关键词:水稻; 黄绿叶; 基因定位; 遗传分析 Genetic Analysis and Gene Mapping of a Yellow-green Leaf Mutant in Rice CHU Zhi-Zhan1, GUO Hai-Bin2, LIU Xiao-Lin3, CHEN Yuan-Ling1, LIU Yao-Guang1,* 1 College of Life Sciences, South China Agricultural University / State Key Laboratory for Conservation and Utilization of Subtropical Agro-bioresources, Guangzhou 510642, China
2 Center of Experimental Teaching for Common Basic Course, South China Agricultural University, Guangzhou 510642, China
3 Yichun University, Yichun 336000, China
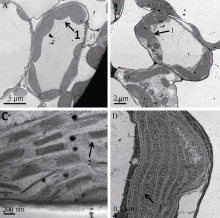
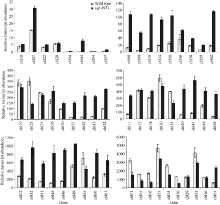
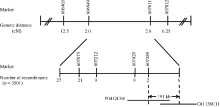
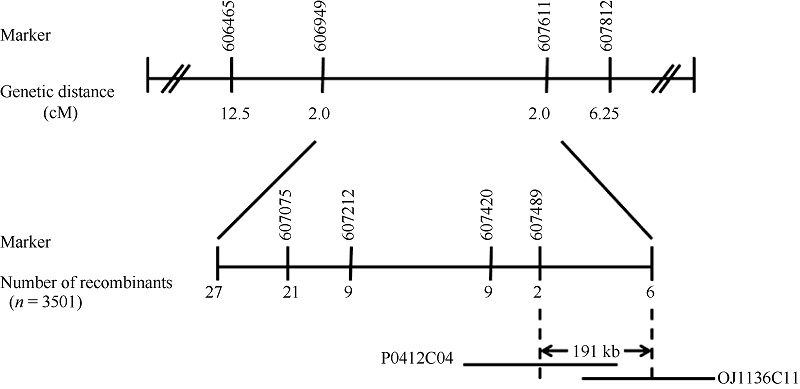
Fund:This study was supported by the Open Fund Project of State Key Laboratory for Conservation and Utilization of Subtropical Agro-bioresources (SKL-CUSAb-2013-04), the Science and Technology Program from Education Department of Jiangxi Province (GJJ14707), and the Natural Science Fund of Guangdong Province-the Launch Program for Doctor (2015A030310485) AbstractA yellow-green leaf rice mutant, temporarily named as ygl8951 (yellow-green leaf 8951), was identified from60Co γ- ray radiation mutation in japonica rice variety Nipponbare. The mutant showed albino phenotype from germination to 3-leaf-stage, then turned yellow-green phenotype till apoptosis. The contents of chlorophyll and carotenoid were obviously decreased in ygl8951 compared with the wild type. Electron microscope observation showed that no grana thylakoids but some stroma thylakoids-like structures were found in chloroplast of ygl8951 mutant. The expression levels of some genes involved in photosystem I and photosystem II were dramatically decreased, while the ribosomal and RNA polymerase genes in chloroplast were increased in ygl8951 mutant compared with the wild type. Mapping-based cloning was used to identify the ygl8951 locus using an F2 population from a crossing between the mutant and Huanghuazhan. The result showed that the mutated locus was located in a 191 kb region on chromosome 6, which was assumed to be a new gene controlling leaf color.
