关键词:玉米; 光照; 中胚轴伸长; 多胺氧化酶; 表达分析 Physiological Mechanism Regulating Light-induced Mesocotyl Elongation by Polyamine Oxidase (PAO) in Maize ZHANG Tong-Zhen1,**, LI Yong-Sheng1,**, LI Yue2, YAO Hai-Mei1, ZHAO Juan1, WANG Chan1, ZHAO Yang1, WANG Han-Ning1, FANG Yong-Feng1,*, HU Jin3,* 1Gansu Key Laboratory of Crop Improvement and Germplasm Enhancement / Gansu Provincial Key Laboratory of Aridland Crop Science / Gansu Agricultural University, Lanzhou 730070, China
2Institute of Crop Science, Gansu Academy of Agricultural Sciences, Lanzhou 730070, China
3College of Agriculture & Biotechnology, Zhejiang University, Hangzhou 310058, China
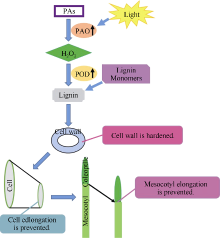
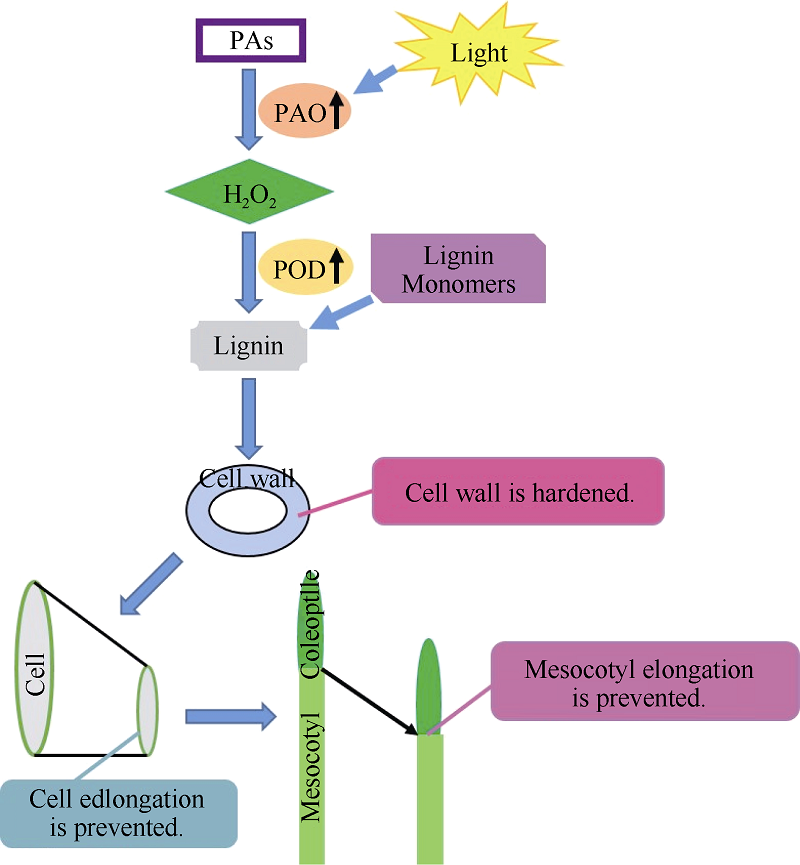
Fund:This study was supported by National Basic Research Program of China (973 Program, 2012CB722902), the National Natural Science Foundation of China (31371708, 31201279), and Research Program Sponsored by Gansu Provincial Key Laboratory of Aridland Crop Science (GSCS201210) AbstractIn this study, maize inbred line PH4CV was cultured under in MS medium under both dark and light conditions to investigate the relationship of length of mesocotyl with activity of polyamine oxidase (PAO), activity of peroxidase (POD), contents of H2O2 and lignin. In addition, after treated with 5 mmol L-1 2-hydroxyethylhydrazine (2-HEH) and 5 mmol L-1 N, N’-Dimethylthiourea (DMTU), the origin and accumulation site of H2O2 in mesocotyl elongation were assessed by using histochemical staining method and the effect of light on expression of ZmPAO gene was evaluated. The results indicated that the elongation of mesocotyl was significantly inhibited while the contents of H2O2 and lignin and the activities of POD and PAO were enhanced by light. Correlation analysis showed that the length of mesocotyl was negatively correlated with the PAO activity, H2O2 content, POD activity, and lignin content, and the PAO activity was positively correlated with the H2O2 content, the POD activity, and the lignin content. The assay with 2-HEH