchiR. Mutation in OsLMS, a gene encoding a protein with two double-strand ed RNA binding motifs, causes lesion mimic phenotype and early senescence in rice (Oryza sativa L. ). Genes Genet Syst, 2012, 87: 169-179[本文引用:1]
[24]
张治安, 陈展宇. 植物生理学实验技术. 长春: 吉林大学出版社, 2008. p 7Zhang ZA, Chen ZY. Experiment Technology of Plant Physiology. Changchun: Jilin University Press, 2008. p 7 (in Chinese)[本文引用:1]
[25]
Rogers SO, Bendich AJ. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol Biol, 1985, 5: 69-76[本文引用:1]
[26]
Shen YJ, JiangH, Jin JP, Zhang ZB, XiB, He YY, WangG, WangC, Qian LL, LiX, Yu QB, Liu HJ, Chen DH, Gao JH, HuangH, Shi TL, Yang ZN. Development of genome-wide DNA polymorphism database for map-based cloning of rice genes. Plant Physiol, 2004, 135: 1198-1205[本文引用:1]
[27]
PanaudO, ChenX, McCouchS R. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSR) in rice (Oryza sativa L. ). Mol Gen Genet, 1996, 252: 597-607[本文引用:1]
[28]
李晴, 朱玉贤. 植物衰老的研究进展及其在分子育种中的应用. 分子植物育种, 2003, 1: 289-296LiQ, Zhu YX. The progress of plant senescence research and plant molecular breeding. Mol Plant Breed, 2003, 1: 289-296 (in Chinese with English abstract)[本文引用:1]
[29]
刘贞琦, 刘振业, 马达鹏, 曾淑芬. 水稻叶绿素含量及其与光合速率关系的研究. 作物学报, 1984, 10: 57-62Liu ZQ, Liu ZY, Ma DP, Zeng SF. A study on the relation between chlorophyll content and photosynthetic rate of rice. Acta Agron Sin, 1984, 10: 57-62 (in Chinese with English abstract)[本文引用:1]
[30]
林植芳, 李双顺, 林桂珠, 孙谷畴, 郭俊彦. 水稻叶片的衰老与超氧物歧化酶活性及脂质过氧化作用的关系. 植物学报, 1984, 26: 605-615Lin ZF, Li SS, Lin GZ, Sun GC, Guo JY. Superoxide dismutase activity and lipid peroxidation in relation to senescence of rice leaves. Acta Bot Sin, 1984, 26: 605-615 (in Chinese with English abstract)[本文引用:1]
[31]
HidegE, KalaiT, Kos PB, AsadaK, HidegK. Singlet oxygen in plants: its significance and possible detection with double (fluorescent and spin) indicator reagents. Photochem Photobiol, 2006, 82: 1211-1218[本文引用:1]
[32]
华春, 王仁雷. 杂交稻及其三系叶片衰老过程中SOD、CAT活性和MDA含量的变化. 西北植物学报, 2003, 23: 406-409HuaC, Wang RL. Changes of SOD and CAT activities and MDA content during senescence of hybrid rice and three lines leaves. Acta Bot Boreali-Occident Sin, 2003, 23: 406-409 (in Chinese with English abstract)[本文引用:1]
[33]
金杨, 周丽芬, 陈析丰, 刘峰, 马伯军. 水稻类病变突变体spl5细胞坏死机制的分析. 浙江师范大学学报(自然科学版), 2009, 32: 326-331JinY, Zhou LF, Chen XF, LiuF, Ma BJ. Mechanisms of cell death in rice lesion mimic mutant spl5. J Zhejiang Norm Univ (Nat Sci), 2009, 32: 326-331 (in Chinese with English abstract)[本文引用:1]
[34]
汪媛. 水稻叶片衰老过程生理变化及蛋白质降解与蛋白酶活性变化研究. 扬州大学硕士学位论文, 江苏扬州, 2010WangY. The Research of Physiological Changes, Protein Degradation and Protease Activity in the Process of Leaf Senescence in Rice. MS Thesis of Yangzhou University, Yangzhou, China, 2010 (in Chinese with English abstract)[本文引用:1]
[35]
赵晨晨, 黄福灯, 龚盼, 杨茜, 程方民, 潘刚. 水稻叶片早衰突变体osled的生理特征与基因定位. 作物学报, 2014, 40: 1946-1955Zhao CC, Huang FD, GongP, YangQ, Cheng FM, PanG. Physiological characteristics and gene mapping of a leaf early-senescence mutant osled in rice. Acta Agron Sin, 2014, 40: 1946-1955 (in Chinese with English abstract)[本文引用:1]
Guo YF, Gan SS. AtNAP, a NAC family transcription factor, has an important role in leaf senescence. Plant J, 2006, 46: 601-612[本文引用:1]
[38]
ZhangK, XiaX, ZhangY, GanS. An ABA-regulated and Golgi-localized protein phosphatase controls water loss during leaf senescence in Arabidopsis. Plant J, 2012, 69: 667-678[本文引用:1]
[39]
KongZ, LiM, YangW, XuW, XueY. A novel nuclear-localized CCCH-type zinc finger protein, OsDOS, is involved in delaying leaf senescence in rice. Plant Physiol, 2006, 141: 1376-1388[本文引用:1]
[40]
JanA, MaruyamaK, TodakaD, KidokoroS, AboM, YoshimuraE, ShinozakiK, NakashimaK, Yamaguchi-ShinozakiK. OsTZF1, a CCCH-tand em zinc finger protein, confers delayed senescence and stress tolerance in rice by regulating stress-related genes. Plant Physiol, 2013, 161: 1202-1216[本文引用:1]
[41]
PitakrattananukoolS, KawakatsuT, AnuntalabhochaiS, TakaiwaF. Overexpression of OsRab7B3, a small GTP-binding protein gene, enhances leaf senescence in transgenic rice. Biosci Biotechnol Biochem, 2012, 76: 1296-1202[本文引用:1]
[42]
LiY, ChenL, MuJ, ZuoJ. LESION SIMULATING DISEASE1 interacts with catalases to regulate hypersensitive cell death in Arabidopsis. Plant Physiol, 2013, 163: 1059-1070[本文引用:1]
[43]
SmykowskiA, ZimmermannP, ZentgrafU. G-Box binding factor1 reduces CATALASE2 expression and regulates the onset of leaf senescence in Arabidopsis. Plant Physiol, 2010, 153: 1321-1331[本文引用:1]






{kind=link}
{kind=link}
{kind=link}
{kind=link}