LeisterD. Chloroplast research in the genomic age. Trends Genet, 2003, 19: 47-56[本文引用:1]
[2]
董凤高, 朱旭东, 熊振民. 以淡绿叶为标记的籼型光-温敏核不育系M2S的选育. 中国水稻科学, 1995, 9: 65-70Dong FG, Zhu XD, Xiong ZM. Breeding of a photo-thermoperiod sensitive genic male sterile indica rice with a pale-green-leaf marker. Chin J Rice Sci, 1995, 9: 65-70 (in Chinese with English abstract)[本文引用:1]
[3]
Larkin RM, Alonso JM, Ecker JR, ChoryJ. GUN4, a regulator of chlorophyll synthesis and intracellular signaling. Science, 2003, 299: 902-906[本文引用:1]
[4]
Wang PY, Gao JX, Wan CM, Zhang FT, Xu ZJ, Huang XQ, Sun XQ, Deng XJ. Divinyl chlorophyll(ide) a can be converted to monovinyl chlorophyll(ide) a by a divinyl reductase in rice. Plant Physiol, 2010, 153: 994-1003[本文引用:1]
[5]
Wu ZM, ZhangX, HeB, Diao LP, Sheng SL, Wang JL, Guo XP, SuN, Wang LF, JiangL, Wang CM, Zhai HQ, Wan JM. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol, 2007, 145: 29-40[本文引用:1]
[6]
LeeS, Kim JH, Yoo ES, Lee CH, HirochikaH, AnG. Differential regulation of chlorophyll an oxygenase genes in rice. Plant Mol Biol, 2005, 57: 805-818[本文引用:1]
[7]
FangJ, ChaiC, QianQ, Li CL, Tang JY, SunL, Huang ZJ, Guo XL, Sun CH, LiuM, ZhangY, Lu QT, Wang YQ, Lu CM, HanB, ChenF, Cheng ZK, Chu CC. Mutations of genes in synthesis of the carotenoid precursors of ABA lead to preharvest sprouting and photo-oxidation in rice. Plant J, 2008, 54: 177-189[本文引用:1]
[8]
Gothand am KM, Kim ES, Chung YY. OsPPR1, a pentatricopeptide repeat protein of rice is essential for the chloroplast biogenesis. Plant Mol Biol, 2005, 58: 421-433[本文引用:1]
SugimotoH, KusumiK, TozawaY, YazakiJ, KishimotoN, KikochiS, IbaK. The virescent-2 mutation inhibits translation of plastid transcripts for the plastid genetic system at an early stage of chloroplast differentiation. Plant Cell Physiol, 2004, 45: 985-996[本文引用:1]
[11]
Zhao CF, Xu JM, ChenY, Mao CZ, Zhang SL, Bai YH, Jiang DA, WuP. Molecular cloning and characterization of OsCHR4, a rice chromatin-remodeling factor required for early chloroplast development in adaxial mesophyll. Planta, 2012, 236: 1165-1176[本文引用:1]
[12]
Jiang HW, Li ML, Liang NT, Yan HB, Wei YB, Xu XL, LiuJ, Xu JF, ChenF, Wu GJ. Molecular cloning and function analysis of the stay green gene in rice. Plant J, 2007, 52: 197-209[本文引用:1]
[13]
SakurabaY, ParkS, PaekN. The divergent roles of STAYGREEN (SGR) homologs in chlorophyll degradation. Mol Cells, 2015, 38: 390-395[本文引用:1]
[14]
ParkS, YuJ, ParkJ, LiJ, YooS, LeeN, JeongS. The senescence-induced stay-green protein regulates chlorophyll degradation. Plant Cell, 2007, 19: 1649-1664[本文引用:1]
[15]
KusabaM, ItoH, MoritaR, MoritoR, LidaS, SatoY, FujimotoM, KawasakiS, TanakaR, HirochikaH, NishimuraM, TanakaA. Rice NON-YELLOW COLORING1 is involved in light- harvesting complex II and grana degradation during leaf senescence. Plant Cell, 2007, 19: 1362-1375[本文引用:1]