and DMTU, the microgram of epidermal cells, and histochemical localization of H2O2 revealed that H2O2 participated in mesocotyl elongation, in which PAO catalyzed polyamines (PAs) degradation to produce H2O2 resulting in the inhibition of mesocotyl elongation. Results of qRT-PCR revealed that the expression level of ZmPAO was relatively stable in dark and rose rapidly of 0.5 hour after exposing to light, with a maximum value after three hours of light treatment, then declined gradually, and finally showed a steady level after ten hours. This study suggests that the activity of PAO can be promoted by light treatment, and initiate the PAs-mediated induction of H2O2, resulting in the oxidation of lignin monomers on cell wall by POD. The produced free radicals are then transformed into lignin in a polyforming process which marked the cell wall hardened and cell elongation inhibited. This study may provide a theoretical basis for understanding the physiological mechanism underlying the PAO-mediated mesocotyl elongation and gain insights into the response of maize mesocotyl to the light stress.
Keyword:Maize; Light; Mesocotyl elongation; Polyamine oxidase; Expression analysis Show Figures Show Figures
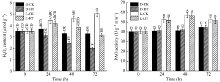
图1 光对玉米中胚轴长度及生物量积累的影响* * 表示在0.01水平上差异显著; * 表示在0.05水平上差异显著。Fig. 1 Effects of light on length and biomass accumulation of mesocotyl in maize* * means significant difference at the 0.01 level; * means significant difference at the 0.05 level.
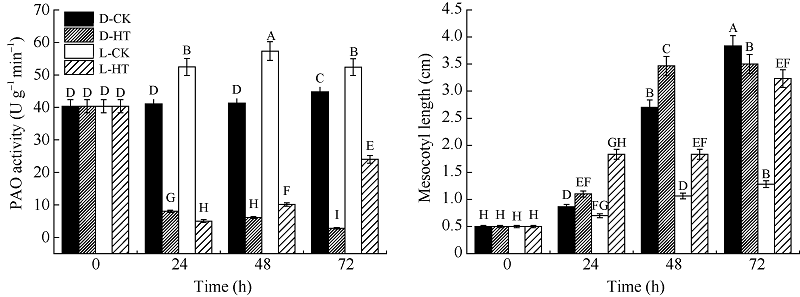
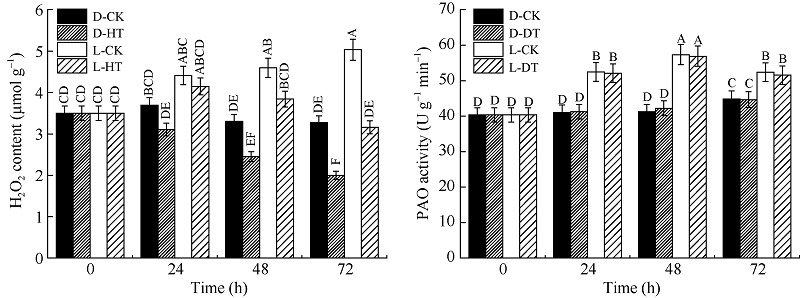
图2 光对玉米中胚轴PAO活性及长度的影响D-CK: 黑暗对照; D-HT: 黑暗, 5 mmol L-1 2-羟乙基肼处理; L-CK: 光照对照; L-HT: 光照, 5 mmol L-1 2-羟乙基肼处理。标以不同大写字母于处理间表示在0.01水平上差异显著。Fig. 2 Effects of light on PAO activity and mesocotyl elongation in maizeD-CK: dark contrast; D-HT: dark, 5 mmol L-1 2-hydroxyethylhydrazine; L-CK: light contrast; L-HT: light, 5 mmol L-1 2-hydroxyethylhydrazine. Bars superscripted by different letters are significantly different at the 0.01 probability level among treatments.
图3 光对玉米中胚轴H2O2含量和长度的影响D-CK: 黑暗对照; D-DT: 黑暗, 5 mmol L-1 N, N’ -二甲基硫脲处理; L-CK: 光照对照; L-DT: 光照, 5 mmol L-1 N, N’ -二甲基硫脲处理。标以不同大写字母于处理间表示在0.01水平上差异显著。Fig. 3 Effects of light on H2O2 content and mesocotyl elongation in maizeD-CK: dark contrast; D-DT: dark, 5 mmol L-1 N, N’ -Dimethylthiourea; L-CK: light contrast; L-DT: light, 5 mmol L-1 N, N’ -Dimethylthiourea.Bars superscripted by different letters are significantly different at the 0.01 probability level among treatments.
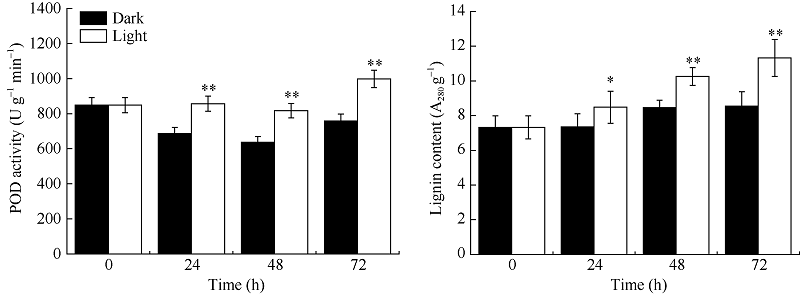
图5 光对玉米中胚轴POD活性和木质素含量的影响* * 表示在0.01水平上差异显著; * 表示在0.05水平上差异显著。Fig. 5 Effects of light on POD activity and lignin content in maize mesocotyl* * means significant difference at the 0.01 level; * means significant difference at the 0.05 level.
表2 中胚轴长度、PAO活性、H2O2含量、POD活性和木质素含量之间的相关系数 Table 2 Correlation coefficients among mesocotyl length, PAO activity, H2O2 content, POD activity, and lignin content
中胚轴长度 Mesocotyl length
PAO活性 PAO activity
H2O2含量 H2O2 content
POD活性 POD activity
木质素含量 Lignin content
中胚轴长度 Mesocotyl length
1
PAO活性 PAO activity
-0.884* *
1
H2O2含量 H2O2 content
-0.815* *
0.836* *
1
POD活性 POD activity
-0.597*
0.592*
0.581*
1
木质素含量 Lignin content
-0.752* *
0.744* *
0.938* *
0.600*
1
* * means significant correlation at the 0.01 level; * means significant correlation at the 0.05 level. * * 表示在0.01水平上显著相关; * 表示在0.05水平上显著相关。
表2 中胚轴长度、PAO活性、H2O2含量、POD活性和木质素含量之间的相关系数 Table 2 Correlation coefficients among mesocotyl length, PAO activity, H2O2 content, POD activity, and lignin content
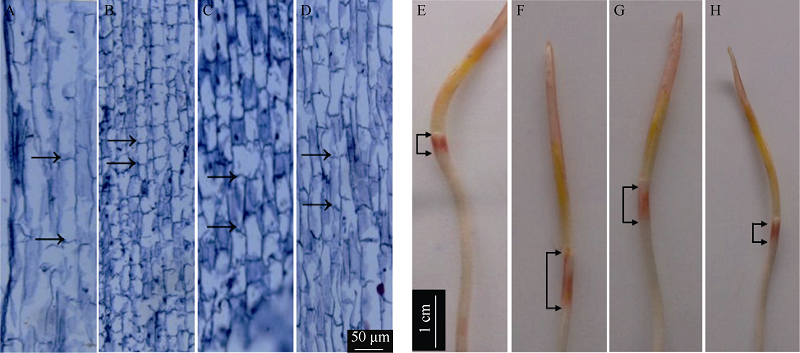
图6 玉米中胚轴表皮细胞伸长和H2O2组织化学染色A: D-CK处理的玉米中胚轴纵切面细胞长度; B: L-CK处理的玉米中胚轴纵切面细胞长度; C: L-HT处理的玉米中胚轴纵切面细胞长度; D: L-DT处理的玉米中胚轴纵切面细胞长度; E: D-CK处理的H2O2组织化学染色; F: L-CK处理的H2O2组织化学染色; G: L-HT处理的H2O2组织化学染色; H: L-DT处理的H2O2组织化学染色。Fig. 6 Microgram of epidermal cells of mesocotyl and histochemical H2O2 localization in maizeA: Cell length in longitudinal section of maize mesocotyl in treatment of D-CK; B: Cell length in longitudinal section of maize mesocotyl in treatment of L-CK; C: Cell length in longitudinal section of maize mesocotyl in treatment of L-HT; D: Cell length in longitudinal section of maize mesocotyl in treatment of L-DT; E: D-CK, histochemical H2O2 localization; F: Histochemical H2O2 localization in treatment of L-CK; G: Histochemical H2O2 localization in treatment of L-HT; H: Histochemical H2O2 localization in treatment of L-DT.
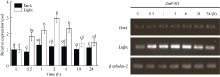
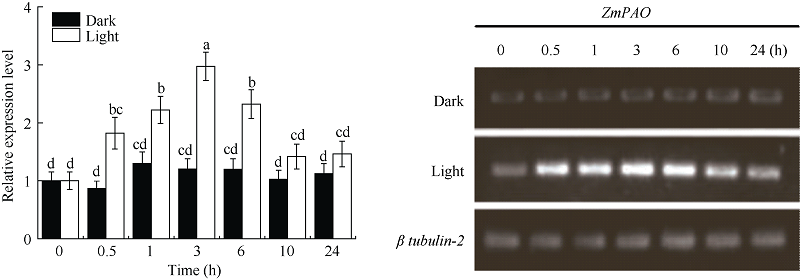