[16]
Wellburn AR. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. Plant Physiol, 1994, 144: 307-313[本文引用:1]
[17]
GuillemautP, Mardchal-DrouardL. Isolation of plant DNA: a fast, inexpensive, and reliable method. Plant Mol Biol Rep, 1992, 10: 60-65[本文引用:1]
[18]
王慧娜, 初志战, 马兴亮, 李日清, 刘耀光. 高通量PCR模板植物基因组DNA制备方法. 作物学报, 2013, 39: 1200-1205Wang HN, Chu ZZ, Ma XL, Li RQ, Liu YG. A high through- put protocol of plant genomic DNA preparation for PCR. Act Agron Sin, 2013, 39: 1200-1205 (in Chinese with English abstract)[本文引用:1]
[19]
刘朝辉, 李小艳, 张建辉, 林冬枝, 董彦君. 一个新的水稻叶绿素缺失黄叶突变体的特征及基因分子定位. 遗传, 2012, 34: 223-229Liu CH, Li XY, Zhang JH, Lin DZ, Dong YJ. Characteristics and molecular mapping of a novel chlorophyll-deficient yellow-leaf mutant in rice. Hereditas (Beijing), 2012, 34: 223-229 (in Chinese with English abstract)[本文引用:1]
[20]
孙小秋, 王兵, 肖云华, 万春美, 邓晓建, 王平荣. 水稻ygl98黄绿叶突变基因的精细定位与遗传分析. 作物学报, 2011, 37: 991-997Sun XQ, WangB, Xiao YH, Wan CM, Deng XJ, Wang PY. Genetic analysis and fine-mapping of ygl98 yellow-green leaf gene in rice. Acta Agron Sin, 2011, 37: 991-997 (in Chinese with English abstract)[本文引用:1]
[21]
孔萌萌, 余庆波, 张慧绮, 盛春, 周根余, 杨仲南. 控制水稻叶绿体发育基因OsALB23的定位. 植物生理与分子生物学学报, 2006, 32: 433-437Kong MM, Yu QB, Zhang HQ, ShengC, Zhou GY, Yang ZN. Genetic mapping of rice gene OsALB23 regulating chloroplast development. J Plant Physiol Mol Biol, 2006, 32: 433-437 (in Chinese with English abstract)[本文引用:1]
[22]
HiratsukaJ, ShimadaH, WhittierR, IshibashiT, SakamotoM, MoriM, KondoC, HonjiY, Sun CR, Meng BY, Li YQ, KannoA, NishizawaY, HiraiA, ShinozakiK, SugiuraM. The complete sequence of the rice (Oryza sativa) chloroplast genome: Intermolecular recombination between distinct tRNA genes accounts for a major plastid DNA inversion during the evolution of the cereals. Mol Gen Genet, 1989, 217: 185-194[本文引用:1]
AlbertssonP. A quantitative model of the domain structure of the photosynthetic membrane. Trends Plant Sci, 2001, 6: 349-354[本文引用:1]
[25]
Hajdukiewicz PT, Allison LA, MaligaP. The two RNA polymerases encoded by the nuclear and the plastid compartments transcribe distinct groups of genes in tobacco plastids. EMBO J, 1997, 16: 4041-4048[本文引用:1]
[26]
周华, 潘佑找, 刘秀艳, 马晓静, 陈素丽, 林冬枝, 王俊敏, 董彦君, 滕胜. 一个新的水稻叶绿素缺失黄叶突变体遗传分析及其基因定位. 分子植物育种, 2013, 11: 145-151ZhouH, Pan YZ, Liu XY, Ma XJ, Chen SL, Lin DZ, Wang JM, Dong YJ, TengS. Genetic analysis and molecular mapping of a novel yellow leaf mutant in rice. Mol Plant Breed, 2013, 11: 145-151 (in Chinese with English abstract)[本文引用:1]
[27]
王军, 王宝和, 周丽慧, 徐洁芬, 顾铭洪, 梁国华. 一个水稻新黄绿叶突变体基因的分子定位. 中国水稻科学, 2006, 20: 455-459WangJ, Wang BH, Zhou LH, Xu JF, Gu MH, Liang GH. Genetic analysis and molecular mapping of a new yellow- green leaf gene ygl-2 in rice. Chin J Rice Sci, 2006, 20: 455-459 (in Chinese with English abstract)[本文引用:1]






{kind=link}
{kind=link}
{kind=link}
{kind=link}