图7ZmPAO基因的表达模式分析标以不同大写字母于处理间表示在0.05水平上差异显著。Fig. 7 Expression pattern of ZmPAOBars superscripted by different letters are significantly different at the 0.05 probability level among treatments.
Dungan GH. Response of corn to extremely deep planting. Agron J, 1950, 42: 256-257[本文引用:1]
[2]
HoshikawaK. Underground organs of the seelings and the systematics of gramineae. Bot Gazette, 1969, 130: 192-203[本文引用:1]
[3]
张磊, 刘志增, 黄亚群, 陈景堂, 祝丽英. 46个玉米自交系耐深播特性分析. 河北农业大学学报, 2007, 30(3): 18-21ZhangL, Liu ZZ, Huang YQ, Chen JT, Zhu LY. Deep-planting tolerance characteristics analysis on 46 maize inbred lines. J Agric Univ Hebei, 2007, 30(3): 18-21 (in Chinese with English abstract)[本文引用:1]
[4]
赵光武, 马攀, 王建华, 王国英. 不同玉米自交系耐深播能力鉴定及对深播胁迫的生理响应. 玉米科学, 2009, 17(5): 9-13Zhao GW, MaP, Wang JH, Wang GY. Identification of deep-seeding tolerance in different maize inbred lines and their physiological response to deep-seeding condition. J Maize Sci, 2009, 17(5): 9-13 (in Chinese with English abstract)[本文引用:1]
[5]
杜金友, 张桂荣, 蔡爱军, 刘艳芳, 高桂花, 靳占忠. 玉米中胚轴长度与内源激素关系的研究. 玉米科学, 2008, 16(3): 70-73Du JY, Zhang GR, Cai AJ, Liu YF, Gao GH, Jin ZZ. Relationship between the mesocotyl elongation and hormones in maize (Zea mays L. ). J Maize Sci, 2008, 16(3): 70-73 (in Chinese with English abstract)[本文引用:2]
[6]
Stuart DA, Durnam DJ, Jones RL. Cell elongation and cell division in elongating lettuce hypocotyl sections. Planta, 1977, 135: 249-255[本文引用:1]
李莉, 马殿荣, 陈温福, 孙健, 梁茜. 杂草稻中胚轴伸长的细胞形态学观察. 沈阳农业大学学报, 2012, 43: 749-753LiL, Ma DR, Chen WF, SunJ, LiangQ. Observation of mesocotyl cell morphology of weed rice. J Shenyang Agric Univ, 2012, 43: 749-753 (in Chinese with English abstract)[本文引用:1]
[9]
周德宝. 植物激素与细胞骨架的排向. 植物生理学通讯, 2005, 41: 224-228Zhou DB. Plant hormones and layout of cytoskeleton. Plant Physiol Commun, 2005, 41: 224-228 (in Chinese with English abstract)[本文引用:1]
KarpinskaB, KarlssonM, SchinkelH, StrellerS, Suss KH, MelzerM, WingsleG. A novel superoxide dismutase with a high isoelectric point in higher plants expression, regulation, and protein localization. Plant Physiol, 2001, 126: 1668-1677[本文引用:1]
[12]
JiangM, ZhangJ. Involvement of plasma-membrane NADPH oxidase in abscisic acid- and water stress-induced antioxidant defense in leaves of maize seedlings. Planta, 2002, 215: 1022-1030[本文引用:2]
[13]
FrahryG, SchopferP. NADH-stimulated, cyanide-resistant superoxide production in maize coleoptiles analyzed with a tetrazolium-based assay. Planta, 2001, 212: 175-183[本文引用:2]
[14]
YodaH, YamaguchiY, SanoH. Induction of hypersensitive cell death by hydrogen peroxide produced through polyamine degradation in tobacco plants. Plant Physiol, 2003, 132: 1973-1981[本文引用:3]
[15]
SuzukiY, YanagisawaH. Purification and properties of maize polyamine oxidases: a flavprotein. Plant Cell Physiol, 1980, 21: 1085-1094[本文引用:1]
AngeliniR, FedericoR, BonfeuteP. Maize polyamine oxidase: antibody production and ultrastructural localization. Plant Physiol, 1995, 145: 686-692[本文引用:1]
[18]
TavladorakiP, Schininà ME, CecconiF. Maize polyamine oxidase: primary structure from protein and cDNA sequencing. FEBS Lett, 1998, 426: 62-66[本文引用:1]
[19]
BindaC, CodaA, AngeliniR, FedericoR, AscenziP, MatteviA. A 30 Å long U-shaped catalytic tunnel in the crystal structure of polyamine oxidase. Structure, 1999, 7: 265-276[本文引用:2]
[20]
CervelliM, TavladorakiP, Di AgostinoS, AngeliniR, FedericoR, MariottiniP. Isolation and characterization of three polyamine oxidase genes from Zea mays. Physiology, 2000, 38: 667-677[本文引用:1]
[21]
FedericoR, AngeliniR. Occurrence of diamine oxidase in the apoplast of pea epicotyls. Planta, 1986, 167: 300-302[本文引用:1]
[22]
LaurenziM, ReaG, FedericoR, TavladorakiP, AngeliniR. De-etiolation causes a phytochrome-mediated increase of polyamine oxidase expression in outer tissues of the maize mesocotyl: a role in the photomodulation of growth and cell wall differentiation. Planta, 1999, 208: 146-154[本文引用:2]
[23]
ConaA, CenciF, CervelliM, FedericoR, MariottiniP, MorenoS, AngeliniR. Polyamine oxidase, a hydrogen peroxide-producing enzyme, is up-regulated by light and down-regulated by auxin in the outer tissues of the maize mesocotyl. Plant Physiol, 2003, 131: 803-813[本文引用:3]
[24]
汪天, 郭世荣, 刘俊, 高洪波. 多胺氧化酶检测方法的改进及其在低氧水培黄瓜根系中的应用. 植物生理学通讯, 2004, 40: 358-360WangT, Guo SR, LiuJ, Gao HB. An improved method for measuring polyamine oxidase and its application to the study of cucumber root under hypoxic stress. Plant Physiol Commun, 2004, 40: 358-360 (in Chinese with English abstract)[本文引用:1]
[25]
BrennanT, FrenkelC. Involvement of hydrogen peroxide in the regulation of senescence in pear. Plant Physiol, 1977, 59: 411-416[本文引用:1]
[26]
Orozco-CardenasM, Ryan CA. Hydrogen peroxide is generated systemically in plant leaves by wounding and systemin via the octadecanoid. Proc Natl Acad Sci USA, 1999, 96: 6553-6557[本文引用:1]
SyrosT. Activity and isoforms of peroxidase, lignin and anatomy, during adventitious rooting cuttings of Ebenus cretica L. J Plant Physiol, 2004, 161: 69-77[本文引用:1]
[29]
TanakaK, NakamuraY, AsamiT, YoshidaS, MatsuoT, OkamotoS. Physiological roles of brassinosteroids in early growth of Arabidopsis: brassinosteroids have a synergistic relationship with gibberellin as well as auxin in light-grown hypocotyl elongation. J Plant Growth Regul, 2003, 22: 259-271[本文引用:1]
[30]
苏国兴, 刘友良. 高等植物体内的多胺分解代谢及其主要产物的生理作用. 植物学通报, 2005, 22: 408-418Su GX, Liu YL. Function of polyamine catabolism and its main catabolic products in higher plants. Chin Bull Bot, 2005, 22: 408-418 (in Chinese with English abstract)[本文引用:1]
[31]
KarpinskaB, KarlssonM, SchinkelH, StrellerS, Suss KH, MelzerM, WingsleG. A novel superoxide dismutase with a high isoelectric point in higher plants expression, regulation, and protein localization. Plant Physiol, 2001, 126: 1668-1677[本文引用:2]
[32]
王艺, 韦小丽. 不同光照对植物生长、生理生化和形态结构影响的研究进展. 山地农业生物学报, 2010, 29: 353-359WangY, Wei XL. Advance on the effects of different light environments on growth, physiological biochemistry and morphostructure of plant. J Mountain Agric & Biol, 2010, 29: 353-359 (in Chinese with English abstract)[本文引用:1]
[33]
杜成凤, 刘天学, 蒋寒涛, 李潮海. 弱光胁迫及光恢复对玉米幼苗活性氧代谢的影响. 核农学报, 2011, 25: 570-575Du CF, Liu TX, Jiang HT, Li CH. Effects of low light stress and light recovery on reactive oxygen metabolism of maize seedlings. J Nuclear Agric Sci, 2011, 25: 570-575 (in Chinese with English abstract)[本文引用:1]
HohlM, GreinerH, SchopferP. The cryptic-growth response of maize coleoptiles and its relationship to H2O2-dependent cell wall stiffening. Physiol Plant, 1995, 94: 491-498[本文引用:2]
[36]
Zhao GW, Wang JH. Effect of gibberellin and uniconazole on mesocotyl elongation of dark-grown maize under different seeding depths. Plant Prod, 2008, 11: 423-429[本文引用:1]
[37]
ReaG, LurenziM, RanquilliE, D’Ovidio R, Federico R, Angelini R. Developmentally and wound-regulated expression of the gene encoding a cell wall copper amine oxidase in chickpea seedlings. FEBS Lett, 1998, 437: 177-182[本文引用:1]
[38]
SchopferP. Hydrogen peroxide-mediated cell-wall stiffening in vitro in maize coleoptiles. Planta, 1996, 199: 43-49[本文引用:1]
[39]
SebelaM, RadovaA, AngeliniR, TavladorakiP, Frebort II, PecP. FAD-containing polyamine oxidases: a timely challenge for researchers in biochemistry and physiology of plants. Plant Sci, 2001, 160: 197-207[本文引用:1]
 , 李永生
, 李永生







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}